  |
||||
|
|
Дмитро Шабанов, Марина Кравченко. |
|
||
|
I. Vertebrates Zoology |
IV.13. Батрахофауна Харківської області: Pelophylax esculentus complex — Гібридогенний комплекс зелених жаб |
IV. Amphibia sensu lato |
||
| ← IV.12. Батрахофауна Харківської області і околиць біостанції (крім зелених жаб) | V.01. Походження, історія і основні групи викопних рептилій → | |||
Фрагмент методички для проведення польової практики з зоології (з невеликими змінами). Частина, що стосується рептилій, написана переважно О.І. Зіненко, а що стосується амфібій — більшою мірою Д.А. Шабановим.
Атемасова Т. А., Влащенко А. С., Гончаров Г.Л., Зіненко О. І., Коршунов О.В., Токарський В. А., Шабанов Д. А., Шандиков Г. А. Навчально-польова практика із зоології хребетних. – Х. : ХНУ імені В. Н. Каразіна, 2019. – 196 с.
Pelophylax esculentus complex — Гібридогенний комплекс зелених жаб
Ймовірно, жодна група амфібій не привертає зараз такої уваги дослідників, як зелені жаби. До складу комплексу середньоєвропейських зелених жаб Pelophylax esculentus complex входять ставкова, Pelophylax lessonae (Camerano, 1882) і озерна, Pelophylax ridibundus (Pallas, 1771) жаби, а також їх гібрид — їстівна жаба, Pelophylax esculentus (Linnaeus, 1758).
Коректно кажучи, нема підстав рахувати гібридних жаб Pelophylax esculentus як окремий вид; втім, їх можна розглядати як гібридну форму, до якої використовується назва, що аналогічна видовій.
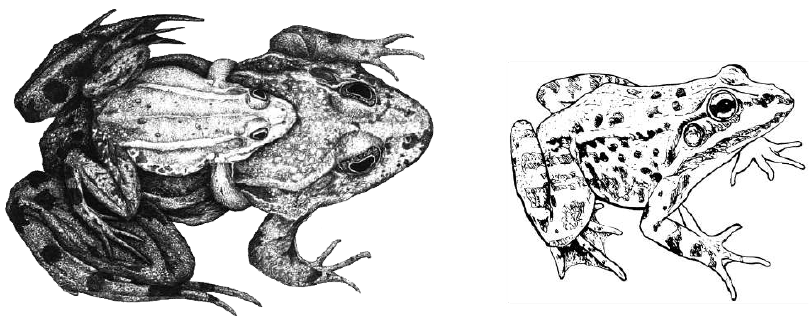
Гібридизація між самцем ставкової жаби та самицею озерної (ліворуч), а також результат цієї гібридизації — міжвидовий гібрид, їстівна жаба (праворуч)
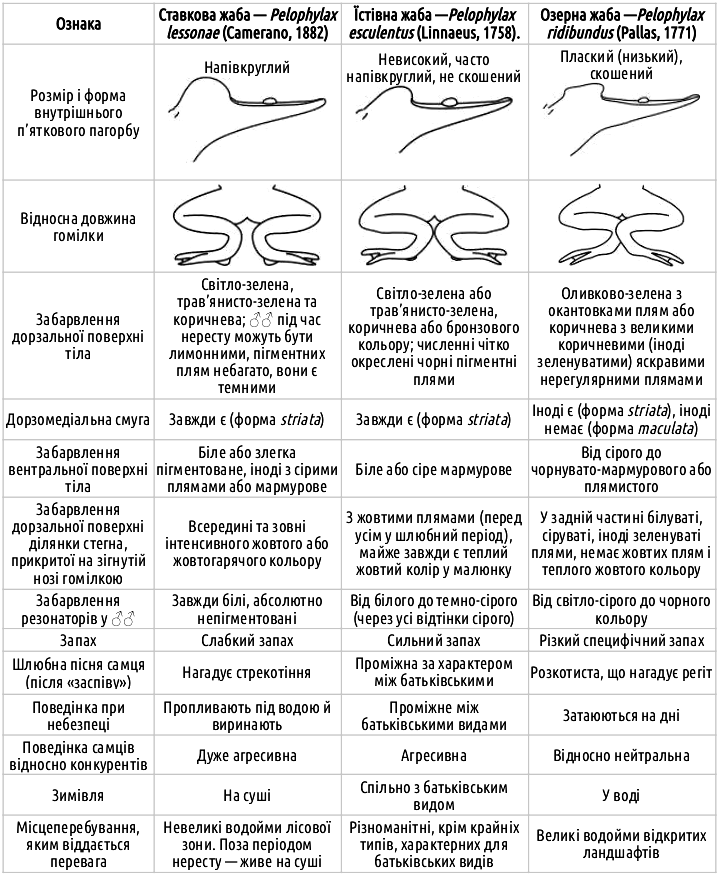
Характерні ознаки представників зелених жаб
Шлюбний спів самця Pelophylax lessonae — ставкової жаби
Шлюбний спів самця Pelophylax esculentus — їстивної жаби
Шлюбний спів самця Pelophylax ridibundus — озерної жаби
Родову назву Pelophylax було відновлено відносно недавно, і в багатьох роботах можна зустріти інші назви, аналогічні щойно наведеним: Rana esculenta complex, Rana lessonae Camerano, 1882, Rana ridibunda Pallas, 1771 і Rana esculenta Linnaeus, 1758. Зверніть увагу, що змінюється не тільки родове ім'я, а й закінчення родового епітета. Крім того, якщо ці види розглядаються в рамках тих родів, в які їх включали автори описів (Лінней, Паллас і Камерано), імена авторів описів і дати публікації в дужки не беруть; якщо ці тварини розглядаються в рамках інших родів, прізвища авторів і дати опису треба брати в дужки.
Історично гібриди були описані першими (ім'я їм дав сам Лінней). Оскільки гібридні жаби демонструють здатність до самостійного існування, вони зберігають ліннеївське ім'я. Звичайно, вони не є видом в звичайному розумінні цього слова. Для такої категорії об'єктів було запропоновано назву «клептон»; у багатьох роботах цей статус підкреслювали, приводячи перед «видовим» ім'ям скорочення «kl.». Тому назва Rana kl. esculenta Linnaeus, 1758 також є синонімом Pelophylax esculentus (Linnaeus, 1758).
Слід підкреслити, що в багатьох випадках точне визначення виду зелених жаб за зовнішніми ознаками виявляється неможливим. Особливо це характерно для регіонів (як наприклад, Сіверсько-Донецького центру різноманіття зелених жаб), де мешкають не тільки диплоїдні, а й триплоїдні Pelophylax esculentus.

Батьківські види зелених жаб відрізняються досить виразно, але відрізнити їх від гібридів (а також розрізнити ди- та триплоїдних гібридів) у деяких випадках буває важно. Зліва направо: озерна, їстівна та ставкова жаби
Геміклональне спадкування у зелених жаб
Міжвидова гібридизація і відтворення гібридів у зелених жаб — наукова проблема, що викликає винятковий інтерес у багатьох вчених як в нашій країні, так і в багатьох інших.
Батьківські види зелених жаб та їх гібриди здатні населяти одні оселища (місцеперебування) і спільно розмножуватися, утворюючи при цьому мішані популяційні системи. Існування таких популяційних систем виявляється можливим завдяки геміклональному (напівклональному) спадкуванню у гібридних жаб. Щоб підкреслити цю обставину, такі сукупності особин різних батьківських видів та їх гібридів називають геміклональними популяційними системами (ГПС).
Геміклональне спадкування виявляється можливим завдяки зміні типового гапло-диплоїдного життєвого циклу з рекомбінацією і редукцією числа хромосом в мейозі, який характерний для еукаріотичних організмів із статевим розмноженням. Щоб зрозуміти значення особливостей спадкування, характерних для зелених жаб, слід порівняти їх з тим випадком, який ми вважаємо типовим. Наприклад, у людини (як і у більшості еукаріот взагалі) чергуються гаплоїдна і диплоїдна фази життєвого циклу. Запліднення забезпечує не тільки перехід від гаплоїдної фази до диплоїдної, але і з'єднання двох батьківських геномів, кожен з яких є випадковою вибіркою з генотипів батьків.
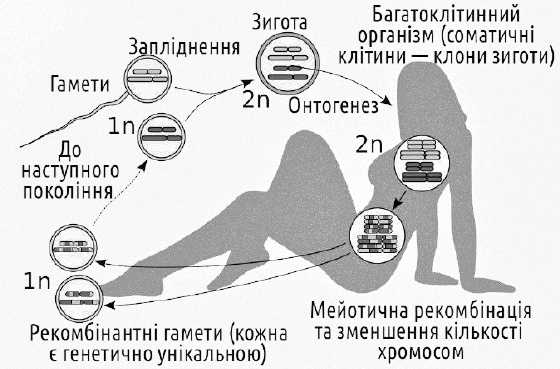
Типовий гапло-диплоїдний життєвий цикл еукаріот з рекомбінацією та редукцією кількості хромосом в мейозі
Геміклональне спадкування зареєстроване у міжвидових гібридів, у яких ступінь відмінності батьківських геномів такий, що мітоз йде нормально, а мейоз (в його типовому варіанті) виявляється неможливим внаслідок порушення впізнавання між хромосомами різних видів.
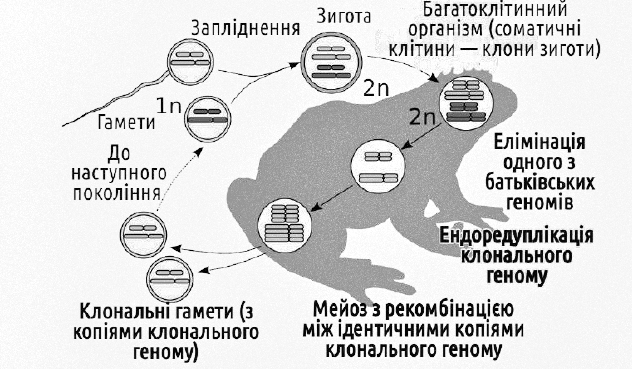
Процеси, що забезпечують геміклональне спадкування, у диплоїдних міжвидових гібридів зелених жаб. На відміну від попереднього рисунку, різниця кольорів умовних хромосом відбиває не їх різну індивідуальність, а їх різну видову приналежність
Нормальне проходження мейозу у таких диплоїдних гібридних жаб виявляється можливим завдяки двом цитогенетичним феноменам, характерним для клітин зародкового шляху — попередників гамет, статевих клітин. Перший незвичайний феномен — елімінація (видалення) одного з геномів. Другий — ендоредуплікація (подвоєння всередині інтерфазного ядра в ході так званого ендомітозу) клонального геному. В результаті ендоредуплікаціі з гаплоїдних клітин знову виходять диплоїдні, що мають дві копії одного і того ж геному. Далі в таких клітинах відбувається нормальний мейоз і гаметогенез. В ході мейозу відбувається кросинговер і обмін ділянками між двома ідентичними геномами, однак, оскільки геноми однакові, кроссинговер не призводить до підвищення генетичної різноманітності. В кінцевому підсумку всі гамети несуть однакові клональні геноми. Таким чином, геміклональність можна розглядати як закономірний (хоча й рідкісний) шлях подолання стерильності міжвидових гібридів. Двома іншими (і суттєво поширенішими) шляхами подолання гібридної стерильності є поліплоїдізація та перехід до клонального відтворення гібридів.
Наслідком зміни базових особливостей життєвого циклу у геміклональних гібридів стає глибока зміна всієї їх біології. Вони існують не у складі моновидових популяцій, а в ГПС — геміклональних популяційних системах. Для них характерна інша генеалогія (не сітчаста, коли кожна особина отримує і передає спадкову інформацію від обох батьків, а фактично лінійна, коли у спадок передається тільки один геном, отриманий від одного з батьків). Різна доля геномів і різний характер добору, який на них діє, можуть призводити до їх функціональної диференціації. Незвичайними виявляються і підтримка стійкості ГПС, і еволюція батьківських видів та їх гібридів.
Щоб описати такий характер спадкування, використовують форму запису, в якій геном ставкової жаби позначають L, а озерної — R (за першими літерами їх латинських назв). Слід пам'ятати, що ці позначення мають відношення не до окремих генів, а до цілих геномів, кожен з яких складається з 13 хромосом. Символ геному, який передається клонально, беруть в дужки. Жіночі та чоловічі геноми позначають відповідно надстроковими індексами X і Y (у жаб, як і у людей, гетерогаметною є чоловіча стать, хоча чоловічий і жіночий хромосомні набори морфологічно не відрізняються). Так, запис ♂XR(YL) позначає диплоїдного гібридного самця P. esculentus, який клонально передає чоловічий геном P. lessonae, а ♀XRXR — звичайну самицю P. ridibundus.
Геміклональне спадкування у зелених жаб було відкрито завдяки дослідженням польського зоолога Лешека Бергера, перші результати яких були оприлюднені у 1964 році. Для західноєвропейських ГПС є характерними гібриди, що клонально передають геноми P. ridibundus.
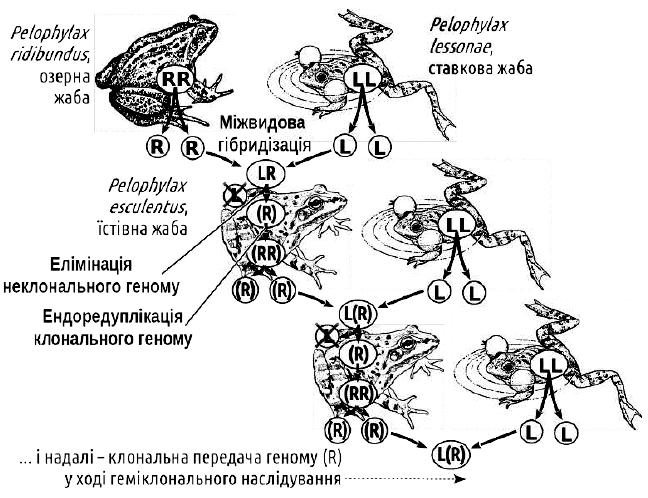
Так відбувається поява і відтворення гібридних зелених жаб у багатьох регіонах Західної Європи. Процеси, що відбуваються в ГПС Східної України, є більш складними
Сіверсько-Донецький центр різноманіття Pelophylax esculentus complex
Гібридизація між батьківськими видами зелених жаб з появою P. esculentus відбувається на найширшому ареалі, що простягнувся від Франції до Поволжя. На цій території зареєстровані вельми різні за своїм складом ГПС. У Західній Європі до їх складу часто входять триплоїдні гібриди; на пострадянській території знахідки гібридів були поодинокими доти, поки в 2002-2004 рр. не було відкрито зону масового поширення тріплоїдних гібридів, що пов'язана з річкою Сіверський Донець.
Складність ГПС зелених жаб, описаних у Сіверському Донці, є безпрецедентною. Наприклад, у Сіверському Донці поруч з біостанцією Харківського національного університету імені В.Н.Каразіна та на території НПП «Гомільшанські ліси» поширені:
— представники одного з батьківських видів: ♀♀ и ♂♂ RR;
— диплоїдні гібриди: ♀♀ и ♂♂ L(R), (L)R і (L)(R);
— триплоїдні гібриди: ♀♀ и ♂♂ LLR і LRR;
— поодинокі незрілі тетраплоїди LLRR і представники іншого батьківського виду, LL, які не доживають до статевої зрілості.
Для позначення типів ГПС використовуються літери, що позначають форми жаб у їх складі: L і R — батьківські види, а E — P. esculentus. Позначка p відповідає наявності в ГПС поліплоїдів (триплоїдів), а позначки f або m — випадку, коли певна форма представлена лише самицями або самцями відповідно. Якщо певна форма представлена лише однією геномною композицією, це можна позначити відповідним індексом. Таким чином, можна зазначити, що поруч з біостанцією у заплаві мешкає R-E-Epf-ГПС.
Як це не парадоксально, статевозрілі P. lessonae у басейні Сіверського Донця не знайдені; усі геноми L передаються тут через гібридів. При схрещуванні між собою гібридів, що передають клональні геноми одного батьківського виду можуть виникати особини цього виду, які зазвичай виявляються нежиттєздатними: R(L) × R(L) → (L)(L) → †. Цей ефект найчастіше розглядають як наслідок так званого «храповика Мюллера» — накопичення генетичних дефектів в геномах, які передаються без рекомбінації. Втім, це пояснення не є вичерпним описом спостережуваного феномена повною мірою, адже нежиттєздатними виявляються і особини, що отримали геноми одного виду, що належать до різних клонів.
Оскільки усі геноми P. lessonae передаються в басейні Сіверського Донця лише через гібридних особин, дія «храповика Мюллера» мала б привести до їх деградації. Вірогідно, це не відбувається через рекомбінацію L-геномів під час гаметогенезу гібридів LLR.
Викладені обставини привели до опису Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex. У його складі виділено три субрегіони, що відрізняються за характерними ГПС зелених жаб.
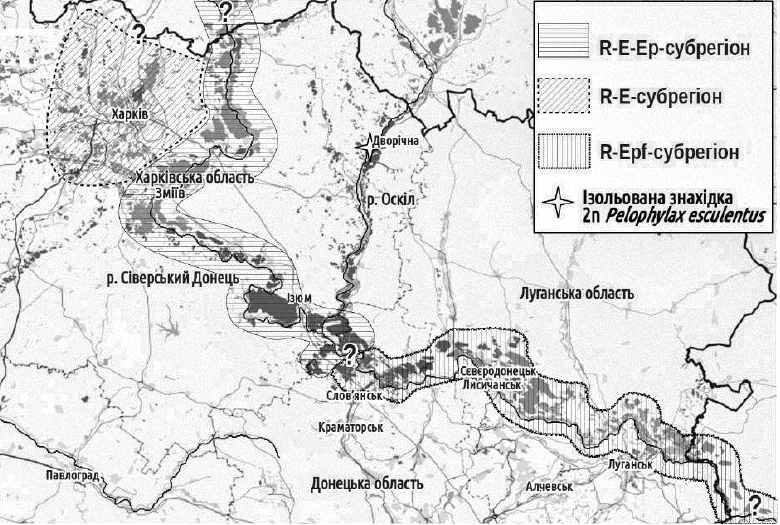
Географічне розташування і склад субрегіонів Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex
R-E-Ep-субрегіон розташований вздовж Сіверського Донця у межах Харківської області, охоплюючи також пов’язані з Донцем водойми. Склад R-E-Ep-ГПС описано вище. Деякі ГПС в цьому субрегіоні можуть відрізнятися за своїм складом. Особливо цікаву історію вивчення має ГПС Іськова ставку, розташованого в околицях селища Гайдари.
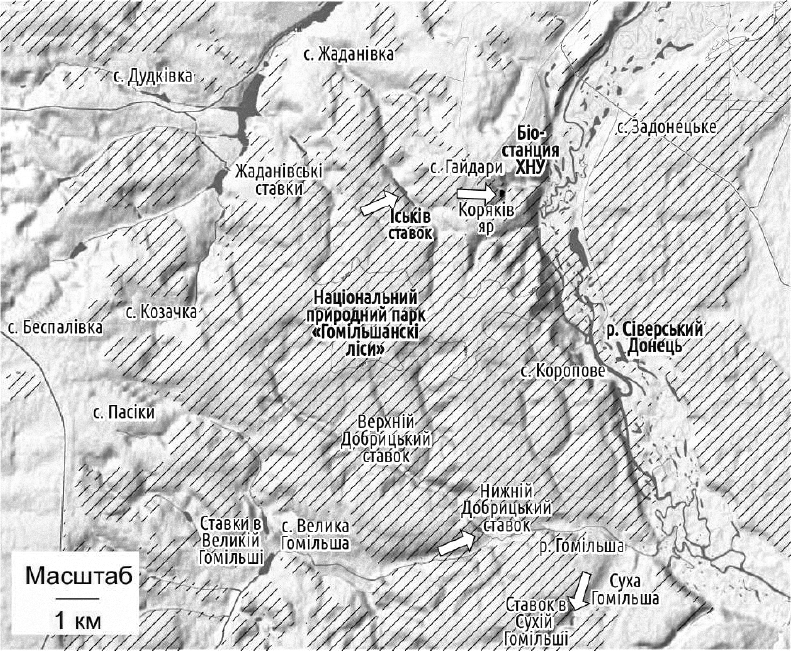
Кілька цікавих для вивчення ГПС зелених жаб знаходяться у “Великих Гайдарах”, поруч з біостанцією ХНУ імені В. Н. Каразіна и НПП «Гомільшанські ліси». Водойми, які населяють ці ГПС, показано стрілками; ліси позначені штрихуванням
В 1995-1996 рр. в Іськовому ставку перебувала ГПС, що складалася майже виключно з диплоїдних P. esculentus. Після низки пертурбацій в цій ГПС, як і колись, переважають диплоїдні P. esculentus. На жаль, на час написання цього посібника чисельність цієї ГПС стала критично низькою. Знахідки самиць P. ridibundus, а також триплоїдних P. esculentus не дають змоги розглядати цю систему як справжню E-ГПС; втім, вона є REEp ГПС, що є близькою до E-HPS. Згідно з висунутою гіпотезою, головний тип схрещування, який реалізується в ГПС Іськова ставку, є таким:
♂(YL)(XR) × ♀(XL)(XR) ⟶ ♂(XL)(YL) : ♂(YL)(XR) : ♀(XL)(XR) : ♀(XR)(XR) ⟶ ♂(YL)(XR) : ♀(XL)(XR).
Згідно з цим припущенням, переважну кількість особин в цій ГПС складають два амфігаметні клони P. esculentus, жіночій та чоловічий. Внаслідок їх схрещування утворюються 4 форми потомства. Дві з них отримують клональні геноми одного виду і гинуть; два інших є ідентичними батьківським особинам. Якщо це припущення є правильним, ми маємо справу з біосистемою, характер відтворення якої є унікальним. Фактично, це два амфігаметні клони, що підтримують один одного.
R-E-Ep-субрегіон розташовано у басейнах двох правих притоків Сіверського Донця — річок Уди та Мжа. На території цього субрегіону знаходиться і м. Харків. Останнім часом в басейні річки Мжа також знайдено триплоїдів.
R-Epf-субрегіон пов’язаний з Сіверським Донцем у Донецькій та Луганській областях. Тут трапляються ГПС, що складаються з ♀RR, ♂RR і ♀LLR. Механізм відтворення гібридів в цих ГПС потребує детального вивчення.
В даний час в Харківському університеті працює група популяційної екології амфібій, що досліджує механізми відтворення ГПС зелених жаб Сіверсько-Донецького центру їх різноманітності; в цій роботі активну участь беруть студенти.