Українська мова (найновіша версія) / Русский язык (обновление прекращено)
3.13. Экологические пирамиды
Чарльз Эльтон предложил способ графического выражения отношений между трофическими уровнями, который стал едва ли не символом экологии как науки. Речь идет об экологических пирамидах. При построении экологических пирамид меры обилия представителей разных трофических уровней показывают в виде лежащих друг на друге прямоугольников. Обычно этот метод используют для описания пастбищных трофических цепей. Выделяют пирамиды численностей, биомасс и продуктивностей.
Построим несколько экологических пирамид и мы. Наши примеры будут достаточно условными: мы предположим, что трофические цепи, которые мы будем описывать, не имеют «разветвлений». Например, моделируя цепь клевер — овцы — волки, мы примем, что овцы питаются только клевером, а волки — только овцами, при этом нас будут интересовать отношения между этими трофическими уровнями в пределах какой-то экосистемы, где соотношение численностей рассматриваемых уровней достигло равновесия. Естественно, экологические пирамиды можно использовать и для описания естественных экосистем, а принятые нами упрощения нужны лишь для большей наглядности наших рассуждений.
Возвратимся к цепи клевер — овцы — волки. Оценив численности популяций в этой цепи, мы увидим, что отдельных растений клевера намного больше, чем овец, а овец — больше, чем волков. Такая пирамида (с основанием шире вершины) называют правильной или прямой. Однако не для всех пастбищных цепей пирамиды численностей будут правильными. Примером может быть цепь дубы — дубовые шелкопряды. На небольшом количестве крупных деревьев может обитать множество гусениц. Пирамида численностей будет для этой цепи перевернутой (рис. 3.13.1).
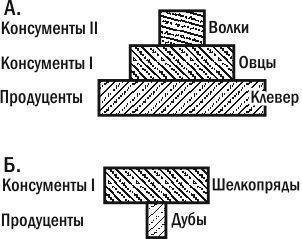
Рис. 3.13.1. Пирамиды численностей. А. прямая. Б. Перевернутая. Перевернутый характер второй пирамиды связан с отличиями особей по размерам
Несложно понять, что перевернутость второй пирамиды связана с отличиями особей продуцентов и консументов по их размерам. На основании данных о том, сколько весит средний дуб и средний шелкопряд, а также пирамиды их численностей, мы можем построить пирамиду биомасс. Естественно, она будет прямой (рис. 3.13.2).
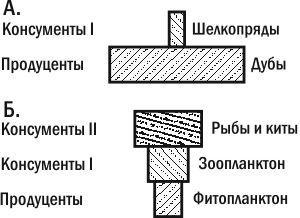
Рис. 3.13.2. Пирамиды биомасс. А. прямая. Б. Перевернутая. Перевернутый характер второй пирамиды связан с различиями особей в «скорости жизни» — разной интенсивности протекания энергии через их биомассу
Могут ли пирамиды биомасс быть перевернутыми? Достаточно редко, но могут. Рассмотрим пастбищные трофические цепи толщи воды в океане. Как ни удивительно, биомасса продуцентов (планктонных водорослей) в таких цепях зачастую оказывается меньше биомассы консументов. Означает ли это, что консументы в таких цепях существуют не за счет продуцентов? Нет.
Понять сказанное поможет простая аналогия. Большой пруд может существовать благодаря впадающему в него маленькому ручейку, хотя в любой момент времени масса воды в пруду намного больше, чем в ручье. Понятно, что это становится возможным потому, что вода в ручье сменяется намного быстрее, чем в пруду. Так и в сообществах толщи воды энергия течет через разные трофические уровни с разной скоростью. Время смены биомассы фитопланктона измеряется часами, зоопланктона — днями, рыб и китов — неделями и месяцами. Чтобы учесть это различие, нам надо отразить в экологических пирамидах интенсивность потока энергии через каждый уровень. На основании данных о биомассе звеньев трофической цепи и о скорости ее смены мы можем построить пирамиду продуктивностей (или потоков энергии; рис. 3.13.3).
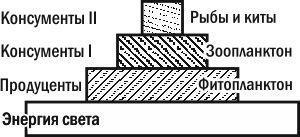
Рис. 3.13.3. Пирамиды продуктивности всегда являются прямыми
Такая пирамида всегда будет прямой. Первое начало термодинамики (закон сохранения энергии) «запрещает» такой пирамиде быть перевернутой, а второе — иметь «этажи» одинаковой ширины, ведь при каждом преобразовании энергии часть ее должна рассеиваться в виде тепла. Кстати, именно поэтому реальные трофические цепи не бывают очень длинными, а экологические пирамиды — высокими. В любой действительной экосистеме до консумента X уровня дошло бы столь мало энергии (после десяти последовательных преобразований!), что с территории, доступной для одной особи, не удалось бы собрать необходимое для нее количество энергии.
Теперь, познакомившись с логикой, по которой строятся экологические пирамиды, рассмотрим два более конкретных примера. Юджин Одум рассчитал параметры условной пищевой цепи, в которой двенадцатилетний мальчик питался исключительно телятиной (учтите: есть только мясо противоестественно!), а телята — только люцерной (это уже более физиологично, не считая того, что и мальчику, и телятам надо начинать жизнь, питаясь молоком своих матерей). Характеристики такой пирамиды приведены в табл. 3.13.1.
Таблица 3.13.1. Пример экологических пирамид для условной трофической цепи
|
|
Численность |
Биомасса |
Продуктивность |
|
Мальчик |
1 |
48 кг |
8,3×103 |
|
Телята |
4,5 |
1 035 кг |
1,2×106 |
|
Люцерна |
2×107 |
8 211 кг |
1,5×107 |
|
Использованное излучение |
— |
— |
6,3×109 |
А следующий пример (рис. 3.13.4) касается реальных данных о биомассе нескольких видов млекопитающих в североамериканском листопадном лесу. Как вы можете увидеть, наивысшую биомассу имеют растительноядные млекопитающие, а наименьшую — плотоядные, что и следовало ожидать, исходя из изложенных здесь соображений.
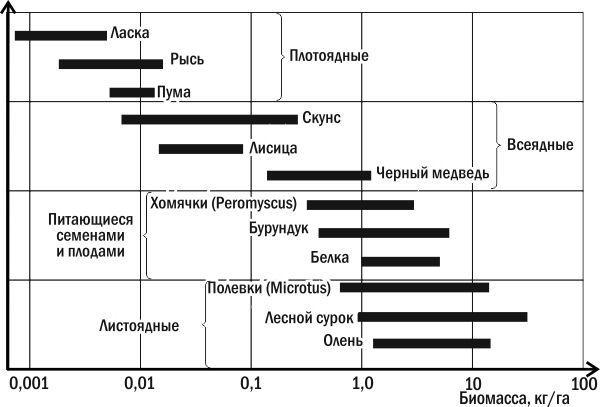
Рис. 3.13.4. Биомасса нескольких видов млекопитающих североамериканского листопадного леса
Как вы поняли, экологические пирамиды не могут быть очень высокими, потому что при переходе с уровня на уровень часть энергии теряется. Однако разные организмы теряют разное количество энергии. В разных сообществах средний уровень экологической эффективности отличается и тесно связан с количеством трофических уровней, как это показано в табл. 3.13.2.
Таблица 3.13.2. Среднее число трофических уровней в разных биомах (Р. Риклефс, 1977)
|
Биом |
Средняя экологическая эффективность (отношение продуктивности хищника к продуктивности жертвы) |
Среднее количество трофических уровней |
|
Открытый океан |
25 % |
7,1 |
|
Морское прибрежье |
20 % |
5,1 |
|
Степь |
10 % |
4,3 |
|
Тропический лес |
5 % |
3,2 |
Конечно, экологическая эффективность на разных трофических уровнях сильно отличается, и особенно она низка у основания экологических пирамид. Питание растительной пищей — более сложная биохимическая и физиологическая «задача», чем питание животной пищей. В большинстве наземных экосистем регистрируется избыток растительной пищи. Зато количество растительноядных животных (консументов I уровня) обычно хорошо контролируется плотоядными животными. Избыток организмов на этом уровне будет эффективно «выедаться» организмами следующего уровня.
Согласно гипотезе естественного равновесия, главным регулятором соотношения трофических уровней является вершинный хищник — консумент самого высокого уровня. Значит, в системе с четным числом трофических уровней эффективнее контролируются консументами нечетные уровни (1-й, 3-й, 5-й…), а с нечетным числом уровней — четные. Поскольку в наземных экосистемах труднее всего контролировать именно первый уровень, уровень продуцентов, можно ожидать, что в наземных системах чаще должно наблюдаться нечетное число трофических уровней. Наблюдения подтверждают это предположение.
Коментарі
Спасибо,
Спасибо, интересно.