Студенческие работы после практики II курса - 2010
Студенческие работы после практики II курса - 2011
Студенческие работы после практики II курса - 2012 (I часть)
Студенческие работы после практики II курса - 2012 (II часть)
Студенческие работы после практики II курса - 2013
Студенческие работы после практики II курса - 2014
Студенческие работы после практики II курса - 2015
Студенческие работы после практики II курса - 2016
Студенческие работы после практики II курса - 2017
ИЗУЧЕНИЕ ВОЗМОЖНОСТИ ОПРЕДЕЛЕНИЯ ФОРМ ЗЕЛЕНЫХ ЛЯГУШЕК ПО ПРИЗНАКАМ ВНЕШНЕЙ МОРФОЛОГИИ И ОКРАСКИ
Дедух Д.В., Зарубенко Е. С.
Харьковский национальный университет имени В. Н. Каразина, биологический факультет
пл. Свободы 4, г. Харьков, 61077, Украина
Rana esculenta complex—группа рода зеленых лягушек, состоящая из родительских видов и их гемиклональных гибридов. В окрестностях биостанции ХНУ совместно обитают как представители одного из родительских видов – Rana ridibunda, так и различных (в частности, как ди-, так и триплоидных) гибридов — Rana kl. esculenta (Шабанов и др., 2006).
Собрали 73 экземпляра половозрелых зеленых лягушек из поймы реки Северский Донец. Лягушек ловили сачком в темное время суток, собирая у берега и в зарослях высшей водной растительности с помощью фонаря. После выполнения работы изученные лягушки были выпущены в место отлова.
Из изученных нами лягушек 15 особей обладали «типичными» признаками R. ridibunda: прямой пяточный бугор и отсутствие желтого узора на бедрах. 17 особей оказались «типичными» R. esculenta: у них был высокий пяточный бугор и желтый узор на бедрах. Оставшиеся лягушки (n=41) обладали комбинациями признаков, характерных и для R. ridibunda, и для R. esculenta. Применяли дискриминантный анализ, в ходе которого сравнили «нетипичных» особей с «типичными» R. ridibunda и R. esculenta по признакам, используемым различными авторами для различения форм зеленых лягушек. Это отношение длины тела к длине голени, отношение длины пяточного бугра к длине голени, цвет спинной стороны тела (темно-зеленый, зеленый, светло-зеленый), форма пятен (меньше глаза, больше глаза), частота пятен (больше фона, одинаковое количество с фоном, меньше фона).
В результате дискриминантного анализа признаки «нетипичных» лягушек образовали облако рассеяния, внутри которого располагались признаки «типичных» R. ridibunda и R. esculenta. Это означает, что пользуясь данным набором признаков невозможно определить принадлежность к определенной форме всех лягушек из изученного местообитания.
Данная работа выполнена как УИРС в рамках учебно-полевой практики по зоологии позвоночных. Авторы выражают благодарность руководителям работы ст. преп. Кравченко М.А. и доц. Шабанову Д. А., а также благодарят А.А. Атемасова за ценную консультацию.
Дедух Д.В., Зарубенко Е. С. (научные руководители – Кравченко М.А., Шабанов Д. А.) Изучение возможности определения форм зеленых лягушек по признакам внешней морфологии и окраски// «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харьков: СПД ФО Михайлов Г.Г., 2008. - с. 364-365.
ИЗУЧЕНИЕ ФЛУКТУИРУЮЩЕЙ АСИММЕТРИИ РИСУНКА У RANA ESCULENTA COMPLEX
Деряженцева А.А., Дьяконова И.В., Микос И.Г.
Харьковский национальный университет имени В.Н. Каразина
пл. Свободы, 4, г. Харьков, 61077, Украина
e-mail: bell-iren@mail.ru
Одной из мер оценки устойчивости развития является флуктуирующая асимметрия (ФА) (Гелашвили Д.Б. и др., 2004). В Rana esculenta complex зарегестированна межвидовая гибридизация, клональное наследование, естественная полиплоидия (Plötner, 2005; Шабанов и др., 2006). Это делает интересным сравнение двух разных форм лягушек из одного местообитания по характерным для них уровням ФА.
Исследовано 149 зеленых лягушек, относящихся к Rana esculenta complex. Все особи были пойманы в реке Северский Донец в окрестностях биостанции ХНУ имени Каразина (с. Гайдары Змиевского р-на Харьковской области). На основании морфофизиологических характеристик лягушки распределялись к той или иной форме. Из исследованных экземпляров 116 обладали типичными для формы Rana ridibunda признаками: низким и косым пяточным бугром, отсутствием желтого узора на бедрах, темными резонаторами у самцов; 24 особи были отнесены к форме Rana esculenta на основании таких критериев: высокий пяточный бугор, желтый цвет на задних конечностях, светло-серая окраска резонаторов у самцов. 9 особей мы не смогли уверенно отнести ни к одной из форм, что, возможно, связанно с их полиплоидностью (морфофизиологические признаки формировались под воздействием сразу нескольких геномов).
Для определения характера асимметрии мы регистрировали следующие признаки: характер дорзомедиальной полосы (ДМП) (симметричная, слегка асимметричная, сильно асимметричная), количество пятен-полос на правой и левой нижних конечностях от бедра до конца голени, а также количество пятен вдоль ДМП слева и справа. Чтобы определить могут ли эти признаки использоваться, как мера ФА, мы провели сравнение групп по этим критериям. Оказалось, что мерой ФА может быть абсолютное («по модулю») значение разницы между количеством пятен слева и справа вдоль ДМП, менявшееся от 0 до 3.
Среднее значение ФА по этому признаку (т.е разница между количеством пятен вдоль ДМП справа и слева) для R. esculenta составляо 0.87, а для R. ridibunda составляло 0.67 (при сравнении по критерию Стьюдента р=0.18). Таким образом, различие гибридов и особей родительской формы недостоверно (возможно из-за недостаточного объема выборки), однако, несмотря на полученные показатели, была замечена тенденция к большей асимметрии гибридных особей.
Было выяснено, что у самцов Rana esculenta complex асимметрия выражена сильнее, что, предположительно, обусловлено более слабой зарегулированностью их развития по сравнению с самками.
Таким образом, мы предполагаем, что количество пятен вдоль ДМП может быть использовано для оценки ФА различных (ди- и триплоидных) гибридов.
Данная работа выполнена в рамках УИРС учебно-полевой практики по зоологии позвоночных под руководством ст. преподавателя Кравченко М.А. и доцента Шабанова Д.А.
Деряженцева А.А., Дьяконова И.В., Микос И.Г. (научные руководители – Кравченко М.А., Шабанов Д. А.) Изучение флуктуирующей асимметрии рисунка у Rana esculenta complex // «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харьков: СПД ФО Михайлов Г.Г., 2008. - с. 365-366.
СКЕЛЕТОХРОНОЛОГИЧЕСКОЕ ОПРЕДЕЛЕНИЕ ВОЗРАСТА ПРЕДСТАВИТЕЛЕЙ ЗЕЛЕНЫХ ЛЯГУШЕК RANA ESCULENTA COMPLEX ИЗ ХАРЬКОВСКОЙ ОБЛАСТИ
Усова Е.Е.
Харьковский национальный университет имени В.Н. Каразина кафедра зоологии и экологии животных пл.Свободы 4, г. Харьков, 61077, Украина
e-mail: е_usova@mail.ru
Группа зеленых лягушек — Rana esculenta complex — одна из интереснейших для изучения групп животных Харьковской области. В нее входят два родительских вида, а также различные ди-, три- и даже тетраплоидные формы их гибридов (Plotner, 2005). Сосуществование гибридов с родительскими видами поддерживается благодаря гемиклональному наследованию (передаче гибридами определенных геномов клонально, без рекомбинации), что представляет значительный научный интерес. При совместном обитании ди- и триплоидных гибридов возникает возможность сравнения их жизнеспособности. Адекватным методом для этого является скелетохронологическое определение возраста и скорости роста.
Нами изучено 62 особи зеленых лягушек, хранящихся в коллекции ЗИН РАН (Санкт-Петербург), Музея природы ХНУ и кафедры зоологии и экологии животных ХНУ. Изучено 50 особей Rana esculenta (гибридов); 9 — Rana ridibunda и 3 — Rana lessonae. Отлов лягушек проводился в Харьковской области с 2004 по 2007 г. Видовая принадлежность и состав геномов в генотипе для большинства изученных особей определялся методом проточной ДНК-цитомерии в ИНЦ РАН (Санкт-Петербург) С.Н. Литвинчуком и Ю.М. Розановым. Часть особей (представители R. lessonae) определены морфологически по комплексу признаков (Лада, 1995; Шабанов и др., 2006).
Для определения возраста были использованы срезы декальцинированных фаланг пальцев, которые затем окрашивали гематоксилином Эрлиха (Ромейс, 1953). Срезы были получены при помощи замораживающего микротома. Определение возраста мы проводили путем подсчитывания числа линий склеивания в периостальной зоне кости с учетом возможной резорбции первоначально отложенных слоев. Как правило, первая линия склеивания резорбируется полностью, а вторая — частично. Возраст особей можно высчитывать по формуле: возраст = число полностью видимых (не затронутых резорбцией) линий склеивания + 2.
Мы установили, что в Харьковской области возраст половозрелых зеленых лягушек, пойманных на нересте, находится в пределах от 4-х до 10-ти лет, с преобладанием 5-ти и 6-ти летних особей. У изученных гибридных особей зарегистрировано существенное разнообразие по темпам их роста. Наибольший возраст имеют не самые крупные особи, а самые крупные особи не являются самыми старыми. К примеру, зарегистрированы особи с длиной тела более 740 мм. в возрасте 5 лет и 8-10 летние с длиной тела не превышающей 689 мм. Нами не были зарегистрированы существенные отличия в скорости роста между диплоидными и триплоидными гибридами, что является предварительным свидетельством нормальной жизнеспособности триплоидных гибридов.
В ходе дальнейших исследований мы планируем увеличить число изученных лягушек и определить динамику роста отдельных особей. При изучении лягушек из одного местообитания (поймы Северского Донца в окрестностях биостанции ХНУ) сравнение скорости роста и продолжительности жизни представителей различных форм даст возможность оценить их жизнеспособность и вклад в воспроизводство гемиклональной популяционной системы.
Работа выполнена под руководством доцента кафедры зоологии и экологии животных ХНУ, к. б. н. Шабанова Д.А. Автор выражает благодарность за помощь в работе Шабановой А.В., Коршунову А.В. и Шабанову Д.А.
Усова Е.Е. (научный руководитель – Шабанов Д. А.) Скелетохронологическое определение возраста представителей зеленых лягушек Rana esculenta complex из Харьковской области// «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харьков: СПД ФО Михайлов Г.Г., 2008. - с. 396-397.
ОСОБЕННОСТИ СТРУКТУРЫ ФОЛИДОЗА ГОЛОВЫ У ПРЫТКИХ ЯЩЕРИЦ (LACERTA AGILIS) ОКРЕСТНОСТЕЙ БИОСТАНЦИИ ХНУ
Садовничая М. А., Чусова О. А.
Харьковский национальный университет имени В.Н.Каразина, биологический факультет
пл. Свободы 4, г. Харьков, 61077, Украина
Определение особенностей фолидоза (щиткования) широко применяется в систематике рептилий и анализе их популяционной структуры. При этом учитывается количество, форма и взаиморасположение отдельных щитков головы и тела.
В работе рассмотрены особенности щиткования лобной части головы 16 особей прыткой ящерицы (Lacerta agilis), собраных в НПП «Гомолышанские леса» (Змиевской р-н, Харьковская обл.) и в окрестностях биостанции ХНУ (Змиевской р-н, с. Гайдары). При описании фолидоза головы прытких ящериц за основу принята сводная таблица разнообразия морфологических признаков передненосовых (пн), предлобных (пл), лобноносового (лн) и других щитков, составленная по результатам исследования географической изменчивости прытких ящериц в рамках бывшего СССР (Яблоков, Ларина, 1985).
При обследовании собранных экземпляров отмечены признаки, не включенные в указанную сводку. Первый из них - вариант строения передненосовых щитков, при котором между ними спереди относительно глубоко вклинивается межносовой (мн) щиток (рис.1). Такое состояние наблюдалось у 5 из 16 изученных нами особей.
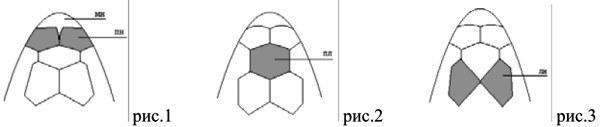
Вторым зарегистрированным признаком, который не входил в опубликованную схему описания, является вариант строения лобноносового щитка, при котором он имеет форму правильного шестиугольника. Обращенные переднелатерально и заднелатерально вершины этого щитка соединяются с передними вершинами предлобных щитков под углом примерно 110° (рис.2). Эта особенность является наиболее распространенным вариантом строения лобноносового щитка для изученной выборки ящериц, и наблюдалась у 15 особей из 16 изученных.
Наконец, отмечен новый вариант строения предлобных щитков, при котором они имеют ромбовидную форму и соприкасаются лишь в одной точке в своей задней части (рис.3). Эта особенность фолидоза наблюдалась у одной из рассмотренных нами особей.
В ходе работы не было зарегистрировано связи между окраской животных и особенностями их щиткования.
Вероятно, наличие «необычных» морфологических признаков фолидоза головы характеризует особенности рассмотренной нами выборки ящериц. В дальнейшем эти признаки могут быть использованы для фенетического анализа популяций прытких ящериц.
Работа выполнена на биологической станции ХНУ им. В.Н.Каразина под руководством ст. преп. Кравченко М.А. и доц. Шабанова Д.А., которым мы выражаем искреннюю благодарность.
Садовничая М. А., Чусова О. А. Особенности структуры фолидоза головы у прытких ящериц (Lacerta agilis) окрестностей биостанции ХНУ// «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харьков: СПД ФО Михайлов Г.Г., 2008. - с. 398-399.
К РАЗРАБОТКЕ МЕТОДОВ ФОРМАЛИЗОВАННОЙ ОЦЕНКИ ЦЕЛЕСООБРАЗНОСТИ ОХРАНЫ ПРИРОДНЫХ ОБЪЕКТОВ
Кравченко М. А.
Харьковский национальный университет имени В.Н. Каразина, биологический факультет, кафедра зоологии и экологии животных
пл. Свободы, 4, г. Харьков, 61077, Украина
e-mail: m_kravchenko@inbox.ru
В условиях быстрого изменения биосферы, вызванного как антропическими воздействиями, так, возможно, и другими причинами, актуальной становится проблема выбора природных объектов, приоритетных с точки зрения их охраны. В данной работе мы рассмотрим некоторые принципы, определяющие основания для охраны природных объектов (популяций, видов, биогеоценозов и т.п.).
Какой-либо объект можно охранять как ради его полезности, так и в силу этических причин, морального долга. Оценкой полезности объекта может быть его стоимость, а оценкой этических оснований для его охраны – его ценность. К примеру, концепция рационального использования природных ресурсов оценивает потенциальные объекты охраны с точки зрения их полезности (актуальной или потенциальной) (Марушевський, 2008), а т.н. «экологическая этика» и другие натуроцентрические концепции рассматривают в первую очередь этические основания для охраны природы (Рьюз, 1992; Борейко, 2004). По нашему мнению, при выборе объектов для охраны необходимо учитывать как их стоимость, так и их этическую ценность. Концепция, на основании которой должен осуществляться такой выбор, должна позволять хотя бы качественно оценивать эти параметры.
Этические основания для охраны объектов связаны с их уникальностью и возможностью их существования в долгосрочной перспективе, потенциальным бессмертием (Кравченко, Шабанов, 2006). В соответствии со сказанным, мы предлагаем следующий способ сравнения оснований для охраны тех или иных объектов

Сама необходимость охраны определенного объекта является следствием той или иной опасности, угрожающей его существованию. В подавляющем большинстве случаев охрана не может полностью ликвидировать угрозу гибели объекта, а лишь уменьшает ее вероятность. Эффективность охраны может быть оценена как снижение вероятности гибели объекта благодаря охранным мероприятиям.
Оценить уникальность объекта можно по вероятности возникновения его аналога, идентичного данному объекту по существенным для нас свойствам. Мерой потенциального бессмертия объекта может быть вероятный срок его существования в том случае, если он сохранится в краткосрочной перспективе (той, на которую планируются мероприятия по его охране).
Изложенный подход объединяет экономические основания для охраны объектов с формализованной оценкой их этической ценности. Кроме двух экономических оценок (стоимости объекта, т.е. возможной выгоды, полученной от его существования, а также стоимости мероприятий по его охране), требующих привлечения экономистов, в приведенной формуле используются вероятностные величины, связанные с особенностями возникновения и гибели охраняемых объектов. Оценка таких вероятностей должна быть задачей специалистов по изучению данной категории систем. Она может производиться как на основании статистической обработки эмпирических данных, описывающих судьбу аналогичных объектов, так и на основании результатов моделирования соответствующих природных систем.
Автор выражает благодарность доценту, к.б.н. Д.А. Шабанову, под руководством которого выполнена данная работа.
Кравченко М. А. К разработке методов формализованной оценки целесообразности охраны природных объектов // «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харьков: СПД ФО Михайлов Г.Г., 2008. - с. 442-443.
ЧТО НОВОГО МОЖНО УЗНАТЬ О ЗЕЛЕНЫХ ЛЯГУШКАХ ПРИ ПОМОЩИ ИМИТАЦИОННОГО МОДЕЛИРОВАНИЯ?
1 Кравченко М.А., 2 Луцик А.А., 1 Шабанов Д.А.
Харьковский национальный университет имени В.Н. Каразина,
1 биологический факультет, кафедра зоологии и экологии животных
2 механико-математический факультет, кафедра высшей математики и информатики
e-mail: 1 m_kravchenko@inbox.ru; d.a.shabanov@gmail.com, 2 aalutsyk@gmail.com
Описывая разнообразие живого мира, мы упрощаем разнообразие наблюдаемых явлений до набора типичных схем. Так, принято считать, что организмы в природе принадлежат к определенным видам и входят в состав популяций, состоящих из особей этого вида. Подавляющее большинство раздельнополых диплоидных организмов передают своим потомкам геномы, являющиеся результатом рекомбинации тех двух геномов, которые каждый из этих организмов получил от родителей. Однако из описанных схем существует немало исключений. Одно из них - гибридизация с образованием гемиклональных гибридов, передающих потомкам геномы одного из двух родительских видов в чистоте. В частности, такая гибридизация характерна для зеленых лягушек.
К группе европейских зеленых лягушек принадлежат два родительских вида: прудовая, Pelophylax lessonae (Camerano, 1882) и озерная, Pelophylax ridibundus (Pallas, 1771) лягушки, а также их гибриды, названные съедобными лягушками, Pelophylax esculentus (Linnaeus, 1758). Названия этих форм даны в соответствии с результатами последней ревизии системы амфибий (Frost et al., 2006 и более ранние работы). Более известны их традиционные названия - Rana lessonae Camerano, 1882, Rana ridibunda Pallas, 1771 и Rana esculenta Linnaeus, 1758. Почему гибридная форма носит имя, подобное видовому? Гибридные лягушки могут длительно существовать без повторных скрещиваний родительских видов и образовывать популяции, в которых они успешно воспроизводятся в течение многих поколений.
Описывая воспроизводство зеленых лягушек, мы обозначаем геном P. lessonae (состоящий из 13 хромосом) символом L, а геном P. ridibundus (тоже 13 хромосом) – R. Гемиклональное наследование у P. esculentus выражается в том, что только один из геномов диплоидных гибридных лягушек переходит в гаметы. Такой геном называется клональным; мы предлагаем обозначать это обстоятельство, заключая символ соответствующего генома в скобки. Второй геном у диплоидных гибридов элиминируется на той или иной стадии гаметогенеза. Например, во многих местообитаниях Восточной Украины совместно обитают и размножаются P. ridibundus и P. esculentus. Такие и подобные им совокупности зеленых лягушек не корректно называть «популяциями», так как их составляют животные, не принадлежащие к одному виду. Мы предлагаем называть такие биосистемы гемиклональными популяционными системами – ГПС. Особи родительского вида производят гаметы с рекомбинантными геномами этого вида, а гибриды – только гаметы с клональным геномом (L). В результате все потомство от скрещивания родительского вида и гибридов с клональным геномом другого вида оказывается гибридным: RR × (L)R → (L)R. Клональный геном при этом передается из поколение в поколение без рекомбинации.
Часто ГПС имеет намного более сложный состав. В окрестностях биостанции ХНУ (с. Гайдары Змиевского р-на Харьковской обл.) обитает ГПС, в состав которой входят лягушки RR, (L)R, L(R), (L)(R), LLR, LRR, а среди ювенильных особей встречаются также LL и LLRR. У три- и тетраплоидных гибридов клональные геномы в скобки не взяты, т.к. их гаметогенез полностью не изучен; особи, обозначенные как (L)(R), одновременно производят гаметы, несущие геномы P. lessonae и P. ridibundus в чистоте.
ГПС – относительно новая категория биосистем для современной науки. Процессы, поддерживающие их воспроизводство, остаются в существенной степени неизученными. Для их изучения мы применили имитационное моделирование с использованием мультиагентной компьютерной модели. Одним из авторов данного сообщения (А.А. Луциком) под руководством доц. М.В. Владимировой и проф. Г.Н. Жолткевича была создана программа Batrahometrics, моделирующая трансформации ГПС зеленых лягушек. С помощью этой модели можно описать начальное состояние ГПС (включающее численность особей определенных форм и возрастов); указать правила, по которым состояние ГПС в год n определяет его же в год n+1; определить распределение вероятностей состояния, в котором эта ГПС окажется по прошествии того или иного времени.
К числу упрощений, принятых при построении модели, относится постоянство численности нерестового стада (1000 особей) и идентичность свойств особей, относящихся к определенной форме (определяемой набором ее геномов). В зависимости от набора геномов особь характеризуется определенной привлекательностью для партнеров, плодовитостью, выживаемостью до наступления половозрелости и после созревания. На каждом шаге работы модели, соответствующем одному году, происходит вероятностное образование пар и вероятностная гибель имевшихся особей. Отслеживая передачу клональных и рекомбинантных геномов из поколения в поколение, программа Batrahometrics отслеживает изменение состава ГПС во времени.
На входе каждого отдельного эксперимента (моделирования трансформаций определенной ГПС в течение определенного времени) учитывается относительно небольшое количество параметров (численность, возраст и половой состав лягушек отдельных форм, а также, при необходимости, динамика их прибытия в модельную популяционную систему). При моделировании изменений программа учитывает изменения в течение каждого «года» (шага работы модели) 16 000 независимых параметров, связанных с отдельными особями в модельной ГПС. Результатом эксперимента является та или иная последовательность трансформаций состава ГПС, а также относительно небольшое количество основных параметров, описывающих достигнутое ей состояние.
Итак, на входе и на выход модели используется небольшое количество параметров, а на каждом шагу анализируется намного большее их число. Это связано с тем, что в ежегодных изменениях ГПС есть как закономерные (детерминированные), так и случайные (стохастические) компоненты. Носитель того или иного генотипа может размножиться и изменить всю популяционную систему, а может и просто погибнуть. Отслеживание «судьбы» каждой отдельной особи и дает возможность моделировать воздействие случайности на трансформации ГПС. Проведя эксперимент с одинаковыми начальными условиями достаточное количество раз, можно определить распределение вероятностей конечного состояния.
Зачем нужны такие эксперименты? Их главное назначение – не определение судьбы конкретных ГПС, а проверка адекватности наших представлений о системах такого рода. Мы узнаем какие-то особенности объектов нашего изучения и строим систему своих представлений о них. Чтобы построить модель, такую систему надо формализовать (что само по себе является весьма полезной работой, выявляющей возможные нарушения логики в общепринятых взглядах). Модель строится на основе системы представлений об объекте и дает возможность сравнить регистрируемое в природе разнообразие объектов с регистрируемым в эксперименте разнообразием состояний модели. Если набор состояний, в которые может переходить ГПС в модели, отличается от набора состояний, наблюдаемых у естественных объектов, то положенные в основу модели представления нуждаются в изменении. Соответствие модели объекту не доказывает правоту системы представлений об объекте, но служит аргументом в ее пользу.
В описанном использовании модели проявляется возможность перехода от редукционистского, поэлементного описания свойств изучаемого объекта к изучению его характеристик, как целого (определению совокупности и распределения вероятностей путей его развития).
С точки зрения авторов, подобный подход может быть применен и при исследовании иных биологических систем различных уровней организации.
Авторы благодарят М.В. Владимирову и Г.Н. Жолткевича за неоценимую помощь в постановке задачи данной работы и ее выполнении.
Кравченко М.А., Луцик А.А., Шабанов Д.А. Что нового можно узнать о зеленых лягушках при помощи имитационного моделирования? (Пленарный доклад на открытии конференции) // «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. - Харьков: СПД ФО Михайлов Г.Г., 2008. - с. 9-11.