|
А.П.Расницын. Теоретические основы эволюционной биологии // В.В.Жерихин, А.Г.Пономаренко, А.П.Расницын. Введение в палеоэнтомологию. М.: КМК. 2008. 371 с
1.2.2.3. Филетика (продолжение)
1.2.3. ТАКСОНОМИЧЕСКАЯ НОМЕНКЛАТУРА
1.2.3.1. Таксономически независимые принципы
1.2.3.2. Таксономически зависимые принципы
|
1.2.3. ТАКСОНОМИЧЕСКАЯ НОМЕНКЛАТУРА
Номенклатура — это совокупность договоренностей о том, как создавать и использовать названия таксонов. Обычно она воспринимается как область усложненных деталей и простых до очевидности базовых принципов. Это неверно, во-первых, поскольку еще далеко не все действующие принципы биологической номенклатуры осознаны и сформулированы в явном виде. Некоторые из них упомянуты в кодексах (см. МКЗН, 1988, 2000; МКБН, 2000), но не в качестве базовых принципов, а многих принципов там просто нет. Во-вторых, некоторые из базовых принципов не являются чистыми соглашениями, а отражают определенные таксономические концепции или представляют следствия из определенных эволюционных гипотез. Линнеев принцип биноминальной номенклатуры вполне конвенционален и потому легко сочетается с любой таксономической концепцией, признающей иерархию родов и видов. Иное дело принцип типа, как это подробно обсуждается далее. Различение этих двух типов принципов представляется полезным, и в дальнейшем они будут именоваться таксономически независимыми и таксономически зависимыми принципами соответственно (табл. 4).
|
Таксономически независимые |
|
|
1 |
невмешательства в принятие таксономических решений за пределами ограничений, налагаемых таксономически зависимыми принципами |
|
2 |
ограниченной сферы приложения |
|
3 |
гомонимии |
|
4 |
приоритета |
|
5 |
стандартизированных, ранг-специфичных названий |
|
6 |
верховного приоритета Международной Комиссии по номенклатуре |
|
Таксономически зависимые |
|
|
7 |
типа (нет разумных причин отвергать) |
|
8 |
иерархии (нет разумных причин отвергать) |
|
9 |
синонимии (применение к паратаксонам ограничено рамками соответствующей частной системы) |
1.2.3.1. Таксономически независимые принципы
Таксономически независимые принципы обычно просты, не требуют больших комментариев и здесь рассматриваются первыми.
1. В действующем Кодексе зоологической номенклатуры сказано, что он «воздерживается от вмешательства в таксономические суждения, которые не могут подвергаться регулированию» (МКЗН, 2000: 24). Это утверждение не вполне справедливо из-за существования таксономически зависимых принципов, которые не позволяют принимать некоторые таксономические решения. Поэтому формулировка требует дополнения словами «за пределами ограничений, налагаемых Кодексом».
2. В зоологии область применения принципов номенклатуры ограничена надпопуляционными группами от подвида до надсемейства (подробнее см. МКЗН, 2000, ст. 1.2). Исключение старших таксонов из сферы действия кодекса представляется ошибочным (Rasnitsyn, 1982, 1989, 1991, 2002b). Вкратце мои соображения можно изложить следующим образом.
Обеспечиваемая кодексами стандартизация образования и использования названий доказала долгой практикой свое благотворное влияние на всю биологию. Отказ от ее распространения на старшие таксоны вряд ли связан с их глубинной спецификой. Серьезных указаний на такую специфику не известно, а многочисленные попытки стандартизировать не типифицированные названия старших таксонов (концовки -ptera у названий отрядов насекомых, -formes у отрядов птиц, -ida у многих названий вымерших отрядов разных групп беспозвоночных) снимают все сомнения в необходимости такой унификации. Дело, конечно, не в унификации, а в типификации названий старших таксонов. Под типификацией (более точно — сквозной типификацией) понимается процедура, обеспечивающая жесткую связь каждого названия каждого таксона с определенным подчиненным таксоном в его составе, сверху вниз по всей иерархии вплоть до типового экземпляра вида (или подвида). Типифицируются именно названия, а не сами таксоны: когда говорят о голотипе вида или типовом виде рода, имеют в виду тип названия (имени).
Принцип типа, соответствующий самой природе таксона (см. ниже) является главным достижением биологической номенклатуры: он обеспечивает устойчивость употребления и упорядоченность изменения названия таксона при самых различных изменениях его состава (границ) и интерпретации (изменение ранга, диагностических признаков и т.п.). Первые попытки типификации старших таксонов насчитывают уже более двух веков (Laicharting, 1781), и понимание ее необходимости также растет. Такая точка зрения победила в ботанике (МКБН, 1985) и находит некоторую поддержку в зоологии (Родендорф, 1977; Starobogatov, 1991; Клюге, 2000). Дело в том, что не типифицированное название отнесено к таксону как к целому и практично только до тех пор, пока интерпретация самого таксона остается устойчивой. В противном случае возникают проблемы с применением названия, что выливается в разнобой и переименования. Однако введение типифицированных названий старших таксонов и, соответственно, их переименование затрагивает интересы многочисленных пользователей, которых мало интересуют проблемы систематики высоких уровней, но пугает перспектива заучивать новые слова. Для тех же, кто работает со старшими таксонами, ситуация оказывается тяжелой: таксономические и филогенетические ошибки и неточности консервируются не типифицированными названиями и накапливаются. Чем дольше это продолжается, тем труднее будет переходить к типификации старших таксонов. Но переходить все равно придется, и потомки не скажут нам спасибо за наш эгоистичный консерватизм.
3. Идентичные названия разных таксонов не должны использоваться в качестве действительных названий (МКЗН, 2000, ст. 52). Область применения принципа гомонимии произвольно ограничена таким образом, чтобы не затрагивать случаев гомонимии, во-первых, между названиями видов в разных родах, во-вторых, между таксонами растений и животных, в-третьих, с таксонами рангом выше надсемейства и между такими таксонами.
4. Выбор между конкурирующими названиями (синонимами или гомонимами) должен осуществляться в пользу того названия, которое было предложено раньше (принцип приоритета; МКЗН, 2000, ст. 23).
5. Принцип стандартизированной и ранг-специфичной формы названий таксонов. Этот принцип представляет основу для многочисленных правил для языка, грамматики и синтаксиса названий, и включает принцип биноминальной номенклатуры (МКЗН, 2000, ст. 4-6, 11, 25-34).
6. Последний среди таксономически независимых принципов — принцип верховенства Международной Комиссии по номенклатуре, которая может принимать решения в нарушение любых положений Кодекса, кроме тех, которые касаются ее самой (МКЗН, 2000, ст. 78).
1.2.3.2. Таксономически зависимые принципы
Таксономически зависимых принципов три:
7. Принцип типа. Это самый важный из таксономически зависимых принципов, и на его примере хорошо видны те ограничения, которые таксономическая концепция может накладывать на номенклатуру. В Кодексе 1988 г. этот принцип сформулирован следующим образом: «Номенклатурный тип служит объективным эталоном, с помощью которого приложение названия определяется независимо от возможных изменений границ таксона» (МКЗН, 1988, ст. 61а; формулировка MКЗН 2000, ст. 61.1, кажется менее удачной: «Фиксация номенклатурного типа номинального таксона обеспечивает объективный эталон для приложения названия этого таксона»). Другими словами, таксон не может быть введен в систему иначе как, в конечном счете, ссылкой на тип, несущий его название. Чтобы оценить смысл этого утверждения, рассмотрим альтернативные возможности.
Первое, что приходит в голову, это возможность введения таксона в систему ссылкой на его признаки. Это означает, что признаки таксона выполняют определяющую, а не просто диагностирующую функцию, а сам таксон соответствует понятию «класс» (Ghiselin, 1974, 1987). Существуют разные возможности такого введения таксона в систему, простейший из которых — комбинаторная система, т.е. многомерная матрица, каждая ячейка которой соответствует определенной комбинации признаков и является местом таксона, обладающего такой комбинацией. Версию такой матрицы представляет политомическая таблица, иногда используемая систематиками, т.е. прямоугольная матрица с рядами, представляющими таксоны, и колонками, соответствующими признакам. В результате каждая ячейка отражает состояние одного конкретного признака в определенном таксоне. Эта форма системы действительно проста и иногда очень полезна, но не соответствует задачам общей системы организмов. Она не обладает гибкостью: мы не можем исправить ее локально, чтобы, например, лучше согласовать ее с признаками конкретного таксона, поскольку введение нового, переопределение или удаление старого признака заставляет пересматривать характеристики всех таксонов. Поэтому даже в качестве средства идентификации комбинативная система используется нечасто.
Более гибка система, в которой таксоны определяются ранжированными признаками. В обычной определительной таблице первый используемый признак имеет самый высокий ранг, поскольку делит определяемую совокупность на самые большие группы. Вводимые далее признаки обеспечивают дальнейшее ее дробление и имеют более низкий ранг. Такая система действительно удобна, хотя и не столь хороша, как общая система организмов, из-за очевидно произвольного ранжирования признаков. Тем не менее, в несколько модифицированной форме (с выделением небольшого числа высокоранговых признаков, достаточного для формирования системы) этот подход был весьма популярен у систематиков. У К. Линнея (Linnaeus, 1751), например, близкую роль играли признаки фруктификаций растений. В наиболее четкой форме этот подход был сформулирован A.A. Любищевым (1923, 1966). Любищев призывал к поиску немногих наиболее высокоранговых признаков (параметров), детерминирующих распределение всех других признаков, — так же, как ядерный заряд определяет свойства атомов и, соответственно, положение элементов в таблице Менделеева. В результате ожидалось построение параметрической системы организмов, которая позволит прогнозировать все важные свойства соответствующих таксонов по этим ключевым признакам (параметрам). Поставленная Любищевым задача казалась безнадежной, и ему самому не удалось ее разрешить. Тем не менее, решение в каком-то смысле было найдено, но совсем не там, где он его искал. По иронии судьбы это произошло в области, которую сам Любищев как убежденный антиселекционист считал ложной.
Решением оказалась кладистическая система с ее центральным утверждением, что признаки организма и, соответственно, его положение в системе наилучшим образом определяются единственным признаком — родством. Родство оказалось любищевским параметром по определению. Смысл и возможности кладистической системы обсуждены выше, здесь я хочу только привлечь внимание к тому факту, что эта система не нуждается в типификации ее таксонов, поскольку ссылка на значение параметра (характер родственных отношений) достаточна для введения таксона в систему.
Действительно, «названия синонимичны, если они указывают на клады [ветви], отходящие от одного и того же предка» (de Queiroz, Gauthier, 1990: 307). Логический вывод из этого утверждения, именно отказ от принципа типа (не говоря уже о Линнеевой системе стандартных таксономических рангов и прочих мелочах) был сделан Ф. Плейелом с соавторами (Sundberg, Pleijel, 1994; Plejel, Rouse, 2002), и соответствующий кодекс филогенетической номенклатуры (Филокод) находится в процессе разработки. Предполагается, что согласно этому кодексу роль типа, фиксирующего применение названия таксона, будет исполнять либо узел кладограммы, либо ее междоузлие, либо апоморфия. Авторы заявляют о больших преимуществах филогенетической номенклатуры в отношении как простоты применения, так и ее стабильности (Plejel, Rouse, 2002). Мне трудно в это поверить из-за высокой степени гипотетичности кладограммы, которая представляет собой систему гипотез о ненаблюдаемой истории наблюдаемых признаков (см. выше). Впрочем, на этом месте разумнее остановиться и дать поработать времени. Оно покажет, насколько жизнеспособна таксономическая номенклатура, построенная на кладистических принципах.
Логической альтернативой класса как группы, определяемой ее признаками, является индивидуум, и предложение считать таксон индивидуумом (Ghiselin, 1974, 1987 и приведенная там библиография) вполне естественно. Обладая свойствами индивидуума, таксон может вводится в систему остенсивно, т.е. прямым указанием (в том числе и ссылкой на его название). Для этого его целостность в пространстве и во времени должна быть достаточно высокой, чтобы позволять таксону рождаться и умирать, но не распадаться на части, каждая из которых могла бы претендовать на наследование имени таксона. Пусть человек на пути от младенца к старику не сохранил ни одного общего атома вещества и ни одного общего признака, но при всех его метаморфозах всегда можно проследить, что он является одним и тем же лицом.
Индивидуум настолько целостен, что использование его имени не вызывает проблем. Mожно просто указать пальцем — вот оно, и не детализировать, к чему именно относится имя — к голове, сердцу, уху или чему-то еще. Ко всему сразу и к каждой детали в отдельности. Именно здесь проходит граница между остенсивным методом и принципом типа: если любая часть индивидуума равно является носителем его названия, специальные правила выбора и использования номенклатурного типа, служащего для введения таксона в систему, оказываются излишними.
Если метод типа не приложим к индивидууму, то таксон не является индивидуумом. По крайней мере он не типичный индивидуум (см., например, комментарии к статье Ghiselin, 1981), так что проблема требует дальнейшего обсуждения. Парадигмальный пример индивидуума, организм, целостен из-за непрерывного взаимодействия его частей. То же самое утверждалось в отношении вида, целостность которого обеспечена обменом генами (Майрова концепция биологического вида). Однако эта модель имеет ограниченное приложение, хотя бы потому, что «Концепция биологического вида применима лишь к тому, что я назвал 'непротяженной ситуацией', когда популяции вида находятся в реальном [репродуктивном] контакте» (Mayr, 1988: 301-302). На самом деле проблем здесь, как мы уже видели, еще больше (см. раздел об онтологии эволюции).
Однако дихотомия класс — индивидуум не исчерпывает всех возможностей. Таксон обладает чертами обеих этих крайностей. В качестве класса он обладает признаками, о чем свидетельствует наличие диагноза, и членами (вопреки Гислину, я являюсь не только частью вида Homo sapiens, но и его экземпляром, т.е. примером, представителем). В качестве индивидуума таксон обладает частями (популяции по отношению к виду) и, что важнее, способностью развиваться (эволюционировать), не теряя своей индивидуальности. Развиваясь, таксон сохраняет свою целостность во времени, в многомерном пространстве признаков и, в той или иной степени, в географическом пространстве. Такая целостность позволяет идентифицировать таксон как индивидуум, т.е. по его имени. В результате таксон, обладая признаками и класса, и индивидуума, заполняет логическое пространство между этими понятиями и превращает класс и индивидуум в два полюса единого спектра.
Таким образом, таксон есть нечто промежуточное между классом и индивидуумом. Он достаточно целостен, чтобы, меняясь, сохранять имя как средство опознания, но недостаточно целостен, чтобы простого «вот оно» было достаточно для его опознания. Недостаточно целостен, потому что таксон — это континуум, сгущение точек, облако в многомерном пространстве признаков. Облако, которое может эволюционировать, т.е. менять свои размеры и положение в пространстве признаков, может исчезнуть (вымереть) или, наоборот, разделится пополам либо отпочковать новое облако, и т.д. Континуум характеризуется как целостностью, выявляемой по наличию гиатуса (нарушения непрерывности) между ним и другими облаками-континуумами, так и признаками его диагноза. Однако оба эти критерия не обнаруживают жесткости и легко изменяются как в ходе эволюции, так и по мере накопления наших знаний о таксоне. В любой момент может обнаружиться их неспособность четко идентифицировать и разграничить таксоны.
Возможность идентификации континуума, в отличие от индивидуума, определяется сходством, но в отличие от класса речь идет об отношении (общем сходстве), а не о сходстве по конкретным признакам. Поэтому указание пальцем не удается адресовать ни облаку в целом, ни его конкретным признакам. Оно должно быть адресовано определенной точке этого облака — его номенклатурному типу. Что бы ни произошло с таксоном-облаком — в ходе ли его эволюции, или при изменении наших представлениях о нем, но имя, закрепленное за номенклатурным типом, как этикетка за музейным экземпляром, всегда покажет нам, о чем идет речь. Если, конечно, тип в сохранности — или хотя бы описан с приемлемой полнотой.
Сказанное делает понятным, что номенклатурный тип есть естественный атрибут традиционной, т.е. филетической, а также фенетической системы, и, соответственно, выводится из эпигенетической теории эволюции. Что же касается кладизма, то кладистический таксон определяется, как мы видели, ссылкой на событие дивергенции или на синапоморфию, приобретенную этим таксоном (его первым членом) в момент возникновения. Именно дивергенция или/и приобретение синапоморфии мыслятся как событие, создающее таксон. Это событие есть признак, единолично и однозначно определяющий кладистический таксон. Таким образом, кладистический таксон есть класс по определению, и для его идентификации не нужен никакой номенклатурный тип: все, что необходимо и достаточно, это, как уже отмечалось, акт дивергенции и/или синапоморфия.
К тому же результату можно прийти, используя иной и более короткий путь рассуждений. Цель любой классификации состоит в том, чтобы покрыть таксонами все подлежащее классифицированию разнообразие без остатка и, по возможности, без перекрывания границ таксонов. Если мы принимаем, что и сами таксоны как естественные группировки, и образуемая ими система существуют в природе, а не придуманы систематиками (есть и такая точка зрения, см., например, С. Расницын, 2000), то главная цель классификации двуедина. Во-первых, прослеживание естественных перерывов непрерывности в классифицируемом пространстве (пользуясь платоновой метафорой, цитированной Hull, 1983: 186, мы расчленяем природу по ее сочленениям). Во-вторых, мы фиксируем и ранжируем выявленные гиатусы в качестве таксономических границ (при искусственной классификации мы налагаем эти границы вместо того, чтобы прослеживать их).
Классы не поддаются описанной процедуре, так как они определяются признаками, для которых естественно перекрываться с другими признаками (если только не определять класс так изощренно, как это я сделал выше, анализируя природу кладистического таксона). Что же касается индивидуумов, то нам не известно природных сил и закономерностей, которые могли бы упаковать индивидуумы плотно, не оставляя свободных промежутков (опять-таки, если не видеть в кладистическом таксоне индивидуум на основании того, что он рождается с дивергенцией своего предка, чтобы затем исчезнуть в результате вымирания или дивергенции). Таксон-континуум, напротив, идеально соответствует указанной процедуре классификации, поскольку единственный законный способ определения континуума — прослеживание и ранжирование его границ.
Не нужно думать, что это уникальное свойство общей системы организмов. Из рассуждений С.В. Мейена (1989) с очевидностью следует, что таковы же, например, мерон, т.е. таксон мерономии (морфологической системы частей организма), и стратон — таксон стратиграфической классификации.
Помимо принципа типа, существуют еще два таксономически зависимых принципа номенклатуры, которые, как и принцип типа, следуют из континуальной природы таксона.
8. Принцип иерархии. Будучи континуумами, таксоны не могут быть объединены иначе как в следующие более широкие континуумы (старшие таксоны), которые должны покрывать включаемые континуумы целиком. Равным образом таксон-континуум не может одновременно входить в два и более старших таксона ни целиком, ни частями, так как иначе эти старшие таксоны будут перекрываться, нарушая тем самым определение континуума. Все это означает, что система имеет строго иерархическую форму.
9. Принцип синонимии. Если таксон может быть членом единственного старшего таксона, он имеет единственное место в системе и, следовательно, ему достаточно одного имени. Все другие названия должны быть признаны недействительными, т.е. быть объявлены синонимами.
1.2.3.3. Таксоны, нарушающие принципы номенклатуры
Жизнь сложна, и наука тоже. Иногда нам приходится нарушать принципы номенклатуры — не по невежеству или разгильдяйству, а по необходимости, когда для этого возникают серьезные причины. Рассмотрим эти возможные причины.
Таксономически независимые принципы представляют собой соглашения между систематиками и в этом смысле субъективны. Причины отказа следовать им тоже всегда субъективны в том смысле, что они не оправданы какими-то таксономическими соображениями. Поэтому они здесь не рассматриваются. Проблема таксономически зависимых принципов более важна и заслуживает анализа. Более детально такой анализ проведен ранее (Расницын, 1986а) и здесь дается в сокращенном виде.
Нарушать таксономически зависимые принципы номенклатуры нам приходится, когда недостаток информации, необходимой для классификации некоего таксономического материала, вступает в конфликт с необходимостью его классифицировать — из-за научной или прикладной его значимости. Неполнота информации в этих случаях обычно связана со специфической дефектностью материала, как это хорошо известно в палеонтологии и в систематике групп с усложненным онтогенезом (например, у паразитических червей и грибов).
Международные кодексы номенклатуры признают некоторые формы таксонов особого статуса, но не решают проблему в целом, а содержащиеся в кодексах регламентации во многом противоречивы. Неудовлетворительность ситуации с таксонами особого статуса трудно считать неожиданной, по крайней мере в зоологии, где такие таксоны традиционно считаются чем-то редким и крайне нежелательным. В целом (как в зоологии, так и в ботанике) они действительно не слишком многочисленны, но в некоторых специфических областях (палеонтология, паразитология, микология) отказ от их использования грозит большой потерей информации об объектах, имеющих важное стратиграфическое, медицинское, агро- или лесохозяйственное значение. Использование «неполноценных» таксонов неизбежно, но при их неполной регламентации кодексами это ведет к номенклатурному разнобою и самодеятельности. В палеонтологии, например, в принципе однотипные группировки в одних случаях обозначают как специальные таксономические категории (ихнотаксоны в ихнологии, в частности «индузироды» и «индузиподроды» в систематике ископаемых домиков, построенных личинками насекомых — ручейников (Вялов, Сукачева, 1976), турмы, антетурмы и т.п. в палеопалинологии, формальные роды и недавно отмененные орган-роды в систематике макроостатков растений, и т.д.). В других случаях берут в кавычки название таксона (Белокрыс, 1984) либо таксономической категории (Пономаренко, 1985), в третьих — просто оговаривают статус таксона в описании или преамбуле. Порой дело доходит до разработки специальных кодексов номенклатуры (Sarjeant, Kennedy, 1973).
Этот разнобой неудобен, хотя и не опасен. Более того, он даже полезен как способ поиска оптимальных форм работы с «неполноценными» таксонами. Опасно порожденное негативным отношением к таким таксонам желание представить их нормальными, полноценными. Удивительно, например, стремление многих специалистов по конодонтам (изолированным микроскопическим элементам скелета древних животных), показать полноценными их таксоны (Melville, 1981a,b). Еще более демонстративна ситуация с современным родом Laberius Kieffer (перепончатокрылые насекомые семейства Dryinidae). Этот род был описан (под названием Labeo Hal., nom. praeocc.) как обычный таксон, но затем был использован как сборная группа для самцов триб Dryinini и Gonatopodini (Kiefer, 1914). Эти трибы характеризуются сильным половым диморфизмом, причем их система построена по признакам самок, а самцов, если они не ассоциированы с самками экспериментально, до последнего времени не удавалось определить даже до трибы. Однако в ревизии семейства (Olmi, 1984) ее автор отказался от такой интерпретации рода Laberius и свел его в синонимы рода Dicondylus Haliday (не обсуждая при этом признаки типового вида Laberius, т.е. не обосновывая своего решения). При этом вымерший вид, описанный в составе рода Laberius, понимаемого как сборная группа (Н. Пономаренко, 1981), был переведен в род Dryinus Latreille с указанием, что единственный экземпляр этого вида сохранился «настолько плохо, что это не позволяет отнести его к какому-либо роду семейства Dryinidae» (Olmi, 1984). Тем самым Dryinus (типовой род семейства) был превращен фактически в сборную группу, т.е. самую неполноценную форму таксона (см. ниже), хотя цель автора была явно обратной — избавиться от неполноценного таксона.
Таким образом, проблема таксонов, не удовлетворяющих тем или иным правилам номенклатуры, остается актуальной. Проанализируем более детально природу и формы таких таксонов (табл. 5).
Таблица 5. Формы таксонов
|
Таксон |
Принадлежит системе |
Тип и диагноз |
При синонимии с ортотаксоном |
|
ортотаксон |
общей |
собственный |
подлежит синонимизации |
|
таксон неясного положения |
общей |
собственный |
подлежит синонимизации |
|
морфотаксон (паратаксон) |
специальной |
собственный |
не подлежит синонимизации |
|
сборный таксон (паратаксон) |
специальной |
старшего таксона |
не подлежит синонимизации |
Самый легкий случай — это тaкcoн нeяcнoгo noлoжeнuя (taxon incertae sedis). Он отличается от нормального таксона (opтoтaкcoнa) тем, что неполнота его характеристики не позволяет уточнить его положение только на определенном уровне иерархии. Например, род неясного положения (genus incertae sedis) на данный момент может быть с достаточной уверенностью отнесен к определенному отряду, но не семейству. Вымерший род Baissobius Rasn., описанный среди перепончатокрылых насекомых подотряда Apocrita, «обладает своеобразным типом редуцированного жилкования, не встречающимся, насколько известно, у других Hymenoptera» (Расницын, 1975, с. 128). При этом его жилкование было достаточно своеобразно, чтобы затруднить определение систематического положения насекомого (другие важные признаки на ископаемом не видны), но недостаточно своеобразно, чтобы оправдать выделение особого семейства. И действительно, новые находки позволили расстаться с этим таксоном неясного положения, превратив его в нормальный род, принадлежащий семейству Embolemidae (Rasnitsyn, 1996b).
Использование таксонов неясного положения можно понять как локальный отказ от принципа синонимии. Род, подчиненный отряду, минуя семейство, фактически выступает в качестве самостоятельного, но не названного семейства, скорее всего синонимичного какому-то из существующих семейств, только пока неизвестно, какому именно. Как видно на примере Baissobius, со временем эта синонимия действительно может стать известной.
Более сложны случаи, объединенные понятием паратаксономии. Определение, трактующее паратаксон как таксон в частных системах для изолированных органов и частей тела животных (Melville, 1979), уже практического применения этого понятия. Действительно, специфика и проблемы, связанные с введением и использованием таксономического названия, мало отличаются в частных системах, созданных для изолированных органов и частей тела организмов, для определенного пола или стадии жизненного цикла, или для следов жизнедеятельности. Поэтому здесь принято другое определение: паратаксон в отличие от ортотаксона и таксона неясного положения принадлежит самостоятельной системе, связанной с основной системой принципом гомонимии, но не принципом синонимии, и потому не подлежит синонимизации даже в тех случаях, когда показана его синонимия с тем или иным ортотаксоном. Таким образом, паратаксон — это таксон, на который не распространяется требование уникальности таксономического положения, т.е. требование вхождения в единую для всех организмов иерархию. Именно к этому, в частности, сводятся все пять признаков своеобразия ископаемого материала, сформулированные С.В. Мейеном (Meyen, Traverse, 1979; Мейен, 1983). Важные соображения о природе и функциях паратаксона приведены в другой работе (Мейен, 1990).
Различают несколько видов паратаксонов. Термин формальный таксон был предложен (Расницын, 1986а) как более широкая интерпретация ботанического понятия форм-род. Недавно (МКБН, 2000) ботаники заменили этот термин более удачным понятием морфотаксон, которым мы и воспользуемся. Морфотаксон — это эквивалент нормального таксона (ортотаксона), но используемый только в рамках специальной системы. Эта система параллельна как основной системе, так и другим таким же частным системам, но независима от них в отношении принципа синонимии. Таковы системы изолированных ископаемых листьев, семян или отпечатков надкрыльев жуков, ископаемых следов передвижения организмов и других форм их жизнедеятельности (например, домиков личинок ручейников), система личиночных стадий современных паразитических червей или не ассоциированных с самками самцов в некоторых группах насекомых, где система ориентирована на признаки самок.
Морфотаксон отличается от ортотаксона только тем, что может быть, а порой точно является синонимом какого-то ортотаксона, и тем не менее он не подлежит синонимизации. Хорошо известно, например, что в ряде случаев для разрозненных частей древних растений (листьев, стволов, корней, мужских и женских фруктификаций, семян и пыльцы), описанных каждая в своей специальной системе, известны также находки в прижизненной связи, позволяющие «собрать» целое растение. Тем не менее каждая из таких частей сохраняет свое название, полученное в качестве морфотаксона. Так, хорошо сохранившиеся карбоновые стволы Lepidodendron, стволы Aspidiaria, Bergeria, Aspidiopsis, Knorria с корой, опавшей на разную глубину, корни Stigmaria, женские стробилы Lepidocarpon, мужские стробилы Lepidostrobus и микроспоры Lycospora (рис. 7) могли принадлежать одному и тому же растению, поскольку прижизненное сочетание разных пар из этого списка более или менее надежно установлено (Мейен, 1987б). И никто при этом не покушается на валидность всех этих родовых названий!
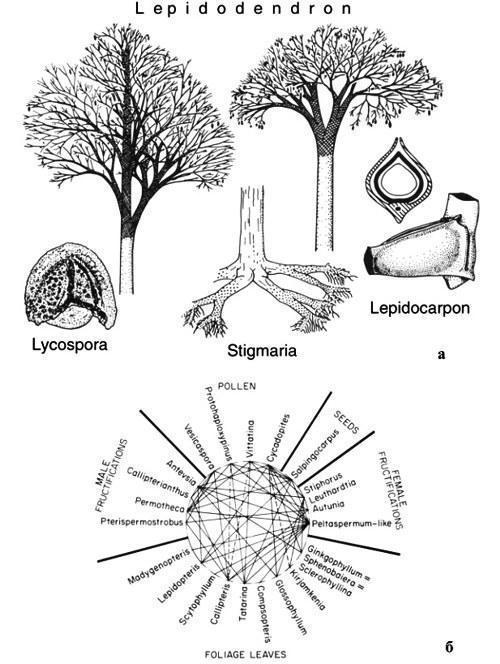
Рис. 7. Формальные таксоны, предложенные для классификации изолированных частей лепидофитов (по Мейену, 1987б, 1988): а — Lepidodendron для стволов с сохранившейся корой (Aspidiaria, Bergeria, Aspidiopsis, Krorria для стволов, в разной степени утративших кору), Stigmaria для корней, Lepidocarpon для женских стробилов (показан общий вид и разрез мегаспорангия), и микроспора Lycospora, извлеченная из микроспорангия Lepidostrobus; б — все эти названия в некоторых случаях обозначают одно и то же растение, части которого в разных сочетаниях неоднократно были найдены в призизненной связи, как это показано линиями (пунктир — менее надежные свидетельства прижизненной связи).
Относительно некоторых случаев этого рода утверждается, что они затрагивают и принцип типа. Иногда резоны таких утверждений трудно понять (как в случае отказа типифицировать ихнотаксоны, т.е. таксоны системы ископаемых следов, в версии МКЗН, 1988, ст. 66), в других случаях причины подробно обсуждаются. А.Г. Пономаренко (1985) установил не типифицированные формальные таксоны, определяемые только признаками их диагноза («Род [Flichea Handlirsch.—А.Р.] понимается как формальное объединение изолированных надкрылий жуков без отчетливых борозд на верхней стороне и с заметной вырезкой посредине наружного края», с. 76]). Как объяснил мне автор, причиной явилось упорное стремление некоторых систематиков синонимизировать и таким образом ликвидировать паратаксоны, сколь произвольной ни была бы такая синонимизация, и в частности упомянутый выше пример с родом Laberius Kieffer. Не типифицированные таксоны Пономаренко — это, по существу, те же морфотаксоны, но искусственно детипифицированные и тем самым выведенные из-под юрисдикции кодекса номенклатуры (поскольку кодекс регламентирует применение названия ссылкой на тип, см. - выше). Детипификация действительно предотвращает необоснованную синонимизацию, но проблема в том, что она исключает всякую синонимизацию и поэтому оказывается лекарством хуже, чем сама болезнь. Мне кажется, что понимание целей и функций паратаксономии будет лучшей защитой от произвольной синонимизации.
Последний и самый бедный содержанием вид паратаксона это сборный таксон (collective taxon). Это паратаксон, обычно в ранге рода, который отнесен к определенному старшему таксону, но в отличие от морфотаксона не может быть выделен в специальную систему, параллельную основной. По существу, виды в сборных группах — это просто виды неясного положения (species incertae sedis). Например, название «Cercaria O.F. Muller, 1773 установлено для рода червей, и многие авторы в XIX в. обращались с ним как с названием номинального рода, как если бы его типовым видом был C. lemna O.F. MUller, 1773. В настоящее время Cercaria употребляется как название сборной группы для личинок трематод, которые не могут быть с уверенностью отнесены к нормальным родам» (МКЗН, 2000, ст. 67.14). Различие между таксонами неясного положения и видами сборного рода состоит только в том, что видовой эпитет не вправе появляться в одиночку, без названия рода. Поэтому для таких видов приходится вводить название сборной группы, которое имеет вид родового названия, но выполняет роль названия того старшего таксона, к которому автор относит виды, включенные в данный сборный род. Так, Cercaria в современном понимании — это эквивалент отряда трематод, Laberius в интерпретации Й. Кифера (Kieffer, 1914) — эквивалент Dryinini + Gonatopodini, а Dryinus в интерпретации М. Ольми (Olmi, 1984) — эквивалент Dryinidae.
Сборные таксоны считаются детипифицированными, но это утверждение ошибочно. Приведенное выше определение означает, что сборный таксон по характеристике и объему совпадает с соответствующим старшим таксоном, и его члены, подобно ортотаксонам, вводятся в систему в конечном счете ссылкой на номенклатурный тип последнего. Виды Cercaria определимы как таковые просто в силу того, что достаточно сходны с личинками типового вида трематод. А то, что типовой вид трематод пока формально не зафиксирован, несущественно: я не сомневаюсь, что старшие таксоны будут типифицированы и в зоологии. Другой пример — сборный род Carabilarva Ponomarenko, предложенный для мезозойских личинок жуков надсемейства Caraboidea, не определимых до семейства (А.Г. Пономаренко, 1985). Поэтому типовым видом рода является тип надсемейства Carabus granulatus Linnaeus, 1758. Следовательно, Carabilarva является младшим объективным синонимом рода Carabus, но не подлежит синонимизации с ним, пока остается паратаксоном.
Последний пример означает существование еще одной формы множественного подчинения в номенклатуре, помимо уже обсуждавшегося случая, когда разные формы сохранности одного естественного таксона описываются в качестве разных морфотаксонов. Здесь уже один и тот же вид оказывается типовым для множества сборных групп. Действительно, род Саrаbus одновременно является типовым для семейства Carabidae (жуки-жужелицы), подсемейства Сагаbinае, трибы Сагаbіпі и т.д., а для далее неопределимых членов каждого из этих таксонов теоретически могут понадобиться свои сборные группы, да еще разные для личинок и различных изолированных частей тела взрослого жука. Легко представить себе, сколько сборных родов в результате могут оказаться основанными на одном и том же номенклатурном типе. При всей непривычности ситуации, она вряд ли может вызвать серьезные номенклатурные или таксономические проблемы, если мы будем понимать, для чего нужны сборные таксоны. Ведь никому не мешают роды и старшие таксоны неясного положения, почему же отношение к видам неясного положения должно быть иным?
Существует еще один важный аспект проблемы неполноценного таксона (таксона особого статуса). Таксон принадлежит этой категории просто потому, что он выделен по «плохим» признакам — плохим в сравнении с другими, нормальными таксонами. Если же других таксонов нет, тот же самый неполноценный таксон оказывается вполне полноценным. Если бы современных ручейников не существовало, ископаемые домики, вероятно, изучались бы вне всякой связи с ископаемыми ручейниками, и их система, основанная на тех же самых «плохих» признаках, считалась бы нормальной. Во многих группах насекомых современные виды ныне различаются практически только по признакам мужских гениталий, а их вымершие представители — в основном по жилкованию крыльев. Пока ископаемых относят к особым, вымершим родам, номенклатурных проблем не возникает, и с ископаемыми удается работать как с ортотаксонами. Там же, где вымерший вид попадает в один род (подрод) с современными, он часто оказывается в категории неполноценного. Слабым утешением здесь служит то обстоятельство, что этого же заслуживают и давно описанные современные виды, для которых еще не удалось описать строение гениталий.
Помимо условности квалификации таксона в качестве орто- или паратаксона, существуют сложности и другого рода. Хороший признак или плохой, это дело, во-первых, меры (велика ли разница), во-вторых, вкуса. Обычное дело, когда спор между систематиками сводится, в конечном счете, к тому, на какую систему признаков следует опираться, разрабатывая систему конкретной группы. Можно только удивляться, что в споре пока еще не возникают взаимные обвинения, что, дескать, система оппонента, основанная на таких-то признаках, полезна только в качестве частной системы паратаксонов, но не общей (основной) системы группы.
Все эти соображения позволяют заключить, что ортотаксон и паратаксон формально четко различны, но содержательной обособленности здесь нет. В зависимости от точки зрения, а тем более, как мы видели, от уровня наших знаний (пример Baissobius), таксон может переходить из одной категории в другую. По-видимому, к паратаксономии нужно относиться прагматически: если использование паратаксонов реально облегчает познание и инвентаризацию биоразнообразия, их нужно вводить и использовать. Если же выигрыш невелик, не нужно усложнять ситуацию введением параллельных, перекрывающихся систем организмов.
1.3. ЗАКЛЮЧЕНИЕ
Проблемы эволюционного крыла современной биологии, куда можно отнести те ее разделы, что исследуют историю и современное состояние биологического разнообразия, включая их теорию и методологию, связаны прежде всего с поисками парадигмы. Уже более столетия продолжается спор двух подходов — «элементаристской» («редукционистской») и «холистской» парадигм. Поразительно, но «размежевания» этих столь различных подходов не происходит, а продукты каждого из них сосуществуют в головах и печатной продукции едва ли не большинства ученых, о чем те обычно и не подозревают.
Элементаристской парадигме, редуцирующей изучаемую систему к ее элементам, соответствует популяционно-генетическая (синтетическая) теория эволюции, представляющая эволюционный процесс как динамику частот дискретных наследственных детерминант (генов) в популяциях в условиях отбора. Отражением этой динамики и является наблюдаемый процесс эволюции, и ее специфика определяется, во-первых, спецификой генного уровня организации, во-вторых, спецификой отбора. Соответственно, эволюционный процесс выступает как существенно равномерный, а единственным регулярным источником дискретности, делающим возможным разбиение продукта эволюции на таксоны, оказываются дивергенция и вымирание. Этой парадигме адекватна таксономическая концепция кладизма, где таксон возникает в ходе эволюции как продукт дивергенции и вводится в систему именно на этом основании, а последовательность дивергенций определяется с помощью синапоморфий.
Альтернативная холистская парадигма редуцирует эволюционирующую систему к целому и в частности к эпигенотипу, т.е. к системе построения живого организма в онтогенезе. В зеркале эпигенетической теории эволюции сложная, пронизанная переплетающимися взаимосвязями и взаимодействиями живая система предстает напряженным компромиссом противоречивых требований оптимизации разных адаптивных функций. Изменение такой системы оказывается существенно затрудненным, мало предсказуемым и более или менее скачкообразным, а специфика эволюционного процесса определяется прежде всего спецификой эпигенотипа и лишь в существенно меньшей степени — спецификой отбора. Из-за скачкообразности эволюционных изменений биологическое разнообразие, возникающее в ходе такой эволюции, оказывается дискретным, разбитым гиатусами на естественные отдельности, каковые и заслуживают использования в качестве таксонов общей системы. Соответственно, смысл таксономии, адекватной эпигенетической концепции и холистской парадигме, состоит в выявлении объективно существующей дискретности и прослеживании гиатусов.
Анализ доступного материала свидетельствует в пользу его соответствия скорее эпигенетической, чем синтетической концепции. Более важной, однако, представляется сама попытка различения концепций и методологий, используемых в анализе эволюционного процесса и его результатов, как восходящих к той или другой из двух конкурирующих парадигм. Представляется, что строгое различение двух парадигм и их следствий позволит не только и даже не столько сделать осмысленный выбор между ними, сколько избавиться от одновременного использования взаимно несовместимых принципов и подходов, т.е. от своего рода плюрализма в одной голове.
Сделанный вывод позволяет несколько продвинуться в выявлении корней и в более четком формулировании методологических концепций, используемых на поле эволюционного крыла биологии, и в частности в филогенетике, таксономии и таксономической номенклатуре. Однако вначале нам пришлось рассмотреть некоторые самые общие проблемы научного (и не только) анализа.
Филогенетические выводы, как любое научное исследование, опираются на (1) планирование предстоящего исследования, (2) наблюдение (включая эксперимент), (3) поиск аналогий, (4) создание гипотез о лежащих в основе видимой картины закономерностях и механизмах, (5) попытки фальсификации этих гипотез (преимущественно путем анализа следствий из них), и (6) оценки результатов этих попыток с помощью презумпций, с целью выбора наиболее перспективной гипотезы для дальнейшей работы.
Набор филогенетических презумпций включает презумпцию познаваемости филогенеза и ряд более частных презумпций, которые можно разделить на используемые в анализе групп и в анализе признаков. Анализ групп исследует отношения предков и потомков и опирается прежде всего на палеонтологическую презумпцию для групп. Презумпции, используемые в анализе признаков, далее делятся на две группы соответственно тому, включены ли они в анализ различий или сходств. Анализ различий — это поляризация трансформационных серий, т.е. определение плезиоморфных (исходных) и апоморфных (измененных) состояний признака. Здесь работают, из более важных, палеонтологическая презумпция для признаков, биогенетическая презумпция, презумпции аналогии, необратимости эволюции, функционального совершенства, сложности, рудиментов и сохраненного распространения признаков. Презумпции анализа сходств используются при решении вопроса, унаследовано ли данное сходство от общего предка или возникло независимо. Сюда относятся прежде всего презумпции парсимонии и взвешенного сходства.
Цель таксономии состоит в создании системы, чьи таксоны осмысленны для максимально широкого круга пользователей и, соответственно, наиболее гомогенны внутри себя и различны между собой. Из трех главных конкурирующих подходов кладизм опирается на синтетическую теорию эволюции, игнорирующую дискретность биоразнообразия. Поэтому кладистическая система учитывает только дивергенции и признает лишь таксоны, охарактеризованные синапоморфиями. Синапоморфия — не признак, а его интерпретация, поэтому кладистическая система не только игнорирует дискретность биоразнообразия, но и излишне гипотетична и привередлива в отборе признаков.
Филетика и фенетика опираются на эпигенетическую теорию эволюции и заняты прослеживанием гиатусов между таксонами. Фенетика использует доступные признаки как таковые и потому слишком близорука, это существенно эмпирический подход. Филетика занимает промежуточное положение между фенетикой и кладизмом. Она пытается отразить полный баланс сходств и различий, включая и те, что еще не изучены. Для этого филетика использует прогностические возможности филогенеза, который выступает в качестве эвристического метода контроля системы, построенной фенетическими методами. Филетика определяет таксон как монофилетический континуум и использует гиатусы для разграничения таксонов, а монофилию — как способ оценить, успешно ли было разграничение, или система требует доработки.
В основе таксономической номенклатуры лежат 9 принципов, из которых шесть представляют чистые соглашения для удобства и единообразия в использовании названий таксонов и независимы от используемого таксономического подхода. Остальные три принципа зависят от таксономии. Из них наиболее важен принцип типа, позволяющий стабилизировать названия таксонов, соответствующих понятию монофилетического континуума. Два других таксономически зависимых принципа — принципы иерархии и синонимии, из которых второй может быть нарушен при работе с дефектным таксономическим материалом, недостаточно охарактеризованным для его полноценной систематизации. В этом случае могут применяться таксоны неясного положения и паратаксоны разного рода, используемые в рамках специальных систем, параллельных основной системе организмов.
В отличие от принципов синонимии, видимо, не существует объективных причин отказываться от соблюдения принципов типа и иерархии, а тем более от таксономически независимых принципов номенклатуры.
ЛИТЕРАТУРА
Алексеев А.С., Дмитриев В.Ю., Пономаренко А.Г. 2001. Эволюция таксономического разнообразия. Экосистемные перестройки и эволюция биосферы. Вып. 5. М.: Геос. 126 с.
Аристотель. Биомедгиз. 1937. 220 с.
Бекетов А.Н. 1860. Гармония в природе // Русский вестник. № 30. С. 197-240, 534-558.
Белокрыс Л.С. 1984. Спирорбины причерноморского миоцена // Палеонтол. журн. № 2. С. 26-39.
Беляев Д.К. 1974. О некоторых вопросах стабилизирующего и дестабилизирующего отбора. История и теория эволюционного учения, Вып. 2. Л.: Наука: 76-84.
Берман Д.И. 2001. Беспозвоночные животные. Заключение // Холодные степи Северо-Восточной Азии / Берман Д.И. (ред.). Магадан: Ин-т биол. Проблем Севера СВНЦ ДВО РАН. С. 96-167.
Вахрамеев В.А., Котова И.З. 1977. Древнейшие покрытосеменные и сопутствующие им растения из нижнемеловых отлдожений Забайкалья // Палеонтол. журн. № 4. С. 101-109.
Вейсман А. 1905. Лекции по эволюционной теории. М.: Изд-во М. и С. Сабашниковых. Т. 1. 505 с.
Воробьева Э.И. 1992. Проблема происхождения наземных позвоночных. М.: Наука. 344 с.
Вялов О.С., Сукачева И.Д. 1976. Ископаемые домики личинок ручейников (Insecta, Trichoptera) и их значение для стратиграфии // Палеонтология и биостратиграфия Монголии. Тр. Совм. Сов.-Монг. палеонтол. экспед. Вып. 3. С. 169-230.
Грант В. 1980. Эволюция организмов. М.: Мир. 480 с.
Гриценко В.В., Креславский А.Г., Михеев А.В., Северное А.С., Соломатин В.М. 1983. Концепции вида и симпатрическое видообразование. М.: Изд-во МГУ 194 с.
Гродницкий Д.Л. 2000. Две теории биологической эволюции. Красноярск: Ин-т леса им. В.Н. Сукачева СО РАН. 180 с.
Гродницкий Д.Л. 2002. Две теории биологической эволюции. 2-е изд. Саратов: Научная книга. 160 с.
Дарвин Ч. 1991. Происхождение видов путем естественного отбора. СПб.: Наука. 540 с.
Деятельность межсекционного семинара по проблемам эволюции с октября 1965 г. по апрель 1966 г. 1967 // Бюл. Моск. о-ва испыт. природы. Отд. биол. Т. 72. Вып. 4. С. 136138.
Жегалло В.И., Каландадзе Н.Н., Кузнецова Т.В., Раутиан А.С. 2001. Судьба мегафауны Голарктики в позднем антропогене / Мамонт и его окружение: 200 лет изучения. М.: ГЕОС. С. 287-306.
Жерихин В.В. 1978. Развитие и смена меловых и кайнозойских фаунистических комплексов (трахейные и хелицеровые) // Тр. Палеонтол. ин-та АН СССР. Т. 165. М.: Наука. 200 с.
Жерихин В.В. 1987. Биоценотическая регуляция эволюции // Палеонтол. журн. № 1. С. 312.
Жерихин В.В. 1993. История биома тропических дождевых лесов // Журн. общ. биол. Т. 54. С. 659-666.
Жерихин В.В., Расницын А.П. 1980. Биоценотическая регуляция эволюции. Микро- и макроэволюция. Материалы симпозиума. Кяэрику. 2-5 сент. 1980. Тарту. С. 77-81.
Заварзин Г.А. 1987. Особенности эволюции прокариот // Эволюция и биоценотические кризисы / Л.П. Татаринов, А.П. Расницын (ред.). М.: Наука. С. 144-158.
Иванова Л.С. 1978. Сравнительное изучение партеногенетических долгоносиков (Coleoptera, СитшИо^ае) Сибири: Автореф. дис ... канд. биол. наук. Новосибирск: Биол. ин-т СО АН СССР. 18 с.
Иорданский Н.Н. 1977. Неравномерность темпов макроэволюции и ключевые ароморфозы // Природа. № 6. С. 36-46.
Иорданский Н.Н. 1979. Основы теории эволюции. М.: Просвещение. 190 с.
Клюге Н.Ю. 2000. Современная систематика насекомых. Часть I. Принципы систематики живых организмов и общая система насекомых с классификацией первичнобескрылых и древнекрылых. СПб.: Лань. 336 с.
Красилов В.А. 1969. Филогения и систематика // Проблемы филогении и систематики: Материалы симпоз. Владивосток. С. 12-30.
Курочкин Е.Н. 2006. Параллельная эволюция тероподных динозавров и птиц // Зоол. журн. Т. 85. С. 283-297
Любищев А.А. 1923. О форме естественной системы организмов // Изв. Биол. н.-и. ин-та при Пермск. ун-те. Т. 2. Вып. 3. С. 99-110. (см. также в кн.: Любищев А.А. Проблемы формы, систематики и эволюции организмов. М.: Наука. 1982. С. 24-36).
Любищев А.А. 1982. Проблемы формы, систематики и эволюции организмов. М.: Наука. 280 с.
Майр Э. 1974. Популяции, виды и эволюция. М.: Мир. 460 с.
Мамкаев Ю.В. 1968. Сравнение морфологических различий в низших и высших группах одного филогенетического ствола // Журн. общей биол. Т. 29. С. 48-55.
Мейен С.В. 1983. Сравнение принципов систематики современных и ископаемых организмов // Математика и ЭВМ в палеонтологии. Кишинев: Штиинца. С. 10-25.
Мейен С.В. 1987. Основы палеоботаники. М.: Недра. 402 с.
Мейен С.В. 1989. Введение в теорию стратиграфии. М.: Наука. 215 с.
Мейен С.В. 1990. Палеоботаническая таксономия и номенклатура: необходимость нового подхода // Теоретические проблемы палеоботаники. М.: Наука. С. 49-76.
МКЗН 1988: Международный кодекс зоологической номенклатуры. Изд. 3. Л.: Наука. 1988. 205 с.
МКЗН 2000: Международный кодекс зоологической номенклатуры. Изд. 4. СПб.: Наука. 2000. 221 с.
Назаров В.И. 1984. Реконструкция ландшафтов северо-востока Белоруссии в антропогене по палеоэнтомологическим данным // Тр. Палеонтол. ин-та АН СССР. Т. 205. М.: Наука. 96 с.
Новокшонов В.Г. 1997. Ранняя эволюция скорпионниц. М.: Наука: 140 с.
Пoнoмapeнкo А.Г., Pacницын А.П. 1971. O фeнeтичecкoй и филoгeнeтичecкoй cиcтeмax // Зooл. жypн. T. 50. № 1. С. 5-14.
Полянский В.И. 1956. О виде низших водорослей // Комаровские чтения. М.-Л.: Изд-во АН СССР. 73 с.
Полянский Ю.И. 1957. О внутривидовой дифференциации и структуре вида у простейших // Вестн. ЛГУ № 21. С. 45-64.
Пономаренко А.Г. 1985. Жесткокрылые из юры Сибири и западной Монголии //Юрские насекомые Сибири и Монголии. Тр. Палеонтол. ин-та АН СССР. Т. 211. М.: Наука. С. 4787.
Пономаренко Н.Г. 1981. Новые перепончатокрылые насекомые семейства Dryinidae из позднего мела Таймыра и Канады // Палеонтол. журн. № 1. С. 139-143.
Поппер К. 1983. Логика и рост научного знания. М.: Прогресс. 606 с.
Разумовский С.М. 1971. О происхождении и возрасте тропических и лавролистных флор // Бюлл. ГБС АН СССР. Т. 82. С. 43-51.
Расницын А.П. 1969. Происхождение и эволюция низших перепончатокрылых // Тр. Палеонтол. ин-та АН СССР. Т. 123. М.: Наука. 196с.
Расницын А.П. 1971. К вопросу о причинах морфофункционального прогресса // Журн. общей биол. Т. 32. № 5. С. 549-556.
Расницын А.П. 1975. Высшие перепончатокрылые мезозоя // Тр. Палеонтол. ин-та АН СССР. М.: Наука. Т. 147. 134 с.
Расницын А.П. 1986а. Инадаптация и эвадаптация // Палеонтол. журн. № 1. С. 3-7.
Расницын А.П. 1987. Темпы эволюции и эволюционная теория (гипотеза адаптивного компромисса) // Эволюция и биоценотические кризисы. М.: Наука. С. 46-64.
Расницын А.П. 1988а. Филогенетика / Современная палеонтология. Т. 1. М.: Недра. С. 480497.
Расницын А.П. 2002. Процесс эволюции и методология систематики // Тр. Русск. энтомол. о-ва. Т. 73. СПб. 107 с.
Расницын А.П. 2005. Избранные труды по эволюционной биологии. М.: КМК. 347 с.
Расницын А.П., Длусский Г.М. 1988. Принципы и методы реконструкции филогенезов // Меловой биоценотический кризис и эволюция насекомых. М.: Наука. С. 5-15.
Расницын С.П. 2000. Будущее энтомологической систематики // Занимательные очерки о деятельности и деятелях противочумной системы России и Советского Союза. Вып. 10. М.: Информатика. С. 189-196.
Раутиан А.С. 1988. Палеонтология как источник сведений о закономерностях и факторахэволюции // Современная палеонтология. T. 2. M.: Недра. C. 76-118.
Родендорф Б.Б. 1977. О рационализации названий таксонов высокого ранга в зоологии // Палеонтол. журн. № 2. С. 14-22.
Родендорф Б.Б., Жерихин В.В. 1974. Палеонтология и охрана природы // Природа № 2. С. 82-91.
Северцов С.А. 1941. Динамика населения и приспособительная эволюция животных. М.-Л.: Изд-во АН СССР. 316 с.
Симпсон Дж. 1948. Темпы и формы эволюции. М.: Изд-во иностр. лит. 358 с.
Татаринов Л.П. 1976. Морфологическая эволюция териодонтов и общие вопросы филогенетики. М.: Наука. 258 с.
Цвелев Н.Н.1993. Гибрридизация как один из факторов увеличения биологического разнообразия и геномный критерий рода у высших растений // Биологическое разнообразие: подходы к изучению и сохранению. СПб.: Зоол. ин-т РАН. С. 193-201.
Чайковский Ю.В. 2003. Эволюция // Ценологические исследования. Вып. 22. М.: Центр системных исследований - ИИЕТ РАН. 472 с.
Шапошников Г. X. 1961. Специфичность и возникновение адаптации к новым хозяевам у тлей (Homoptera, Aphidodea) в процессе естественного отбора (экспериментальное исследование) // Энтомол. обозр. Т. 40. № 4. С. 739-762.
Шапошников Г. X. 1965. Морфологическая дивергенция и конвергенция в эксперименте с тлями (Homoptera, Aphidinea) // Энтомол. обозр. Т. 44. № 1. С. 3-25.
Шапошников Г. X. 1966. Возникновение и утрата репродуктивной изоляции и критерий вида // Энтомол. обозр. Т. 45. № 1. С. 3-35.
Шапошников Г.Х. 1978. Динамика клонов, популяций и видов и эволюция // Журн. общ. биол. Т. 39. № 1. С. 15-33.
Шарп Д. 1910. Насекомые. СПб.: Брокгауз-Ефрон. 1054 с.
Шварц Е.А. 2004. Сохранение биоразнообразия: сообщества и экосистемы. М.: КМК. 112 с.
Шварц С.С. 1980. Экологические закономерности эволюции. М.: Наука. 278 с.
Шер А.В. 1997а. Природная перестройка в Восточно-Сибирской Арктике на рубеже плейстоцена и голоцена и ее роль в вымирании млекопитающих и становлении современных экосистем (Сообщение 1) / Криосфера Земли. Т. 1. № 1. С 21-29.
Шер А.В. 1997б. Природная перестройка в Восточно-Сибирской Арктике на рубеже плейстоцена и голоцена и ее роль в вымирании млекопитающих и становлении современных экосистем (Сообщение 2) // Криосфера Земли. Т. 1. № 2. С. 3-11.
Шишкин М.А. 1984. Индивидуальное развитие и естественный отбор // Онтогенез. Т. 15. № 2. С. 115-136.
Шишкин М.А. 1987. Индивидуальное развитие и эволюционная теория // Эвoлюция и биoцeнoтичecкиe кризисы. M.: Hayra. C. 76-124.
Шишкин М.А. 1988а. Эволюция как эпигенетический процесс // Coвpeмeннaя пaлeoнтoлoгия. T. 1. M.: Heдpa. C. 142-169.
Шишкин М.А. 1988б. Закономерности эволюции онтогенеза // Современная палеонтология. T. 1. M.: Недра. C. 169-209.
Шишкин М.А. 2006. Индивидуальное развитие и уроки эволюционизма // Онтогенез. № 37. С. 179-198.
Эшби У.Р. 1964. Конструкция мозга. М.: Мир.
Alberch P. 1980. Ontogenesis and morphological diversification // Amer. Zool. Vol. 20. P. 653667.
Ashlock P.D. 1971. Monophyly and associated terms // Syst. Zool. Vol. 20. P. 63-69.
Baer K.E., von. 1828. Uber Entwickelungsgeschichte der Thiere. Beobachtung und Reflexion. Bd. 1. Konigsberg: Gebr. Borntrager. 271 S.
Bonde N. 1984. Primitive features and ontogeny in phylogenetic reconstructions // Vidensk. Meddr. dansk naturh. Foren. Vol. 145. P. 219-236.
Brothers D.J. 1975. Phylogeny and classification of the aculeate Hymenoptera, with special reference to Mutillidae // Univ. Kansas Sci. Bull. Vol. 50. P. 483-648.
Carson H.L.T. 1975. The genetics of speciation at the diploid level // Amer. Natur. Vol. 109. P. 8392.
Cope E.D. 1904. The primary factors of organic evolution. Chicago: Open Court Publ. Co. 547 p.
Cracraft J. 1974. Phylogenetic models and classification // Syst. Zool. Vol. 23. P. 71-90.
Eldredge N., Gould S. J. 1972. Punctuated equilibria: An alternative to phyletic gradualism. // Models in Paleobiology / Schopf T.J.M. (ed.) San Francisco: Freeman. P. 82-115.
Eskov K.Yu. 1987. A new archaeid spider (Chelicerata: Araneae) from the Jurassic of Kazakhstan, with notes on the so-called «Gondwanian» ranges of recent taxa // Neues Jb. Geol. Palaeont. Abh. 175. P. 81-106.
Eskov K.Yu. 1992. Archaeid spiders from Eocene Baltic amber (Chelicerata: Araneida: Archaei- dae) with remarks on the so-called «Gondwanian» ranges of Recent taxa // Neues Jb. Geol. Palaeont. Abh. 185. P. 311-328.
Farrer D.R. 1990. Species and evolution in asexually reproducing independent fern gametophytes // Syst. Bot. Vol. 15. P. 98-111.
Farris J.S. 1979. The information content of the phylogenetic system // Syst. Zool. Vol. 28. P. 483519.
Farris J.S. 1983. The logical basis of phylogenetic analysis // Advances in cladistics / Platnick N.I., Funk V.A. (eds.) New York: Columbia Univ. Press. Vol. 2. P. 7-36
Ghiselin M.T. 1974. A radical solution of the species problem // Syst. Zool. Vol. 23. P. 536-544. Ghiselin M.T. 1981. Categories, life and thinking // Behavioral and Brain Sciences. Vol. 4. P. 269-313.
Ghiselin M.T. 1987. Species concepts, individuality and objectivity // Biology and Philosophy. Vol. 2. P. 127-143.
Gidley J.W. 1912. The Lagomorpha, an independent order // Science. Vol. 36. P. 285-286.
Gould S.J., Eldredge N. 1977. Punctuated equilibria: the tempo and mode of evolution reconsidered // Paleobiology. Vol. 3. P. 115-151.
Gressit J.L. 1978. Evolution of endemic Hawaiian cerambycid beetles // Pacif: Insects. Vol. 18. P. 137-167.
Hennig W. 1966a. Phylogenetic systematics. Urbana: University of Illinois Press. 263 p.
Hull D.L. 1983. Karl Popper and Plato's metaphor // Advances in cladistics / Platnick N.I., Funk V.A. (eds.). Vol. 2. P. 177-189.
Humphries C.J., Parenti L.R. 1986. Cladistic biogeography. Oxford Monographs in Biogeography. Vol. 2. Oxford: Clarendon Press. 98 pp.
Kieffer J. J. Bethylidae. 1914. Das Tierreich. Berlin, R. Fridlandar u. Sohn. Lief. 41. 595 S.
Kukalova-Peck J. 1978. Origin and evolution of insect wings and their relation to metamorphosis, as documented by the fossil record // J. Morphol. Vol. 15. P. 53-126.
Labandeira C.C., Beall B.S., Hueber F.M. 1988. Early insect diversification: evidence from a Lower Devonian bristletail from Quebec // Science. Vol. 242. P. 913-916.
Laicharting J.N., von. 1781. Verzeichniss und Beschreibung der Tyroler Insecten. Bd. 1. Zurich: Fuessley. xii+248 S.
Linnaeus C. 1751. Philosophia botanica in qua explicatur fundamenta botanica cum definitionibus partium, exemplis terminorum, observationibus rariotum, adjectis figuris aeneis. Stockholm. P. G. Kiesewetter. viii + 362 p.
Mayr E. Toward a new philosophy of biology: observations of an evolutionist. Cambridge, London: Belknap Press, 1988. xi + 564 p.
McCune A.R. 1990. Evolutionary novelty and atavism in the Semionotus complex: Relaxed selection during colonization of an expanding lake // Evolution. Vol. 44. P. 71-85.
Melville R.V. 1981. The International code of zoological nomenclature. Results of vote on proposals for substantive amendments (fifth installment) // Bull. Zool. Nomencl. Vol. 38. P. 30-48.
Melville R.V. 1981. International code of zoological nomenclature. Deferment of proposal to introduce provisions to regulate paranomenclature // Bull. Zool. Nomencl. Vol. 38. P. 166-167.
Meyen S.V., Traverse A. 1979. Proposal on Article 3: remove «form-genus» too // Taxon. Vol. 28. P. 595-598.
Mickevich M.F. 1978. Taxonomic congruence // Syst. zool. Vol. 27. P. 143-158.
Nelson G.J. 1978. Ontogeny, phylogeny, paleontology, and the biogenetic law // Syst. Zool. Vol. 27. P. 324-345.
Olmi M. 1984. Revision of Dryinidae // Mem. Amer. Ent. Inst, Ann. Arbor. Vol. 37. Pt. 1-2. 1913 p.
Osborn H.F. 1934. Aristogenesis: the creative principle in the origin of species // Amer. Natur. Vol. 68. P. 193-703.
Oster G., Alberch P. 1982. Evolution and bifurcation of developmental programs // Evolution.Vol. 36. P. 444-459.
Platnick N.I. 1979. Philosophy and the transformation of Cladistics // Syst. Zool. Vol. 28. P. 537546.
Plejel F., Rose G.W. 2002. Ceci n'est pas une pipe: names, clades and phylogenetic nomenclature // J. Zool. Syst. Evol. Research. Vol. 41. P. 162-174.
Poljansky G. I. 1977. Some aspects of the species in asexually reproducing Protozoa // Protozoology. Vol. 3. P. 17-23.
Queiroz K., de, Donoghue M.J. 1990a. Phylogenetic systematics or Nelson's version of cladistics? // Cladistics. Vol. 6. P. 61-75.
Queiroz K., de, Donoghue M.J. 1990b. Phylogenetic systematics and species revisited // Cladistics. Vol. 6. P. 83-90.
Queiroz K., de, Gauthier J. 1990. Phylogeny as a central principle in taxomony: phylogenetic definition of taxon names // Syst. Zool. Vol. 39. P. 307-322.
Rachootin S.P., Thomson K.S. 1981. Epigenetics, paleontology and evolution / Scudder G.G.E., Reveal J.E. (eds.) Evolution today. Pittsburg: Hunt Inst. P. 181-194.
Rasnitsyn A.P. 1981. A modified paranotal theory of insect wing origin // J. Morphol. Vol. 168. P. 331-338.
Rasnitsyn A.P. 1982. Proposal to regulate the names of taxa above the family group. Z.N.(S.) 2381 // Bull. Zool. Nom. Vol. 39. P. 200-207.
Rasnitsyn A.P. 1989. Vespida vs. Hymenoptera // Sphecos. Vol. 18. 8 p.
Rasnitsyn A.P. 1991. Typified names of higher taxa once again // Sphecos. Vol. 22. P. 5-7.
Rasnitsyn A.P. 1996a. Conceptual issues in phylogeny, taxonomy, and nomenclature // Contributions to Zoology. Vol. 66. P. 3-41.
Rasnitsyn A.P. 1996b. New Early Cretaceous Embolemidae (Vespida = Hymenopterea: Chrysi- doidea) // Norden B.B., Menke A.S. (eds.) Contributions on Hymenoptera and associated insects dedicated to Carl V. Krombein. Memoirs of the Entomological Society of Washington. Vol. 17. P. 183-187.
Rasnitsyn A.P. 2000b (1999). Taxonomy and morphology of Dasyleptus Brongniart, 1885, with description of a new species (Insecta: Machilida, Dasyleptidae) // Russian Entomol. J. Vol. 8. No. 3. P. 145-154.
Rasnitsyn A. P. 2006. Ontology of evolution and methodology of taxonomy // Paleontological Journal. Vol. 40. Suppl. 6. P. S679-S737.
Rasnitsyn A.P., Golovatch S.I. 2004. The identity of Phryssonotus burmiticus (Cocerell , 1917) (Diplopoda, Polyxenida, Synxenida) in Cretaceous amber from Myanmar // J. syst. Paleontol. Vol. 2. No. 2. P. 153-157.
Rasnitsyn A.P., Ross A.J. 2000. A preliminary list of arthropod families present in the Burmese amber collection at The Natural History Museum, London / Ross A.J. (ed.) The history, geology, age and fauna (mainly insects) of Burmese amber, Myanmar // Bulletin of The Natural History Museum, Geology Series. Vol. 56. No. 1. P. 21-24.
Rausch R.L. 1982. Cestodes in mammals: the zoogeography of some parasite-host assemblages // Mem. Mus. nation. hist. natur. Ser. A. Vol. 23. P. 179-183.
Rotondo G.M., Springer V.G., Scott G.A.J., Schlanger S.O. 1981. Plate movement and island integration — a possible meshanism in the formation of endemic biotas, with special reference to the Hawaiian Islands // Syst. Zool. Vol. 30. P. 12-21.
Saether O.A. 1986. The myth of objectivity — post-Hennigian deviations // Cladistics. Vol. 2. P. 1-13.
Saether O.A. 1979. Underlying synapomorphies and anagenetic analysis // Zoologica Scr. Vol. 8. P. 305-312.
Sarjeant W. A. S., Kennedy W. J. 1973. Proposal of a Code for the nomenclature of trace-fossils // Canad. J. Earth Sci. Vol. 10. P. 460-475.
Schram F. R. 1983. Method and madness in phylogeny / Schram F. R. (ed.). Crustacean Phytogeny. Rotterdam: Balkema. P. 331-350.
Shear W.A., Bonamo P.M., Griedson J.D., Rolf W.D.I., Smith E.L., Norton R.A. 1984. Early land animals in North America: Evidence from Devonian age arthropods from Gilboa, New York // Science. Vol. 224. P. 492-494.
Shedrinsky A., Wampler Th.P., Grimaldi D. 2001. Burmese amber revisited (data on chemical composition of newly excavated material) / Krzeminska E., Krzeminski W. (eds.). Fossil insects. Second International Congress on Palaeoentomology, 5-9 September, 2001, Krakow, Poland. Abstracts volume. Institute for systematics and evolution of animals, Polish Academy of Science, Krakow. P. 66-67.
Sher A.V., Kuzmina S.A., Kiselyov S.V., et al. 2006. QUINSIB. The Database on Quaternary Insects of Northeastern Siberia: Preliminary Version 3 (04.06).
Smuts J.C. 1987. Holism and evolution. Cape Town, N & S Press. 361 p.
Starobogatov Ya.I. 1991. Problems in the nomenclature of higher taxonomic catgories // Bull. Zool. Nom. Vol. 48. P. 6-18.
Sundberg P., Pleijel F. 1994. Phylogenetic classification and the definition of taxon names // Zoologica Scr. Vol. 23. P. 19-25.
Suno-Uchi N., Sasaki F., Chiba S., Kawata M. 1997. Morphological stasis and phylogenetic relationships in tanpole shrimps, Triops (Crustacea: Notostraca) // Biol. J. Linnean Soc. Vol. 61. P. 439-457.
Tasch P. 1969. Branchiopoda. Treatise on Invertebrate Paleontology // Arthropoda. Vol. 4. 1. Pt. R. P. 128-191.
Tuxen S.L. 1963. Art- und Gattungsmerkmal bei den Proturen // Entomol. medd. Vol. 32. P. 84-98.
Tuxen S.L. 1967. Australian Protura, their phylogeny and zoogeography // Ztschr. zool. Syst. und Evolutionforsch. Vol. 5. P. 1-53.
Van Valen L. 1973. A new evolutionary law // Evol. Theory. Vol. 1. P. 1-30.
Van Valen L. 1976. Energy and evolution // Evol. Theory. Vol. 1. P. 179-229.
Van Valen L. 1982. Integration of species: stasis and biogeography // Evol. Theory. Vol. 6. P. 99112.
Wiley E.O. 1979. An annotated Linnean hierarchy, with comments on natural taxa and competing systems // Syst. Zool. Vol. 28. P. 308-337.
Wilson R.W. 1989. Rodent origin. // Papers on fossil rodents / Black C.C., Dawson M.R. (eds.). / Sci. Ser. Los Angeles Mus. Vl. 33. P. 3-6.
Woese C.R. 1987. Microevolution in the microscopic world // Patterson C. (ed.). Molecules and morphology in evolution: conflict or compromise? Cambridge, etc.: Cambridge University Press. P. 177-202.
Zherikhin V.V. 1999. Cladistics in palaeontology: problems and constraints. Proc. First Internat. Palaeoentomol. Conf., Moscow, 1998. AMBA projects Internat., Bratislava. P. 193-199.
Zherikhin V.V. 2002a. Insect trace fossils // Rasnitsyn A.P., Quicke D.L.J. (eds.). History of Insects. Dodrecht etc., Kluwer Academic Publishers. P. 303-324.
|
А.П.Расницын. Теоретические основы эволюционной биологии // В.В.Жерихин, А.Г.Пономаренко, А.П.Расницын. Введение в палеоэнтомологию. М.: КМК. 2008. 371 с
1.2.2.3. Филетика (продолжение)
1.2.3. ТАКСОНОМИЧЕСКАЯ НОМЕНКЛАТУРА
1.2.3.1. Таксономически независимые принципы
1.2.3.2. Таксономически зависимые принципы
|
|