УДК: 567.828: 612.663
https://orcid.org/0000-0003-3247-6882
ПОРУШЕННЯ ФЕРТИЛЬНОСТІ У МІЖВИДОВИХ ГІБРИДІВ ЗЕЛЕНИХ ЖАБ ІЗ СІВЕРСЬКО-ДОНЕЦЬКОГО ЦЕНТРУ РІЗНОМАНІТТЯ PELOPHYLAX ESCULENTUS COMPLEX
Боброва А.А., Макарян Р.М., Шейко В.П., Шабанов Д.А.
Харківський національний університет імені В.Н. Каразіна
Досліджено 46 диплоїдних самців P. esculentus та 14 самців P. ridibundus з Сіверсько-Донецького центра різноманіття Pelophylax esculentus complex. Визначено здатність продукувати активні сперматозоїди у відповідь на гормональну стимуляцію сурфагоном, наявність активних сперматозоїдів у суспензії з розрізаних сім’яників, а також розміри сім’яників. 85% P. esculentus та 29% P. ridibundus серед досліджених особин мають порушення фертильності. Особини P. esculentus із порушеннями фертильності мають сім’яники, що є меншими за розміром у порівнянні з нормальними особинами.
Ключові слова: Pelophylax esculentus complex, Pelophylax ridibundus, Сіверсько-Донецький центр різноманіття зелених жаб, сім’яники, фертильність.
Impaired fertility in interspecific hybrids of green frogs from Seversko-Donetskiy center of Pelophylax esculentus complex diversity. Bobrova A. A., Makaryan R. M., Sheiko V. P., Shabanov D. A. – 46 diploid males P. esculentus and 14 males P. ridibundus from Seversko-Donetskiy center of green frogs diversity were investigated. An ability to produce active sperm in response to hormonal stimulation by surfagon, the presence of active sperm in homogenized testes suspension, as well as the size of the testes were determined. Among studied individuals, 85% of P. esculentus specimens and 29% of P. ridibundus specimens had an impaired fertility. The size of testes of P. esculentus individuals with impaired fertility was smaller than normal.
Key words: Pelophylax esculentus complex, Pelophylax ridibundus, Seversko-Donetskiy center of green frogs diversity, fertility.
ВСТУП
У типовому випадку відтворення диплоїдних гібридів стає можливим завдяки тому, що в клітинах зародкової лінії відбувається елімінація одного з батьківських геномів та ендоредуплікація (подвоєння без поділу клітини) іншого. Такі гібриди продукують статеві клітини, що містять один з батьківських геномів, який передається без рекомбінації, клонально. У разі схрещування таких гібридів з представниками іншого батьківського виду усе потомство складається з гібридних особин. Слід підкреслити, що описана схема є лише одним з випадків (хоча й типовим за сучасними уявленнями). У багатьох регіонах поруч з диплоїдними гібридами поширені й триплоїдні й навіть трапляються тетраплоїдні; деякі диплоїдні гібриди продукують диплоїдні гамети, гамети з частково рекомбі-нованими геномами або навіть виробляють суміш статевих клітин, що належать до обох батьківських видів (6; 8).Гібридогенний комплекс зелених жаб, Pelophylax esculentus complex (або, за старою назвою – Rana esculenta complex) став моделлю для вивчення геміклональної (напівклональної) міжвидової гібридизації (8). Внаслідок схрещування двох батьківських видів, ставкової жаби, Pelophylax lessonae (Camerano, 1882) та озерної жаби, Pelophylax ridibundus (Pallas, 1771) виникають міжвидові гібриди, для позначення яких використовується ім’я, що аналогічне видовому, Pelophylax esculentus (Linnaeus, 1758).
Розташований у Східній Україні регіон, де водночас зустрічається висока кількість різних форм зелених жаб, отримав назву Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex (5; 6). У ньому відсутні статевозрілі представники P. lessonae. Внаслідок цього усі геноми P. lessonae, що входять у генотипи диплоїдних та триплоїдних P. esculentus, мають довгу історію передачі через низку гібридних особин. Іншою особливістю цього центру є те, що певна частка особин P. esculentus у ньому є триплоїдними.
Відомо, що гаметогенез у гібридів F1, тобто особин, що утворилися від схрещування P. lessonae і P. ridibundus, може стикатися зі значними склад-нощами. Вперше це виявив Лешек Бергер, який відкрив гібридогенез у зелених жаб (7). Бергер встановив, що серед гібридів F1 зустрічаються особини, у сім’яниках яких сперматозоїди або відсутні, або знаходяться у кількості, набагато нижчій за норму.
Вивчення особливостей сперматогенезу амфібій має не лише теоретичне значення. В останні роки розробляються технології, що дозволяють прижиттєве отримання сперми самців безхвостих амфібій. Цю сперму можна застосувати як для запліднення ікри, так і для тривалого зберігання у замороженому стані. Такі можливості є корисними для збереження генофонду рідкісних та зникаючих видів та внутрішньовидових форм, а також для розширення можливостей дослідників під час проведення штучних схрещувань. Піонером таких досліджень є В. К. Утєшев, за участю якого розроблено технології прижиттєвого отримання сперми від одного з батьківських видів гібридогенного комплексу зелених жаб, P. lessonae (9 та попередні роботи). Після правильно обраної гормональної стимуляції з клоаки самців безхвостих можна отримати уринальну сперму, що містить достатню для штучного запліднення ікри кількість сперматозоїдів.
Необхідність у визначенні частки самців, що є повністю або майже повністю безплідними, стала наслідком нашої роботи по адаптуванню технології прижиттєвого отримання сперми до самців P. esculentus та P. ridibundus (детальні результати цієї роботи будуть оприлюднені в іншій публікації). Під час виконання цієї задачі ми стикнулися з тим, що гормональна стимуляція, яка викликала появу активних сперматозоїдів в уринальній спермі одних особин, не приводила до бажаного ефекту у інших.
МАТЕРІАЛИ ТА МЕТОДИ ДОСЛІДЖЕННЯ
Для вивчення гормональної стимуляції застосували 43 диплоїдних самця P. esculentus та 8 самців P. ridibundus. З числа особин, для яких ми встановили реакцію на гормональну стимуляцію, 11 P. esculentus та 2 P. ridibundus були розтяті для визначення розмірів сім’яників та наявності в них активних сперматозоїдів. Інші особини, яких використовували для досліджень гормональної стимуляції, були випущені у місцях збору. Крім того, було розтято 3 P. esculentus та 6 P. ridibundus, випадково зібраних у природному місцеперебуванні. Таким чином, ми дослідили здатність продукувати активні сперматозоїди у 60 особин. Відносно невелика кількість жаб, які були розтяті, пояснюється тим, що ми намагалися мінімізувати вилучення жаб із природних місцеперебувань і обмежились кількістю особин, яка дала нам можливість порівняти два способи визначення наявності активних сперматозоїдів.
Жаб збирали у червні-липні 2014 р. в околицях біологічної станції Харківського національного університету імені В. Н. Каразіна (с. Гайдари Зміївського району Харківської області), засліплюючи їх ліхтарем у темряві.
Визначення жаб. У Сіверсько-Донецькому центрі різноманіття зелених жаб мешкають диплоїдні P. esculentus та P. ridibundus, які добре відрізняються за зовнішніми ознаками, а також триплоїдні P. esculentus, зовнішнє визначення яких є вкрай ненадійним (1; 5). Тому визначення жаб проводилося таким чином: їх плоїдність визначали за розміром еритроцитів із застосуванням раніше оприлюдненої методики (2). Оскільки усі досліджені особини були диплоїдними, їх належність до P. esculentus або P. ridibundus визначали за комплексом зовнішніх ознак (1).
Методика гормональної стимуляції. Статевозрілим самцям зелених жаб ін’єктували сурфагон, синтетичний аналог люліберину, гонадотропного рілізинг-гормону (9). Під час ін’єкції голка шприцу вводилася крізь шкіру у стегновий підшкірний лімфатичний мішок, а з нього проводилася до черевного підшкірного лімфатичного мішку. Ми застосовували різні дози гормону, найчастіше – 1 мкг сурфагону на 1 г живої ваги жаби.
Через 2 години після ін’єкції гормону шляхом м’якого масажу черевної ділянки отримували уринальну сперму (9). Рідину, що при масажі витікає із клоаки, збирали в чашки Петрі. Проби цієї рідини досліджували під мікроскопом у камері Горяєва, в якій проводили підрахунок кількості активних сперматозоїдів. Активними вважали сперматозоїди, що рухаються та мають нормальну для жаб форму.
У тому разі, якщо у певної особини реєструвалась реакція на гормональну стимуляцію, різниця у реакції на різні дози сурфагону стосувалася лише кількості активних сперматозоїдів в уринальній спермі, а не їх наявності або відсутності. Особини, у яких сперматозоїди не з’являлися після дози в 1 мкг гормону на 1 г живої ваги жаби, не реагували і на інші, у тому числі підвищені, дози гормону.
Методика розтину та визначення наявності активних сперматозоїдів. Жаб наркотизували парою ефіру для наркозу, після чого руйнували їм головний та спинний мозок введенням препарувальної голки через потилицевий отвір. У вбитих тварин розкривали черевну порожнину та виймали сім’яники. Жабу (спинною стороною донизу) та її відокремлені сім’яники розміщували на склі офісного сканеру формату А4, після чого проводили сканування з роздільною якістю 600 dpi. Виміри довжини тіла жаби та довжини сім’яників проводили по фотографіях за допомогою програми PDF XChange VIEWER. Результати вимірів перераховували у міліметри, застосовуючи скановане зображення штангенциркулю як мірну лінійку. Дані заносили до бази даних, створеної у програмі Statistica, 8 (StatSoft Inc.); у цій базі обчислювали відносну усереднену довжину сім’яників та їх асиметричність (відношення різниці у довжині правого та лівого сім’яника до їх усередненої довжини), а також проводили статистичну обробку даних.
Для визначення наявності активних сперматозоїдів у сім’яниках розрізали один з сім’яників ножицями на кілька шматків, які поміщали у чисту річкову воду. Через кілька хвилин, що необхідні для активації сперматозоїдів (3), пробу отриманої суспензії мікроскопували та реєстрували наявність або відсутність активних сперматозоїдів.
РЕЗУЛЬТАТИ ДОСЛІДЖЕНЬ ТА ЇХ ОБГОВОРЕННЯ
Співвідношення самців зелених жаб, у яких реєструвалася наявність активних сперматозоїдів або їх відсутність, показано в таблиці 1. Слід підкреслити, що незалежними дослідженнями є перше (визначення наявності активних сперматозоїдів в уринальній спермі після гормональної стимуляції) та третє (визначення наявності активних сперматозоїдів у суспензії, отриманій з сім’яників особин, що були випадково обрані з природного місцеперебування). Друге дослідження, результати якого показані в таблиці 1, є підтвердженням відповідності двох застосованих підходів.
Зареєстровані порушення фертильності P. esculentus пов’язані з недорозвиненістю їх сім’яників. На рис. 1 показані результати двохфакторного дисперсійного аналізу, в якому встановлювався зв’язок між, з одного боку, належністю самця до P. esculentus або P. ridibundus та наявністю або відсутністю у нього активних сперматозоїдів, і, з іншого боку, відносним розміром сім’яників. Різниця у здатності продукувати активні сперматозоїди між особинами P. esculentus з різним розміром статевих залоз, а також між P. esculentus і P. ridibundus є значущою.
Таблиця 1
Частка особин, у яких реєструвалася наявність або відсутність активних сперматозоїдів за результатами різних досліджень
|
№ |
Тип дослідження |
Міжвидові гібриди, P. esculentus |
Представники батьківського виду, P. ridibundus |
|
1. |
Наявність активних сперматозоїдів в уринальній спермі, отриманій після гормональної стимуляції |
наявні: 7 (16%) відсутні: 36 (84%) разом: 43 |
наявні: 6 (75%) відсутні: 2 (25%) разом: 8 |
|
2. |
Наявність активних сперматозоїдів у суспензії, отриманої з сім’яників особин, для яких було встановлено їх реакцію на гормональну стимуляцію |
наявні: 4 відсутні: 7 разом: 11 (відповідність методів —100%) |
наявні: 2 відсутні: 0 разом: 2 (відповідність методів —100%) |
|
3. |
Наявність активних сперматозоїдів у суспензії, отриманої з сім’яників особин, що були випадково обрані з природних місцеперебувань |
наявні: 0 відсутні: 3 разом: 3 |
наявні: 4 відсутні: 2 разом: 6 |
|
4. |
Разом для досліджень 1 і 3 |
наявні: 7 (15%) відсутні: 39 (85%) разом: 46 |
наявні: 10 (71%) відсутні: 4 (29%) разом: 14 |

Рис. 1. Зв’язок здатності самців жаб продукувати активні сперматозоїди з відносним розміром сім’яників, та з належністю до батьківського виду або до міжвидових гібридів
На рис. 2 показаний розмір сім’яників у тих самців, що виробляли або не виробляли активну сперму. Можна побачити, що серед P. esculentus активні сперматозоїди продукували ті особини, що мали більші за розміром статеві залози. Серед P. ridibundus порушення фертильності спостерігали у особин, які мали найбільші та найменші сім’яники.

Рис. 2. Залежність здатності продукувати активні сперматозоїди самців жаб, що належать до батьківського виду або до міжвидових гібридів, від відносного розміру сім’яників
Значного зв’язку між наявністю активних сперматозоїдів та асиметрією сім’яників не зареєстровано (рис. 3).
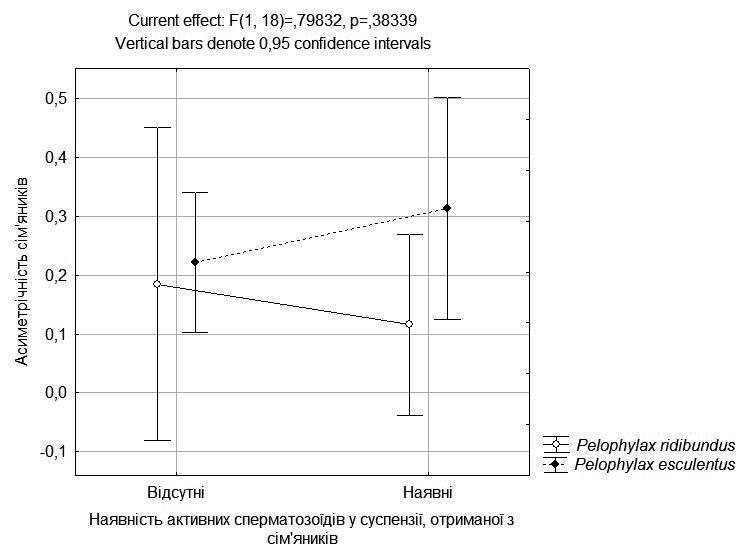
Рис. 3. Зв’язок здатності самців жаб продукувати активні сперматозоїди з асиметричністю сім’яників, а також з належністю до батьківського виду або до міжвидових гібридів
Вкрай низька частка самців P. esculentus, що продукують активні сперматозоїди (15%), викликає подив. Слід підкреслити, що ми не вважаємо, що особини, у яких не зареєстровані активні сперматозоїди, є повністю безплідними. Втім, є усі підстави вважати, що фертильність (тобто здатність до розмноження) таких особин є або значною мірою порушеною, або повністю втраченою.
Отриманий нами результат є певною мірою парадоксальним. Досліджені жаби були зібрані у районі, де приблизно половина особин належить до P. esculentus. Якщо відтворення P. esculentus стикається зі значними складнощами, лишається незрозумілим, яким чином вони відтворюються. Як ми зазначили, у регіоні дослідження відсутні представники одного з батьківських видів, саме тому ми можемо бути впевнені, що досліджували не гібридів F1, а особин, які мають геноми P. lessonae, що передавалися через гібридних особин протягом багатьох поколінь.
Несподіваним результатом також стала і реєстрація досить великої частки (29%) особин з порушеннями фертильності серед представників батьківського виду.
Вірогідно, порушення фертильності P. esculentus, зареєстровані ще Бергером (7), – наслідок того, що узгоджена елімінація усіх 13 хромосом одного набору є складним процесом. Його безпомилковість має забезпечуватися досить довершеним цитогенетичним механізмом. Можна припустити, що цей механізм у його повному вигляді не може бути наслідком добору, що йшов у батьківських видів, тому що в їх представників не відбувається елімінація одного з геномів. Ймовірно, стабілізація гаметогенезу з плином послідовних поколінь геміклональних гібридів може бути наслідком добору на рівні клітинних ліній під час гаметогенезу (4). Нащадків можуть залишити тільки ті гібриди, що сформують повноцінні статеві клітини; продовження отримають лише ті клітинні лінії, в яких відбудеться елімінація одного геному та ендоредуплікація іншого. Але, як показало наше дослідження, у Сіверсько-Донецькому центрі різноманіття зелених жаб гаметогенез гібридних самців лишається значною мірою дестабілізованим.
Автори сподіваються, що відповіді на задані питання дадуть подальші дослідження.
ВИСНОВКИ
Більшість диплоїдних самців P. esculentus (85% з дослідженої вибірки) та значна частина P. ridibundus (29% вибірки) з Сіверсько-Донецького центра різноманіття зелених жаб мають порушення фертильності, що проявляються у відсутності достатньої кількості активних сперматозоїдів в уринальній спермі, яку збирають після гормональної стимуляції за методом В. К. Утєшева, а також у суспензії, отриманій із розрізаних сім’яників. Здатність до продукування активних сперматозоїдів пов’язана з розміром сім’яників; особини P. esculentus із порушеннями фертильності мають сім’яники, що є меншими за розміром за нормальні.
Подяки. Автори щиро вдячні В. К. Утєшеву (Інститут теоретичної та експериментальної біофізики РАН, г. Пущіно, Російська Федерація), який виступив ініціатором розробки методів отримання уринальної сперми у представників Pelophylax esculentus complex Сіверсько-Донецького центру різноманіття зелених жаб та надав цінні консультації з цього приводу.
Література
- Атемасова Т. А., Влащенко А. С., Зиненко А. И., Токарский В. А., Шабанов Д. А., Шандиков Г. А. Учебно-полевая практика по зоологии позвоночных / Т. А. Атемасова, А. С. Влащенко, А. И. Зиненко, В. А. Токарский, Д. А. Шабанов, Г. А. Шандиков. – Х. : ХНУ имени В. Н. Каразина, 2008. – 180 с.
- Бондарева А. А., Бибик Ю. С., Самило С. М., Шабанов Д. А. Цитогенетические особенности эритроцитов зеленых лягушек из Северско-Донецкого центра разнообразия Pelophylax esculentus complex / А. А. Бондарева, Ю. С. Бибик, С. М. Самило, Д. А. Шабанов // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2012. – Вип.15 (№1008) – С. 116–123.
- Дабагян Н. В., Слепцова Л. А. Травяная лягушка Rana temporaria L. / Н. В. Дабагян, Л. А. Слепцова // Объекты биологии развития. – М.: Наука, 1975. – С. 442-462.
- Шабанов Д. А. Гібридогенний комплекс зелених жаб як модель для вивчення багаторівневого добору / Д. А. Шабанов // Вісник Прикарпатського національного університету імені Василя Стефаника. Серія – Біологія. – Івано-Франківськ: Вид-во Прикарпатського національного університету імені Василя Стефаника, 2012. – Вип. XVII. – С. 90-94.
- Шабанов Д. А., Коршунов О. В., Кравченко М. О. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми / Д. А. Шабанов, О. В. Коршунов, М. О. Кравченко // Біологія та валеологія. – Вип. 11. – Харків: ХДПУ, 2009. – С. 164-125.
- Шабанов Д. А., Литвинчук С. Н. Зеленые лягушки: жизнь без правил или особый способ эволюции? / Д. А. Шабанов, С. Н. Литвинчук // Природа. – 2010. – № 3 (1135). – С. 29-36.
- Berger L. Viability, sex and morphology of F2 generation within forms of Rana esculenta complex. Zool. Poloniae, 1971. – V. 21. – P. 345-393.
- Plötner J. Die Westpaläarktichen Wasserfrösche / J. Plötner. – Bielefeld, 2005. – 161 s.
- Uteshev V., Shishova N., Kaurova S., Manokhin А., Gakhova E. Collection and Cryopreservation of Hormonally Induced Sperm of Pool Frog (Pelophylax lessonae) / V. Uteshev, N. Shishova, S. Kaurova, А. Manokhin, E. Gakhova // Russian Journal of Herpetology, 2013. – Vol 20, No 2. – P. 105-109.
Нарушения фертильности у межвидовых гибридов зеленых лягушек из Северско-Донецкого центра разнообразия Pelophylax esculentus complex. Боброва А. А., Макарян Р. М., Шейко В. П., Шабанов Д. А. – Исследовано 46 диплоидных самцов P. esculentus и 14 самцов P. ridibundus из Северско-Донецкого центра разнообразия Pelophylax esculentus complex. Определены способность производить активные сперматозоиды в ответ на гормональную стимуляцию сурфагоном, наличие активных сперматозоидов в суспензии из разрезанных семенников, а также размеры семенников. 85% P. esculentus и 29% P. ridibundus среди исследованных особей имеют нарушения фертильности. Особи P. esculentus с нарушениями фертильности имеют семенники, меньшие по размеру, в сравнении с нормальными особями.
Ключевые слова: Pelophylax esculentus complex, Pelophylax ridibundus, Северско-Донецкий центр разнообразия зеленых лягушек, семенники, фертильность.