ГІБРИДОГЕННИЙ КОМПЛЕКС ЗЕЛЕНИХ ЖАБ ЯК МОДЕЛЬ ДЛЯ ВИВЧЕННЯ БАГАТОРІВНЕВОГО ДОБОРУ
Шабанов Д.А.
Харківський національний університет імені В.Н. Каразіна, кафедра зоології та екології тварин,
е-mail: d.a.shabanov@gmail.com
При спостереженні та моделюванні еволюції гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) виявляються феномени, що для свого пояснення потребують залучення різних підходів. До їх числа належать концепції «егоїстичного гену», «егоїстичного індивіду» та групового добору. Ці концепції не протирічать одна одній, а є доповнюваними (у сенсі принципу доповнюваності Н. Бора). Завдяки цьому названий гібридогенний комплекс є вдалою моделлю для вивчення багаторівневого добору.
Ключові слова: Pelophylax esculentus complex, геміклональне спадкування, геміклональні популяційні системи, «егоїстичний ген», «егоїстичний індивід», груповий добір, багаторівневий добір.
D. A. Shabanov. Hybridogenetic complex of water frogs as a model for multilevel selection studies. Monitoring of hybridogenetic complex of water frogs (Pelophylax esculentus complex) and modeling of its evolution reveal phenomena that may be explained through the concepts of “selfish gene”, “selfish individual” and “group selection”. In fact these concepts are not contradictive and complement each other (in terms of the complementarity principle by Niels Bohr). That is why this hybridogenetic complex is a good model for multilevel selection studies.
Key words: Pelophylax esculentus complex, hemiclonal inheritance, hemiclonal population systems, «selfish gene», «selfish individual», group selection, multilevel selection.
Вступ
Одним із суперечливих питань теоретичної біології, що викликає в останні десятиліття палкі дискусії науковців, є з’ясування рівня, на якому відбувається природний добір. З часів Ч. Дарвіна вважалося, що одиницею добору є організм (4). Творці синтетичної теорії еволюції зробили акцент на вивченні популяцій, як арені змін алельних частот, які пов’язані з виживанням і розмноженням організмів.
Пояснюючи походження альтруїзму, Дарвін припускав, що це – наслідок покращеного виживання груп людей, а не окремих альтруїстів (3). В. К. Вінн-Едвардс назвав таку точку зору груповим добором та намагався пояснити ним широке коло ознак живих організмів (18). Аргументи Вінн-Едвардса були значною мірою розбиті Д. К. Уільямсом (16). Річ у тім, що у ситуації, коли «групові інтереси» вимагають одного способу дій, а індивідуальні - іншого, особини, що реалізують індивідуальні інтереси, будуть підвищувати частку внеску своїх алелей в майбутній генофонд популяції. Наприклад, у випадку, коли популяція існує в умовах гострої нестачі ресурсів, можна припустити, що для її стабільного існування було б вигідно, якби всі особини у її складі зменшили свої витрати на репродукцію. Але та особина, що не зменшить (або підвищить) свою репродукцію, підвищить частку своїх нащадків у майбутніх поколіннях. Природний добір буде сприяти саме таким особинам.
Через певний час маятник хитнувся у протилежний бік. Завдяки Дж. Холдейну, У. Гамільтону, Дж. Мейнарду Сміту та, значною мірою, яскравому популяризатору їх спільних поглядів Р. Докінзу поширення набуває концепція «егоїстичного гену», що розглядає гени як основні одиниці добору (5). З цієї точки зору організми – це лише засоби, завдяки яким гени відтворюють себе самі. Головним аргументом на користь цієї концепції стало уявлення про добір родичів та сукупну пристосованість гена, які були розроблені У. Гамільтоном під час досліджень походження альтруїстичної поведінки у соціальних комах (13).
У наш час спільної точки зору з цього дискусійного питання не знайдено. Значна кількість біологів, особливо, молекулярних, вважають концепцію «егоїстичного гена» остаточно доведеною. Їм протистоять фахівці класичної спеціалізації, які працюють з організмами та вважають цю концепцію редукціоністським спрощенням. Їх погляди можна назвати концепцією «егоїстичного індивіду». Останнім часом суперечки про те, який рівень природного добору є головним, значно посилилися.
Класик еволюційної біології, відомий мірмеколог Е. Уілсон почав відроджувати уявлення про груповий добір. Повною мірою вони відбилися у монографії «Соціальне завоювання Землі», що надрукована у 2012 році (17). Ці уявлення базуються на детальному описі перехідних стадій еволюції соціальності у комах, а також на результатах моделювання процесів еволюції. Такі погляди викликали люту критику з боку Р. Докінза (12). В цілому, можна вважати, що 2012 рік ознаменувався відродженням інтересу до дискусій про головний рівень добору.
Подальший розвиток дослідження охарактеризованої проблеми потребує залучення нових аргументів. На погляд автора, бажано дослідити цю проблему на нових моделях для вивчення, які б виходили за межі традиційно застосованих прикладів (і, бажано, не стосувалися б еволюції альтруїстичної поведінки, як надто складного питання).
У цій роботі обговорено вимоги, які слід висувати до адекватної моделі для дослідження того, який рівень добору єпровідним. Це обговорення дає можливість висловити припущення про те, чи є уявлення про «егоїстичній ген», «егоїстичний індивід» та груповий добір справжніми альтернативами. Насамкінець, у роботі обґрунтовується, чому гібридогенний комплекс зелених жаб є гарною моделлю для вивчення багаторівневого добору.
Основна частина
Альтернативні чи доповнювані концепції? Слід вказати, що сама назва концепції «егоїстичного гену» не є цілком коректною. Згідно з цими поглядами еволюційну перевагу отримують ті варіанти генів (алелей), ефект яких найефективніше приводить до збільшення частки їх копій у наступних поколіннях. Саме тому ці погляди краще називати концепцією «егоїстичного алеля», надалі – ЕА. Аналогічно будемо використовувати скорочення ЕІ для позначення концепції «егоїстичного індивіда», та ЕГ – («егоїстична група») для концепції групового (або ж міждемового (2)) добору.
Як встановити, яка з концепцій (ЕА, ЕІ або ЕГ) є вірною у тому чи іншому випадку? Значна частина суперечок між прихильниками цих поглядів пов’язана з тим, що усі ці концепції добре описують велику кількість процесів. Розглянемо простий приклад.
Ми розглядаємо дві групи, які складаються з особин, що розмножуються і передають нащадкам свої гени. Одна з груп росте швидше, тому що особини у її складі розмножуються (і передають гени) ефективніше, ніж особини іншої групи. Процеси у цьому прикладі однаково успішно описуються концепціями ЕА, ЕІ та ЕГ; спостереження над такими процесами не дають змоги обрати більш адекватне їх пояснення. Але так буває не завжди.
У вступі коротко розглянуто приклад, коли добір на рівні групи сприятиме зниженню плодючості, а добір на рівні особини – зростанню. У таких випадках з’являється можливість встановити, добір якого рівню стає більш ефективним. Спостереження за модельними біосистемами, на які діє протилежно спрямований добір різних рівнів, дає можливість обирати між конкуруючими концепціями. У прикладі, що розглядається, реєстрація зниження плодючості є доказом ЕГ, а її збереження на попередньому рівні – доказом ЕІ або ЕА (з цих двох концепцій випливають однакові прогнози).
Таким чином, для вирішення проблеми, яка обговорюється, слід шукати випадки, коли добір на різних рівнях є протилежно спрямованим.
Так, класичним прикладом дії ЕА можна вважати мейотичний драйв (2) – випадок, коли певний алель підвищує вірогідність свого потрапляння у гамети. На низці прикладів показано, що такі алелі дійсно поширюються у природі, порушуючи розмноження організмів та відтворення популяцій. Таким чином, хоча б у деяких випадках застосування концепції ЕА є необхідним.
Відомим є й приклад, який не можна пояснити з точки зору ЕА. Річ у тому, що у організмів зі статевим розмноженням та мейозом, який відбувається під час гаметогенезу, кожен алель має (у типовому випадку) лише 50 % вірогідність потрапляння до гамет. Цей механізм значно знижує ефективність добору на генному рівні і, незважаючи на це, є широко розповсюдженим у біосфері. Пояснити його можливо лише на рівні ЕГ (гіпотеза вікарія з Брея та інші гіпотези) або ЕІ (гіпотеза Чорної королеви тощо) (10).
Оскільки сучасній науці відомі феномени, що можна пояснити лише з точки зору однієї з перелічених концепцій, можна встановити, що вони не є альтернативними. Ці концепції є доповнюваними (у сенсі принципу доповнюваності Н. Бора), тобто такими, що здаються взаємно заперечуючими, але насправді пояснюючими різні аспекти досліджуваного процесу (1).
Особливості Pelophylax esculentus complex. Гібридогенний комплекс середньоєвропейських зелених жаб, Pelophylax esculentus complex, складається зі ставкових жаб, Pelophylax lessonae (Camerano, 1882), озерних жаб, Pelophylax ridibundus (Pallas, 1771) та їхніх гібридів. Для цих гібридів застосовують назву, яка є подібною до видової, їстівні жаби, Pelophylax esculentus (Linnaeus, 1758) (15).
У типовому випадку під час гаметогенезу P. esculentus у гамети, що вони утворюють, переходить один з геномів (гаплоїдних комплексів хромосом) батьківського виду, або P. lessonae, або P. ridibundus. Розглянемо це на найпростішому прикладі.
У водоймах, що розташовані у м. Харкові, спільно мешкають представники P. ridibundus і P. esculentus. Позначаючи склад геномів у їх генотипах, для геному P. lessonae будемо застосовувати літеру L, а P. ridibundus – R. Можна вказати, що в цьому місцеперебуванні мешкають жаби RR і LR.
Зрозуміло, що представники P. ridibundus продукують гамети з геномом R. Складнішим є випадок P. esculentus.
У Харкові поширені диплоїдні P. esculentus (у деяких інших місцеперебуваннях, наприклад, у Сіверсько-Донецькому центрі різноманіття зелених жаб, масово поширені й триплоїдні (7, 11, 14)). Під час гаметогенезу один з геномів, рекомбінантний геном (у обговорюваному випадку – геном R) таких жаб елімінується (знищується). Інший геном, клональний (у даному разі – геном L) подвоюється внаслідок ендоредуплікації. Після цього утворюються гамети, що несуть ідентичні копії батьківського геному L. Таке успадкування має назву геміклонального (напівклонального) (15).
Для позначення клонального геному його символ будемо брати у дужки. Особини, які отримали від батьків два однакових клональних геноми, не є життєздатними. У такому разі, ми можемо встановити, що в описуваних місцеперебуваннях можливими є три типи схрещувань:
(1) RR × RR → RR
(2) (L)R × (L)R → (L)(L) →†
(3) (L)R × RR → (L)R
Перший та другий тип схрещувань не впливає на співвідношення P. ridibundus і P. esculentus, а третій призводить до збільшення частки P. esculentus у місцеперебуванні. Можна впевнитися, що під час спільного розмноження представники батьківських видів і гібриди зелених жаб утворюють своєрідні системи, де передаються як клональні, так і рекомбинантні геноми – геміклональні популяційні системи, далі – ГПС (11). Для ГПС є характерними трансформації їх складу, що викликані внутрішніми та зовнішніми причинами; ці трансформації можна вивчати як завдяки спостереженню за природними системами, так і з застосуванням імітаційного моделювання (8, 9).
Рівні добору в P. esculentus complex. З урахуванням викладеного, можна визначити, на яких рівнях діє добір, що обумовлює особливості трансформацій ГПС P. esculentus complex.
В розглянутому прикладі популяційних систем у м. Харкові значних трансформацій ГПС не відбувається. Скоріше за все, це пов’язано з тим, що в умовах великого міста представники P. ridibundus мають вищу життєздатність, ніж P. esculentus. Але так буває не завжди. Те, що відбувається у тому разі, якщо життєздатність представників батьківського виду та гібридних жаб є однаковою, можна встановити із застосуванням імітаційного моделювання (9). В прикладі, що показаний на рис. 1, у позначеннях генотипів вказана також статева приналежність геномів. Слід вказати, що система визначення статі у зелених жаб подібна до такої у людей: чоловіча стать є гетерогаметною, жіноча – гомогаметною.
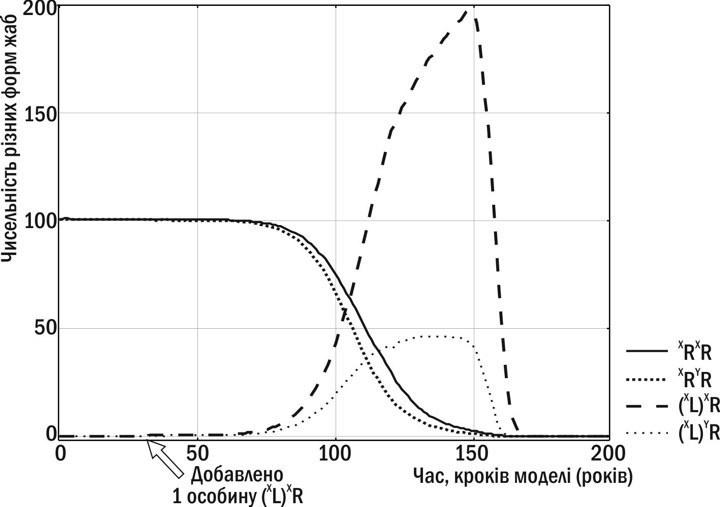
Рис. 1. Трансформації ГПС P. esculentus complex за даними імітаційного моделювання (9). Потрапляння в популяцію P. ridibundus особини P. esculentus з жіночим клональним геномом – (XL)XR – призводить до того, що всі особини в ГПС несуть клональний геном, відтворення ГПС припиняється та ГПС гине
Геміклональне спадкування геному (у випадку, що показаний на рис.1 – геному XL) можна розглядати як граничний випадок мейотичного драйву, при якому у гамети переходить не окремий алель, а цілий геном. Той стан, у який потрапила показана на рис. 1 ГПС, є шкідливим «з точки зору» як ЕГ, так і ЕІ. Можливість саме такого перебігу обставин не тільки витікає з результатів моделювань, але й певною мірою підтримується результатами спостережень.
Але передача клональних геномів не завжди призводить до порушень відтворення ГПС. На рис. 2 показано, що відбудеться, якщо в ГПС будуть передаватися клональні геноми, що належать до різних батьківських видів. ГПС у цьому разі може увійти в стабільний стан.
Ми маємо підстави припустити, що ті ГПС, що перебувають у нестабільному стані, змінюються або зникають, а ті, в яких утворився стабільний набір геномів, розселяються. У такому разі ці процеси є прикладом групового добору (до речі, не пов’язаним з альтруїзмом). Найважливіший аргумент Р. Докінза (6) проти групового добору полягає в тому, що група позбавлена генотипу, і не може передавати свої властивості у спадок. В розглянутому випадку цей аргумент не працює. Аналогом генотипу особини на рівні групи є її генофонд. Важливіша характеристика генофонду ГПС – набір клональних геномів, які в ній передаються. «Розмноження» груп відбувається в ході дисперсії молоді, при якій дочірня група може одержувати відносно репрезентативну вибірку клональних геномів з генофонду материнської групи.
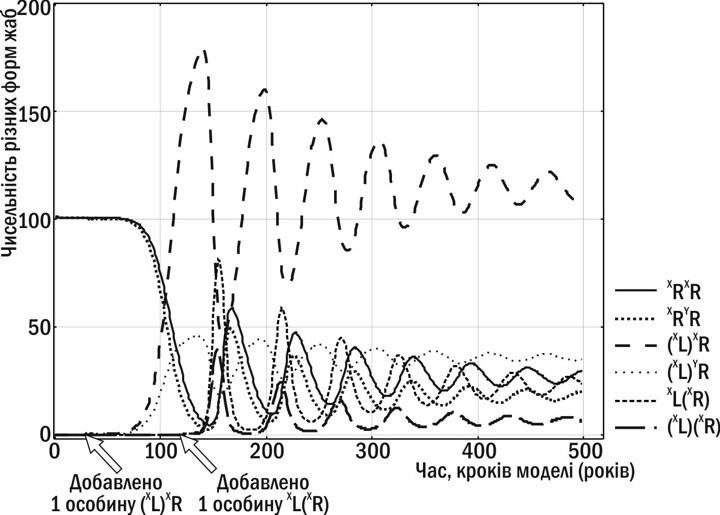
Рис. 2. Трансформації ГПС P. esculentus complex за даними імітаційного моделювання (9). ГПС, в якій передаються різні клональні геноми, переходить до стабільного стану та нормально відтворюється
Ефективність групового добору зі порівнянні з генним і індивідуальним знижена, оскільки передача репрезентативної вибірки з генофонду – стохастичний, менш стійкий процес, ніж передача генів при розмноженні. Але у такого добору є й сильні сторони: наприклад, в ньому можуть успадковуватися «придбані» ознаки (підтримувані добором клональні геноми, що потрапили у ГПС або виникли внаслідок еволюції раніше існуючих геномів).
З викладеного стає ясним, що трансформації ГПС P. esculentus complex є складним процесом, на якій може впливати добір на різних рівнях. У певних випадках (далеко не повністю описаних у цій роботі) добір на різних рівнях може виявлятися протилежно спрямованим. Це робить гібридогенний комплекс зелених жаб чудовою моделлю для вивчення багаторівневого добору.
Висновки
1. Концепції «егоїстичного гена», «егоїстичного індивіда» та групового добору не є альтернативами. Вони взаємно доповнюють одна одну (у сенсі Н. Бора).
2. Для окремого вивчення добору на певному рівні необхідні моделі, де добір на різних рівнях є протилежно спрямованим. Наприклад, якщо добір алелей є протилежним добору індивідів та груп, і спостереження доводять, що зміни йдуть у напрямку, що задається добором алелей, така ситуація є оптимальною для вивчення добору алелей.
3. Геміклональне спадкування геномів у Pelophylax esculentus може бути прикладом дії «егоїстичного гена», а трансформації нестійких геміклональних популяційних систем зелених жаб – прикладом групового добору. В цілому гібридогенний комплекс зелених жаб дає широкі можливості для вивчення багаторівневого добору.
Подяки
Викладені в цій статті міркування є наслідком спільної роботи автора по вивченню зелених жаб з багатьма колегами. Особливо важливим для автора було співробітництво з М. О. Кравченко, О. В. Коршуновим, М. В. Владимировой, Г. А. Ладою, С. М. Литвинчуком, Л. Я. Боркіним і С. Ю. Морозовим-Леоновим. Автор висловлює усім названим та неназваним колегам щиру подяку.
Література
1. Бор Н. Атомная физика и человеческое познание / Нильс Бор. – М: Иностр. лит., 1961. – 152 с.
2. Грант В. Эволюционный процесс. Критический обзор эволюционной теории / Верне Грант. – М.: Мир, 1991. – 488 с.
3. Дарвин Ч. Происхождение человека и половой отбор. Соч. / Чарльз Дарвін – М.: Изд-во АН СССР, 1953. – Т. 5. – 1040 с.
4. Дарвин Ч. Происхождение видов путем естественного отбора или Сохранение благоприятных рас в борьбе за жизнь, Перевод с шестого издания (Лондон, 1872) / Чарльз Дарвін – СПб.: "Наука", 1991. – 539 с.
5. Докинз Р. Эгоистичный ген / Ричард Докинз. – М.: «Мир», 1993. – 317 с.
6. Докинз Р. Расширенный фенотип: длинная рука гена / Ричард Докинз. – М.: Астрель: CORPUS, 2010. – 510 с.
7. Изучение популяционных систем зеленых лягушек (Rana esculenta complex) в Харьковской области: история, современное состояние и перспективы / Д. А. Шабанов, А. И. Зиненко, А. В. Коршунов (и др.) // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2006. – Вип. 3 (№ 729). – С. 208–220.
8. Исследование устойчивости гемиклональных популяционных систем гибридогенного комплекса зеленых лягушек при помощи имитационного моделирования / М. А. Кравченко, Д. А. Шабанов, М. В. Владимирова (и др.) // Вісник Дніпропетровського університету. Біологія. Екологія. – 2011. – Вип. 19, т. 1. – С. 51–64.
9. Кравченко М. А. Моделирование трансформаций гемиклональных популяционных систем зеленых лягушек (Pelophylax esculentus complex; Amphibia, Ranidae) с помощью рекуррентных разностных уравнений / М.А. Кравченко, Д.А. Шабанов // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2010. – Вип.12 (№ 920). – С. 70–82.
10. Ридли М. Секс и эволюция человеческой природы / Метт Ридли. – М: Эксмо, 2011. – 448 с.
11. Шабанов Д. А.Зеленые лягушки: жизнь без правил или особый способ эволюции? / Д. А. Шабанов, С. Н. Литвинчук // Природа. – 2010. – № 3 (1135). – С. 29–36.
12. Dawkins R. The descent of Edward Wilson. Prospect, 24 May 2012 (Online).
13. Hamilton W. D. The genetical evolution of social behavior (I and II) / Hamilton W. D. // Journal of 'Theoretical Biology. – 1964. – V. 7 – P. 1–16; 17–52.
14. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine / L. J. Borkin, A. V. Korshunov, G. A. Lada (et al.) // Russian Journal of Herpetology. – 2004. – Vol. 11, No 3. – P. 194–213.
15. Plotner J. Die westpalaarktichen Wasserfrosche / Jorg Plotner. – Bielefeld: Laurenti-Verlag, 2005. – 161 s.
16. Williams G. C. Adaptation and natural selection: a critique of some currentevolutionary thought / George Christopher Williams. – Princeton, 1966. – 307 p.
17. Wilson E. The Social Conquest of Earth / Edward Wilson. – N.Y.: Liveright Publishing Corporation, 2012. – 352 p.
18. Wynne-Edwards V.C. Animal Dispersion in Relation to Social Behaviour / Vero Corner Wynne-Edwards. – Edinburgh: Oliver & Boyd, 1962. – 653 p.
Стаття поступила до редакції 15.10.2012 р.; прийнята до друку 22.10.2012 р.