Ниже приведен русский перевод автореферата. Украинский текст автореферата в формате pdf лежит здесь.
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ УКРАИНЫ
ДНЕПРОПЕТРОВСКИЙ НАЦИОНАЛЬНЫЙ УНИВЕРСИТЕТ ИМЕНИ ОЛЕСЯ ГОНЧАРА
ШАБАНОВ ДМИТРИЙ АНДРЕЕВИЧ
УДК (597.841::574.4):[477.5+292.485]
ЭВОЛЮЦИОННАЯ ЭКОЛОГИЯ ПОПУЛЯЦИОННЫХ СИСТЕМ
ГИБРИДОГЕННОГО КОМПЛЕКСА ЗЕЛЕНЫХ ЛЯГУШЕК
(PELOPHYLAX ESCULENTUS COMPLEX)
ЛЕВОБЕРЕЖНОЙ ЛЕСОСТЕПИ УКРАИНЫ
03.00.16 — экология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени доктора биологических наук
Днепропетровск — 2015
Диссертация является рукописью
Работа выполнена на кафедре зоологии и экологии животных биологического факультета Харьковского национального университета имени В. Н. Каразина Министерства образования и науки Украины
Официальные оппоненты доктор биологических наук, профессор
Серебряков Валентин Валентинович,
Киевский национальный университет
имени Тараса Шевченко, кафедра зоологии, заведующий;
доктор биологических наук, профессор
Домнич Валерий Иванович,
Запорожский национальный университет,
кафедра охотоведения и ихтиологии, заведующий;
доктор биологических наук, доцент
Жуков Александр Викторович
Днепропетровский национальный университет
имени Олеся Гончара, кафедра зоологии и экологии, профессор.
Защита состоится «_26_» марта 2015 года в _10.00_ на заседании специализированного ученого совета Д 08.051.04 для защиты диссертаций на соискание научной степени доктора биологических наук в Днепропетровском национальном университете имени Олеся Гончара Министерства образования и науки Украины по адресу: 49010, г. Днепропетровск, проспект Гагарина, 72, корпус 17, факультет биологии, экологии и медицины, ауд. 611.
С диссертацией можно ознакомиться в научной библиотеке Днепропетровского национального университета имени Олеся Гончара по адресу: м. Днепропетровск, ул. Казакова, 8.
Автореферат разослан «_26_» _февраля_ 2015 г.
 Ученый секретарь специализированного ученого совета,
Ученый секретарь специализированного ученого совета,
кандидат биологических наук, доцент А. А. Дубина
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Пятьдесят лет назад, в 1964 году польский гидробиолог Лешек Бергер обнаружил, что классический объект экспериментальной биологии, известные еще Карлу Линнею съедобные лягушки, Pelophylax esculentus (Linnaeus, 1758), не являются обычным видом (Berger, 1964). Это межвидовой гибрид лягушки прудовой, Pelophylax lessonae (Camerano, 1882) и лягушки озерной, Pelophylax ridibundus (Pallas, 1771). Усилиями группы европейских исследователей было показано, что воспроизводство Pelophylax esculentus поддерживается благодаря гемиклональному наследованию (клональной передаче из поколения в поколение отдельных геномов родительских видов в относительно неизменном виде). В настоящее время установлено, что гемиклональное наследование потребовало изменения базовых механизмов, обеспечивающих воспроизводство подавляющего большинства эукариотических организмов. Необычность зеленых лягушек проявляется на всех уровнях их биологии: от функционирования их популяционных систем и способа эволюции до механизмов работы их клеток. Изучение столь необычного феномена позволяет расширить наше понимание закономерностей, относящихся к сфере популяционной и эволюционной экологии, а также целого ряда других биологических наук.
Десять лет назад при участии диссертанта в Харьковской области, в лесостепной части бассейна реки Северский Донец были описаны необычные по своему составу популяционные системы зеленых лягушек, Pelophylax esculentus complex (Mass…, 2004; Массовая..., 2005; Изучение…, 2006). За время их изучения стало ясно, что они принадлежат к числу самых сложных из числа известных науке. Изучение популяционных систем гибридогенного комплекса зеленых лягушек в лесостепной части бассейна Северского Донца потребовало создания на кафедре зоологии и экологии животных Харьковского национального университета имени В. Н. Каразина рабочей группы, научным руководителем которой является диссертант. Данное исследование призвано интегрировать результаты, полученные рабочей группой при участии диссертанта, и создать теоретические рамки, организующие дальнейшее изучение открытого в ходе совместной работы естественного феномена.
К началу выполнения диссертационного исследования состав популяций и популяционные систем представителей Pelophylax esculentus complex Левобережной лесостепи Украины был изучен крайне неравномерно. Состав этих систем в части Левобережной Лесостепи, расположенной в водосборном бассейне Днепра (Левобережно-Днепровской физико-географической провинции), был установлен в ряде работ, выполненных с использованием электрофореза белковых маркеров и иными методами сотрудниками Института зоологии имени И. И. Шмальгаузена НАНУ (Морозов-Леонов, 1998; Некрасова, Морозов-Леонов, 2001; Некрасова, 2002). Поэтому в нашей работе достаточно было изучить состав таких систем только для восточной части этой провинции. Расположенную в бассейне Северского Донца (притока Дона) Среднерусскую физико-географическую провинцию, относящуюся к Лесостепи Украины, следовало исследовать максимально полно, в связи с ее недостаточной изученностью. Кроме того, популяционные системы Pelophylax esculentus complex, аналогичные тем, которые регистрируются в Лесостепи Украины, встречаются и в той части Степи, где распространены элементы лесостепных ландшафтов. Поэтому часть исследований необходимо было выполнить в Донецко-Донской северостепной физико-географической провинции.
Связь работы с научными программами, планами, темами. Работа выполнена в соответствии с научно-исследовательскими темами кафедры зоологии и экологии животных биологического факультета Харьковского национального университета имени В. Н. Каразина: «Неортодоксальное видообразование (гибридизация, неменделевское наследование, полиплоидия и сетчатая эволюция): изучение на примере амфибий» (№ госрегистрации 0109U007439; 2009-2010 гг.); «Разработка технологий получения клонального потомства позвоночных животных путем скрещивания полуклональных межвидовых гибридов (на примере зеленых лягушек)» (№ госрегистрации 0112U001026; 2012-2014 гг.).
Цель и задачи исследования. Цель работы — выяснить закономерности функционирования и эволюции популяционных систем Pelophylax esculentus complex Левобережной лесостепи Украины.
Для достижения указанной цели в работе поставлены следующие задачи:
— установить состав популяционных систем Pelophylax esculentus complex, характерных для Среднерусской физико-географической провинции, для восточной части Левобережно-Днепровской физико-географической провинции, а также для прилежащей к Лесостепи части Донецко-Донской северостепной физико-географической провинции;
— выделить в регионе исследования отдельные субрегионы, отличающиеся по составу распространенных в них популяционных систем Pelophylax esculentus complex; выявить биогеоценотические и географические факторы, определяющие распределение по их территории популяционных систем Pelophylax esculentus complex;
— определить те из субрегионов исследования, которые представляют наибольший интерес для изучения и имеют наибольшую ценность с точки зрения сохранения их биоразнообразия; установить характерные особенности распространенных в этих субрегионах представителей Pelophylax esculentus complex;
— выяснить специфику биосистем, в которых происходит гемиклональное воспроизводство Pelophylax esculentus;
— разработать способы изучения устойчивости и закономерностей трансформаций популяционных систем Pelophylax esculentus complex с использованием имитационного моделирования;
— построить динамическую типологию популяционных систем, включающих диплоидных представителей Pelophylax esculentus complex;
— определить уровни отбора, влияющего на динамику биосистем, в которых происходит гемиклональное воспроизводство Pelophylax esculentus; установить возможные механизмы поддержки отбором гемиклонального воспроизводства гибридов в Pelophylax esculentus complex.
Объект исследования — популяционные системы Pelophylax esculentus complex Левобережной лесостепи Украины.
Предмет исследования — закономерности функционирования и поддержания устойчивости популяционных систем Pelophylax esculentus complex, их разнообразие и эволюционное значение.
Методы исследования:
— качественное определение типа популяционных систем зеленых лягушек по составу обитающих в них форм;
— формализованное описание местообитаний популяций и популяционных систем Pelophylax esculentus complex,
— оценка численности и состава популяционных систем с использованием мечения и повторного отлова;
— первичное определение видовой принадлежности особей по признакам внешней морфологии;
— экспресс-определение плоидности особей с использованием цитометрии;
— скелетохронологическое определение возраста особей и ретроспективная оценка скорости их роста на основании анализа размера линий склеивания на срезах трубчатых костей;
— сравнительные экологические, морфологические, биохимические и физиологические исследования представителей Pelophylax esculentus complex;
— генотипирование особей с использованием проточной ДНК-цитометрии, электрофореза белковых маркеров;
— кариоанализ раздавленных препаратов мацерированных тканей, раскапанных метафазных пластинок и раскапанных клеток;
— определение геномного состава гамет с использованием проточной ДНК-цитометрии, электрофореза белковых маркеров, анализа маркеров хромосом типа ламповых щеток и анализа результатов скрещиваний;
— имитационное моделирование и эксперименты с имитационными моделями; статистическая обработка результатов моделирования (анализ методом главных компонент);
— сравнение результатов моделирования с эмпирическими данными, формализация эмпирических и литературных данных.
Научная новизна полученных результатов. Результаты, отражающие научную новизну, заключаются в следующем: впервые:
— в Левобережной лесостепи Украины обнаружены регионы, где массово распространены полиплоидные Pelophylax esculentus;
— в регионе исследования выделены пять субрегионов, которые отличаются по набору типов популяций и популяционных систем представителей Pelophylax esculentus complex, распространенных на их территории;
— установлены биогеоценотические связи, определяющие состав локальных популяционных систем Pelophylax esculentus complex внутри каждого из описанных субрегионов;
— обнаружен и описан Северско-Донецкий центр разнообразия Pelophylax esculentus complex;
— предложено понятие «гемиклональная популяционная система» (ГПС) и обосновано, что ГПС являются особой категорией биосистем, характерной для гибридогенных комплексов видов;
— обосновано, что трансформации ГПС являются особым типом динамики биосистем; установлены ключевые факторы, определяющие эти трансформации;
— разработана концептуальная модель трансформаций ГПС, на основе которой созданы имитационные модели этих процессов;
— разработана динамическая типология ГПС Pelophylax esculentus complex, включающих диплоидные межвидовые гибриды;
— с помощью имитационного моделирования ГПС Северско-Донецкого центра разнообразия Pelophylax esculentus complex, состоящих из диплоидных представителей, сформулированы доступные для проверки предположения о составе передающихся в них клональных геномов;
— выдвинута гипотеза, объясняющая феномен гибридной амфиспермии конкуренцией в клетках зародышевого пути клональных геномов, измененных вследствие отбора на их способность вызывать элиминацию альтернативных геномов;
— различия в устойчивости разных типов ГПС Pelophylax esculentus complex рассмотрены как проявление группового отбора;
— выдвинута гипотеза, рассматривающая способность к гемиклональной гибридизации как приспособление к гомеологичной межвидовой рекомбинации, поддерживаемой отбором видов;
получили дальнейшее развитие:
— представления о разнообразии популяционных систем Pelophylax esculentus complex;
— классификация типов популяционного воспроизводства и представления о связи этих типов с особенностями биосистем индивидуального, популяционного и видового уровней;
— способы использования имитационных моделей для исследования эмпирически зарегистрированного разнообразия естественных биосистем;
— понятийная база для описания и моделирования популяционных систем гибридогенных комплексов видов с гемиклональным наследованием;
— представления о формах типологии биосистем и способах построения динамических типологий сложных систем;
— концепция устойчивости биосистем, как их способности сохранять свой динамический тип с течением времени;
— концепция многоуровневого отбора;
— представления об отборе клеточных линий в гонадах Pelophylax esculentus;
— теоретические представления о закономерностях эволюции рекомбинации;
— концепция отбора видов.
Практическое значение полученных результатов. Полученные в ходе работы результаты важны для планирования и организации охраны уникальных популяционных систем Pelophylax esculentus complex. Разработанные способы использования имитационных моделей позволяют оценивать устойчивость сложных биологических систем и полезны для тестирования гипотез, компенсирующих недостаток эмпирических данных. Продемонстрирована возможность использования имитационных моделей для разработки динамической типологии биосистем. Полученные в ходе работы данные и обобщения используются при преподавании общих и специальных курсов «Экология», «Моделирование в экологии», «Батрахология и герпетология», «Биологическая статистика в зоологии и экологии», «Зоология позвоночных», а также при проведении учебно-полевой практики по зоологии позвоночных на биологическом факультете ХНУ имени В. Н. Каразина.
Личный вклад соискателя. Диссертационная работа является самостоятельным и оригинальным исследованием.
Ряд публикаций, отражающих рассмотренные в диссертации темы, подготовлены в составе рабочей группы популяционной экологии амфибий ХНУ имени В. Н. Каразина. Автор является научным руководителем этой группы; он играл руководящую роль при постановке задачи для исследования, принимал личное участие во всех этапах работы, в том числе, полевых, и руководил интерпретацией результатов. Некоторые из результатов, представленных в данной работе, были частично отражены в кандидатских диссертациях А. В. Коршунова (2010) и М. А. Кравченко (2013), выполненных под руководством автора. Ряд публикаций подготовлен в результате международного сотрудничества с научными коллективами России, Германии и других стран. В этих публикациях автор принимал участие в постановке задачи, лично собирал материал, принимал участие в его обработке и интерпретации результатов.
Апробация результатов диссертации. Материалы диссертации обнародованы на III, IV и V Международных научных конференциях Zoocenosis-2005, Zoocenosis-2007 и Zoocenosis-2009 (Днепропетровск); научной конференции «Современные проблемы зоологии и экологии» (Одесса, 2005); IX и XI Международных научно-практических экологических конференциях, посвященных популяционной экологии (Белгород, 2006 и 2010); III и V съездах Герпетологического общества им. А. М. Никольского (Пущино, 2006; Минск, 2012); научной конференции биологического факультета ХНУ, посвященной 100-летию со дня рождения Е.И. Семененко (Харьков, 2006); Международной конференции «Современные проблемы биологической эволюции» (Москва, 2007); III, IV, V, VI и VII съездах Украинского герпетологического общества (Мелитополь, 2007; Днепропетровск, 2008; Харьков, 2010; Киев, 2011; Вилково, 2013); III Международной конференции молодых ученых «Биология: от молекулы до биосферы» (Харьков, 2008); Международной научно-практической конференции «Биоразнообразие и устойчивое развитие» (Симферополь, 2010); І Всеукраинской научно-практической конференции «Современная проблематика и методология биоэкологических исследований: популяционный подход» (Ивано-Франковск, 2012); конференции-школе «Аномалии и патологии амфибий и рептилий: методология, эволюционное значение, возможность оценки здоровья среды» (Екатеринбург, 2013); конференции, посвященной 100-летию Биологической станции ХНУ имени В. Н. Каразина (Гайдары, 2014).
Публикации. По материалам диссертации опубликована 51 научная публикация, в том числе 31 статья в профильных научных журналах и сборниках научных трудов, из них — 9 в изданиях, включенных в международные библиографические базы, 7 статей в других изданиях, а также 14 научных публикаций в материалах и тезисах конференций.
Структура и объем диссертации. В состав диссертации входит перечень условных обозначений, введение, 9 глав, заключение, выводы и список использованной литературы. Работа содержит 36 таблиц і 62 рисунка. Материалы диссертации изложены на 364 страницах компьютерного набора, из которых основного текста — 275 страниц. Список использованной литературы содержит 419 источников, из которых 125 — латиницей.
ПЕРЕЧЕНЬ УСЛОВНЫХ ОБОЗНАЧЕНИЙ И ОПРЕДЕЛЕНИЯ СПЕЦИФИЧЕСКИХ ТЕРМИНОВ
Термины приведены в логическом, а не в алфавитном порядке.
|
Pelophylax esculentus complex |
Группа европейских зеленых лягушек: прудовая лягушка, Pelophylax lessonae (Camerano, 1882), озерная лягушка Pelophylax ridibundus (Pallas, 1771), а также различные формы их межвидовых гибридов: съедобная лягушка, Pelophylax esculentus (Linnaeus, 1758), для которой используется название, аналогичное видовому |
|
Гемиклональное наследование |
Наследование у межвидовых гибридов, при котором один из родительских геномов (клональный геном) передается (в типичном случае) в гаметы целиком, без рекомбинации |
|
Клональные геномы: (L), (R) |
Геномы, передающиеся потомству как целое, без рекомбинации: (L) — геном P. lessonae, (R) — геном P. ridibundus |
|
Рекомбинантные геномы: L, R |
Геномы, передающиеся с рекомбинацией (как у родительских видов): L — геном P. lessonae, R — геном P. ridibundus |
|
XL, XR, (XL), (XR) |
Женские геномы (несущие X-хромосому) |
|
YL, YR, (YL), (YR) |
Мужские геномы (несущие Y-хромосому) |
|
Кариогенетические формы |
Формы представителей P. esculentus complex, отличающиеся по составу геномов в их генотипе и в продуцируемых ими гаметах |
|
R(L), L(R), (L)(R) |
Диплоидные P. esculentus с разными клональными геномами |
|
LLR, LRR |
Триплоидные P. esculentus |
|
ГПС, |
Совокупность совместно обитающих и размножающихся представителей различных форм гибридов и особей родительских видов, в которой из поколения в поколение передаются как клональные, так и рекомбинантные геномы |
|
Трансформации ГПС |
Форма динамики ГПС, заключающаяся в изменении в них численности и соотношения различных кариогенетических форм |
|
R-E-L-тип ГПС; R-Ep-тип ГПС |
Обозначения типов ГПС в аналитической классификации, где буква R обозначает P. ridibundus, E — P. esculentus, а L — P. lessonae, а литерой p обозначается наличие в ГПС полиплоидных P. esculentus |
|
Конспецифичный геном |
Клональный геном того же вида, что и преобладающий родительский вид в ГПС (например, (R) в ГПС R-E-типа); понятие применимо для ГПС R-E- и L-E-типа |
|
Гетероспецифичный геном |
Клональный геном иного вида, чем преобладающий родительский вид в ГПС (например, (L) в ГПС RE-типа); понятие применимо для ГПС RE- и LE-типа |
|
Размножение |
Феномен организменного уровня организации биосистем, связанный с появлением у организма потомков; служит основой популяционного воспроизводства |
|
Воспроизводство |
Феномен популяционного уровня организации биосистем, связанный с поддержанием определенного состава вследствие размножения организмов |
|
Гибридолиз |
Появление особей родительского вида в результате скрещивания гибридов, клонально передающих геномы одного вида |
|
Гибридная амфиспермия |
Производство межвидовыми гибридами (P. esculentus) смеси гамет, часть из которых несет геном P. lessonae, а часть — P. ridibundus. Диплоидные гибриды с гибридной амфиспермией в данной работе обозначаются (L)(R). |
|
Клон |
Совокупность клеток или особей с идентичным (с точностью до ошибок копирования) генотипом. Организм человека в типичном случае является клоном клеток; однояйцевые близнецы являются организмами-клонами |
|
Гемиклон |
Совокупность особей, имеющих идентичный (с точностью до ошибок копирования) клональный геном и различные рекомбинантные геномы |
ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ
СОВРЕМЕННОЕ СОСТОЯНИЕ ПРОБЛЕМЫ
Приведено краткое описание истории открытия и изучения гибридогенеза в Pelophylax esculentus complex, а также обсуждение его экологического значения. Показано, что типичный для эукариот жизненный цикл (рис. 1.А.) является причиной характерной для них иерархии биосистем (рис. 2.А.) Гемиклональное наследование, характерное для P. esculentus и других межвидовых гибридов, является глубоким видоизменением этого цикла (рис. 1.Б.). Вопрос, как такое видоизменение отражается на различных уровнях биосистем, в том числе рассматриваемых в рамках экологии, остается недостаточно изученным.
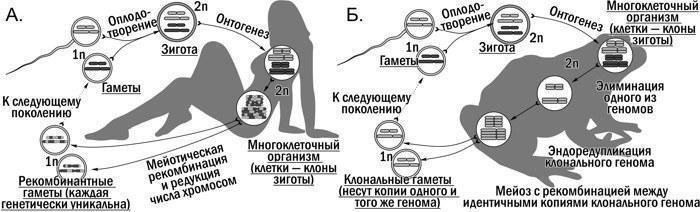
Рис. 1. Сравнение характерного для эукариот гапло-диплоидного жизненного цикла с оплодотворением и мейозом (А.) и видоизменения типичного жизненного цикла, характерного для межвидовых гибридов с гемиклональным наследованием (Б.)
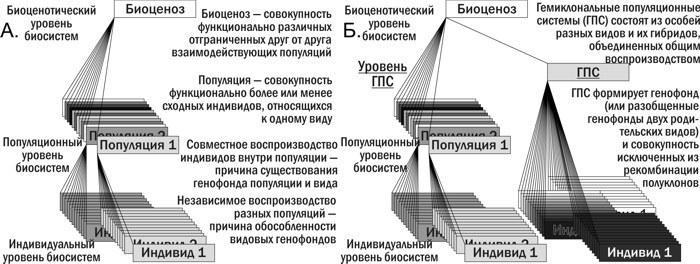
Рис. 2. А. Иерархия биосистем, типичная для эукариот с половым размножением, и некоторые особенности характерных для них биосистем (глава 1). Б. Видоизменения иерархии биосистем, характерные для гибридогенных комплексов видов с гемиклональным наследованием (глава 6)
Биосистемы, в которых воспроизводятся P. esculentus, называют (Uzzell, Berger, 1975) популяционными системами. Распространенная типология отражает набор форм лягушек в составе систем: L-тип — популяции P. lessonae; L-E-тип — P. lessonae и P. esculentus; L-E-R-тип — оба родительских виды и их гибриды; E-тип — так называемые «чистые» популяции P. esculentus т.п. (Plötner, 2005). Наличие среди P. esculentus полиплоидных (3n и 4n) особей обозначают буквой p. Если P. esculentus представлены только самками, используют букву f, а только самцами — m.
Воспроизводство P. esculentus в популяционных системах различного состава (рис. 3) становится возможным потому, что различные кариогенетические формы гибридов производят гаметы, которые могут принадлежать разным родительским видам. Существуют гибриды, способные одновременно производить гаметы обоих видов (явление гибридной амфиспермии). При скрещивании гибридов, передающих клональные геномы одного и того же вида, в потомстве образуются особи этого вида (явление гибридолиза), имеющие пониженную жизнеспособность и, в большинстве случаев, погибающие во время метаморфоза или вскоре после него.
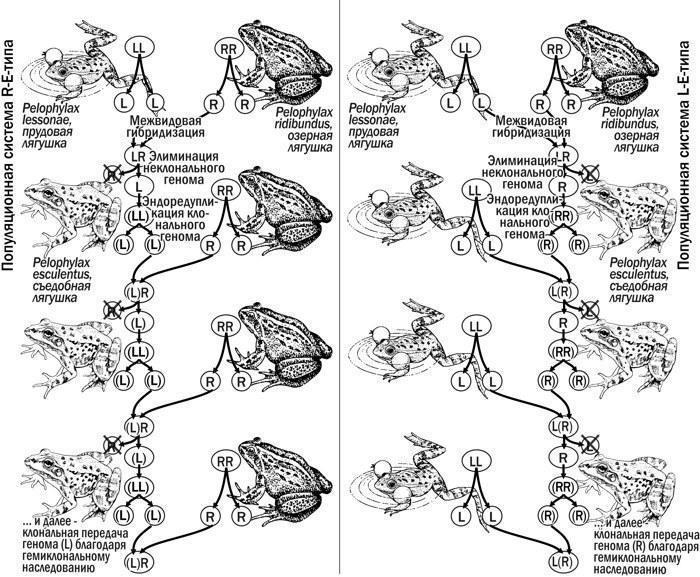
Рис. 3. Воспроизводство P. esculentus как в популяционных системах R-E-типа, так и в системах L-E-типа возможно потому, что различные кариогенетические формы P. esculentus продуцируют гаметы, несущие различные геномы
Приведен краткий очерк истории изучения представителей Pelophylax esculentus complex в биоценозах Левобережной лесостепи Украины, проводится сравнение изученности их популяционных систем в различных физико-географических провинциях Левобережной лесостепи Украины. Обсуждается использование моделирования в популяционной биологии, развитие представлений о разнообразии экологических стратегий у животных и современное состояние дискуссии об уровнях отбора в эволюционной биологии.
МАТЕРИАЛ И МЕТОДЫ ЕГО ИССЛЕДОВАНИЯ
В работе с применением комплекса методов определен состав 134 популяций и популяционных систем Pelophylax esculentus complex, места сбора которых показаны на рис. 6 (стр. 15). На рис. 6 точки, соответствующие локалитетам исследований, наложены на космический снимок, полученный из базы Google Earth, и подписаны условными названиями выборок. Различные области Украины и территория Российской Федерации окрашены в разные цветовые оттенки.
Методами, позволяющими надежную идентификацию состава геномов особей, исследовано более 300 выборок, включающих более 2500 особей зеленых лягушек (из некоторых локалитетов было изучено по несколько выборок). Наиболее подробно изучались популяционные системы, расположенные в окрестностях НПП «Гомольшанские леса» и Северско-Донецкой биологической станции имени профессора В. М. Арнольди ХНУ имени В. Н. Каразина. Оценку численности и состава популяционных систем методом мечения и повторного отлова провели для 3 популяционных систем (Иськов пруд, Нижний Добрицкий пред и пруд в с. Жовтневое, рис. 6). Изучение экологической и географической обусловленности состава выполнили по результатам формализованного описания 102 популяционных систем и популяций, а также особенностей их местообитаний.
Возраст и темпы роста установили методами скелетохронологии для 500 особей, плоидность методом цитометрии — примерно для 1 000 особей. Экспресс-определение принадлежности зеленых лягушек к той или иной форме (табл. 1) провели примерно для 10 000 особей. В подавляющем большинстве случаев перечисленные исследования не требовали изъятия особей из природных местообитаний. Прямыми методами (различные модификации кариоанализа) кариотипы соматических клеток и сперматоцитов определили более чем для 150 особей. Состав гамет методами проточной ДНК-цитометрии спермы определили более, чем для 150 особей, методом анализа хромосом типа ламповых щеток в ооцитах — примерно для 30 особей. Проведено около 50 искусственных скрещиваний, состав потомства определен для 12 выводков. Фертильность методом гормональной стимуляции определили для 70 самцов. Количество итераций в экспериментах с имитационной моделью превысило 130 000.
Приведены подробные описания примененных методов исследования. К методам исследования популяционного и биогеоценотического уровней биосистем принадлежат определение численности популяционных систем и отдельных форм с применением мечения, а также описание особенностей местообитания. К методам организменного уровня относятся предварительное определение таксономической принадлежности особей (табл. 1), методики индивидуального мечения лягушек, скелетохронологическое определение возраста лягушек и ретроспективная оценка скорости роста по скелетохронологическим данным, методика получения уринальной спермы самцов с использованием гормональной стимуляции.
Таблица 1. Признаки трех форм зеленых лягушек, доступные для прямого наблюдения (Лада, 1995; Изучение ..., 2006; Коршунов, 2010, с дополнениями)
|
Признак |
Прудовая лягушка — Pelophylax lessonae (Camerano, 1882) |
Съедобная лягушка — Pelophylax esculentus (Linnaeus, 1758). |
Озерная лягушка — Pelophylax ridibundus (Pallas, 1771) |
|
Размер и форма внутреннего пяточного бугра |
Полукруглый
|
Невысокий, часто полукруглый, не скошенный
|
Плоский (низкий), скошенный
|
|
Относительная длина голени |
|
|
|
|
Окраска дорзальной поверхности тела |
Светло-зеленая, травянисто-зеленая и коричневая, ♂♂ во время нереста могут быть лимонными, пигментных пятен мало и они темные |
Светло-зеленая или травянисто-зеленая, коричневая или цвета бронзы; многочисленные пигментные пятна черные, как правило, четко очерченные |
Оливково-зеленая с окантовками или коричневая с большими коричневыми (реже зеленоватыми) яркими, нерегулярными пятнами |
|
Дорзомедиальная полоса |
Всегда есть (форма striata) |
Всегда есть (форма striata) |
Иногда есть (форма striata), иногда нет (форма maculata) |
|
Окраска вентральной поверхности тела |
Как правило, белая или слегка пигментированная, реже с серыми пятнами или мраморная |
Белая или серая мраморная |
От серой до черновато-мраморной или пятнистая |
|
Окраска верхней поверхности части бедра, прикрытой на полусогнутой ноге голенью |
Внутри и снаружи интенсивно желтого или оранжевого цвета |
Часто с желтыми пятнами (прежде всего в брачный период), почти всегда есть желтый цвет в рисунке |
В задней части беловатые, сероватые или реже зеленоватые пятна, не бывает желтых пятен |
|
Окраска резонаторов у ♂♂ |
Всегда белые, абсолютно непигментированные |
От белого до темно-серого (через все оттенки серого) |
От светло-серого до черного цвета |
|
Запах |
Слабый запах |
Сильный запах |
Резкий специфичный запах |
|
Брачная песня самца (после «запевки») |
Стрекочущая
|
Промежуточная по характеру между родительскими
|
Раскатистая, напоминающая хохот
|
|
Поведение при опасности |
Проплывают под водой и выныривают |
Промежуточное между родительскими видами |
Затаиваются на дне |
|
Поведение самцов относительно конкурентов |
Весьма агрессивное |
Агрессивное |
Относительно нейтральное |
|
Зимовка |
На суше |
Вместе с родительским видом |
В воде |
|
Предпочитаемые местообитания |
Небольшие водоемы лесной зоны, вне периода нереста живет на суше |
Различные, кроме крайних типов, характерных для родительских видов |
Крупные водоемы открытых ландшафтов |





К методам исследования на клеточном уровне относятся экспресс-метод определения плоидности путем измерения средней длины эритроцитов, исследования формы эритроцитов при сравнении ди- и триплоидов; определение уровня конденсации гетерохроматина. К методам исследования на уровне кариотипа относятся кариоанализ в раздавленных препаратах мацерированных тканей, кариоанализ раскапанных мацерированных клеток, определение плоидности с помощью подсчета ядрышек в интерфазном ядре, исследования маркеров хромосом типа ламповых щеток. Из молекулярных методов исследования использованы определение количества ядерной ДНК в соматических клетках, определение генотипов исследуемых животных и их половых клеток с видоспецифичными белковыми маркерами, определение количества гемоглобина и активности лактатдегидрогеназы.
РАСПРОСТРАНЕНИЕ ПОПУЛЯЦИОННЫХ СИСТЕМ PELOPHYLAX ESCULENTUS COMPLEX В БИОГЕОЦЕНОЗАХ ЛЕВОБЕРЕЖНОЙ ЛЕСОСТЕПИ УКРАИНЫ
Состав представителей Pelophylax esculentus complex, зарегистрированных в регионе исследования, показан на рис. 4 и в табл. 2, а расположение популяций и популяционных систем зеленых лягушек — на рис. 7 (см. далее).
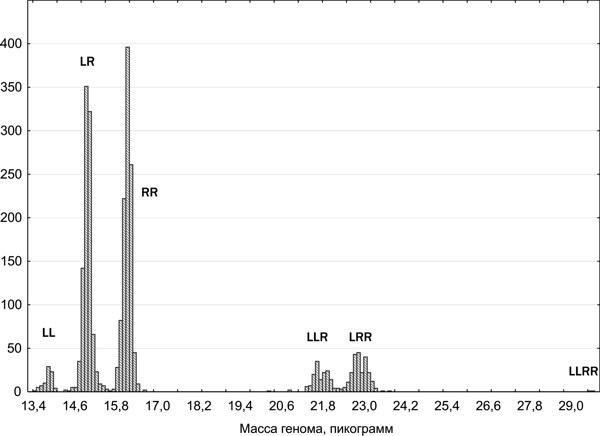
Рис. 4. Распределение зарегистрированных представителей Pelophylax esculentus complex по массе их геномов по результатам проточной ДНК-цитометрии
Таблица 2. Состав представителей Pelophylax esculentus complex, зарегистрированный в разных типах популяций и популяционных систем
|
Тип популяции или |
Состав форм, зарегистрированных в местообитании |
К-во |
% |
|
R-популяция |
RR |
50 |
37% |
|
R-E-тип |
RR, LR |
35 |
25,75% |
|
L-E-тип |
LL, LR |
1 |
0,75% |
|
L-E-R-тип |
LL, RR, LR |
10 |
7% |
|
R-E-Ep-тип |
RR, LR, LLR, LRR (в Верхнем Добрицком и Нижнем Добрицком — также LLRR) |
28 |
21% |
|
R-Ep-тип |
RR, LLR, LRR |
1 |
0,75% |
|
Ep-тип |
LLR, LRR |
1 |
0,75% |
|
R-E-Epf-тип |
RR, LR, ♀♀LLR (в Жовтневом — также ♀♀LRR) |
4 |
3% |
|
R-Epf-тип |
RR, ♀♀LLR |
4 |
3% |
Кроме родительских видов в регионе исследования зарегистрированы ди-, три- и, в редких случаях, тетраплоидные гибриды. Из точных способов определения лягушек, использованных в работе, наиболее массовым была проточная ДНК-цитометрия, которую выполняли Ю. М. Розанов и С. М. Литвинчук (ЦИН РАН, Санкт-Петербург). Этот метод базируется на том, что масса генома P. lessonae составляет 6,92 пг, а генома P. ridibundus — 8,07 пг (рис. 4). Определение ди- и триплоидов зеленых лягушек подтверждено различными методами кариоанализа. Состав представителей Pelophylax esculentus complex, распространенных на востоке Левобережно-Днепровской физико-географической провинции, соответствует тому, которой был обнаружен другими исследователями в ее западной части (Морозов-Леонов, 1998; Некрасова, Морозов-Леонов, 2001; Некрасова, 2002). На территории Среднерусской и Донецко-Донской северностепной физико-географических провинций обнаружены регионы массового распространения триплоидных P. esculentus.
БИОГЕОЦЕНОТИЧЕСКИЕ И ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ РАСПРОСТРАНЕНИЕ ПОПУЛЯЦИОННЫХ СИСТЕМ PELOPHYLAX ESCULENTUS COMPLEX В РЕГИОНЕ ИССЛЕДОВАНИЯ
Для определения факторов, влияющих на состав популяционных систем зеленых лягушек, применен канонический анализ (поиск связей между двумя списками переменных: «левым», касающимся наличия или отсутствия определенной формы в местообитании, и «правым» набором признаков местообитания). Биогеоценотические признаки местообитаний позволяют объяснить около 25% дисперсии по составу представителей зеленых лягушек. На рис. 5.А. показано размещение переменных, используемых в сокращенном варианте канонического анализа с биогеоценотическими признаками, на плоскости, заданной абсолютными (без учета знака) значениями канонических корней. Биогеоценотические признаки, расположенные на этой плоскости близко к переменным, показывающим наличие определенных форм лягушек, влияют на распространение этих форм.
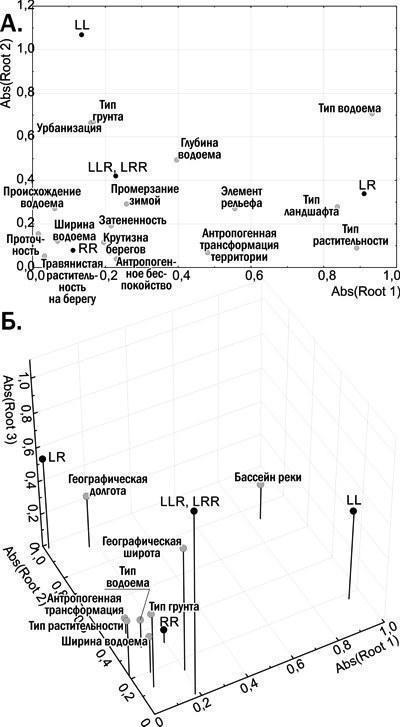
Рис. 5. Взаимосвязи переменных,использованных в каноническом анализе. А. Влияние сокращенного набора признаков биогеоценоза на распространение зеленых лягушек. Б. Совместное рассмотрение географических и биогеоценотических данных
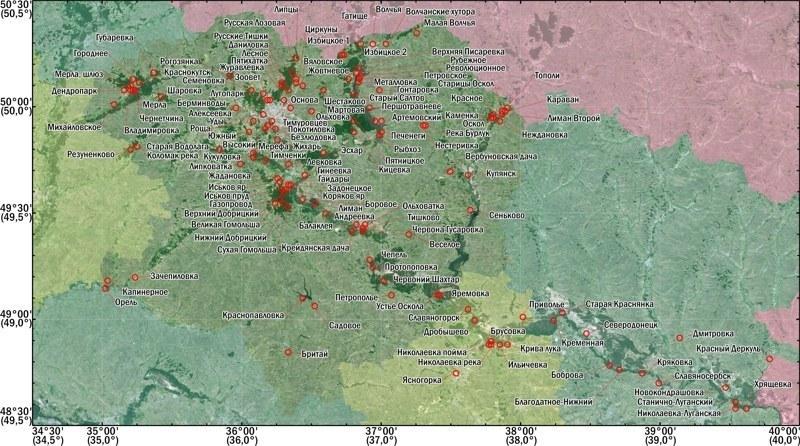
Рис. 6. Места сбора и условные названия изученных выборок
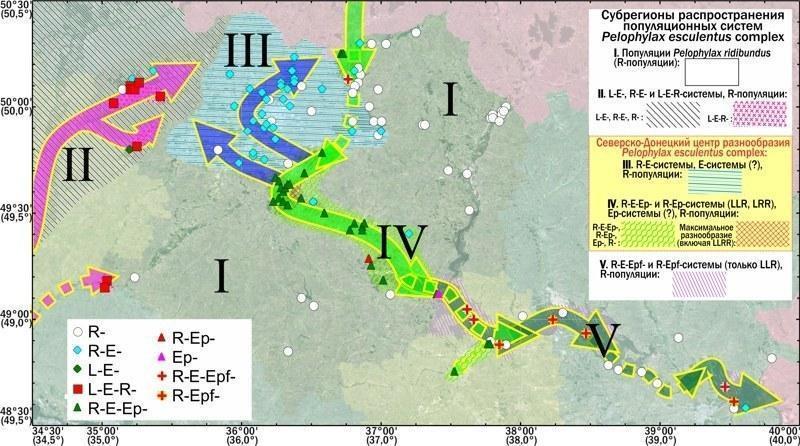
Рис. 7. Типы популяционных систем, субрегионы в регионе исследования, расположение Северско-Донецкого центра разнообразия Pelophylax esculentus complex и вероятные пути распространения различных форм зеленых лягушек
Добавление к анализу географических признаков (бассейн Днепра или Дона, географические координаты) значительно повышает долю дисперсии (составу представителей зеленых лягушек в местообитании), которую удается объяснить по результатам канонического анализа. В варианте, показанном на рис. 5.Б с помощью довольно короткого перечня признаков объясняется 41% дисперсии состава лягушек. Впрочем, в анализе, в котором сочетаются географические и биогеоценотические факторы, влияние географических оказывается более существенным. Так, на рис. 5.Б. все биогеоценотические факторы расположены у начала координат, рядом с переменной, обозначающей наличие в местообитании P. ridibundus, населяющего в регионе исследований почти любой водоем. Важность географических факторов в определении состава популяций и популяционных систем зеленых лягушек дает основание разделить регион исследования на пять субрегионов, отличающихся по составу характерных для них популяционных систем P. esculentus complex (рис. 7).
I субрегион образуют территории, на которых существуют только обычные видовые популяции P. ridibundus. Он охватывает другие субрегионы с южного и северо-восточного направлений. На большей части этого субрегиона нет местообитаний, отвечающих биогеоценотическим требованиям P. lessonae и P. esculentus. Но в этот субрегион входит также левый приток Северского Донца Оскол и его пойма (зона лесов, простирающаяся с севера на юг около 37º30' восточной долготы, см. рис. 6). В этом районе есть пригодные для P. esculentus местообитания, но, вероятно, во время расселения этих лягушек они не смогли попасть в Оскол и его пойму.
II субрегион расположен в бассейне Днепра и характеризуется R-E-L-, R-E-, LE-типами популяционных систем и популяциями P. ridibundus. Он ограничен наличием пригодных для P. esculentus и P. lessonae водоемов, расположенных рядом с участками лесов. В его состав входит изолированный (вероятно, реликтовый) анклав, расположенный в заказнике «Русский Орчик» на границе Харьковской, Полтавской и Днепропетровской областей. Следует подчеркнуть, что половозрелые P. lessonae найдены лишь в бассейне Днепра. В бассейне Северского Донца они отсутствуют.
III субрегион расположен в окрестностях г. Харькова в бассейнах рек Уды и Мжа. Для него характерны R-E-популяционные системы и популяции P. ridibundus. С востока III субрегион граничит с окрестностями Северского Донца, входящих в IV субрегиона, где кроме популяций P. ridibundus распространены R-Ep- и R-E-типы популяционных систем. В этом субрегионе имеются триплоидные P. esculentus обоих полов и обеих геномных композиций (LLR и LRR), и только в популяционной системе Жовтневое среди триплоидов утрачены самцы. IV субрегион тянется вдоль Северского Донца в Донецкой области.
В Донецкой и Луганской областях находится V субрегион, для которого характерны только P. ridibundus и самки триплоидов P. esculentus с композицией геномов LLR. В этом субрегионе остаются участки, характерные для IV субрегиона.
Распределение различных форм зеленых лягушек в регионе исследования позволяет предположить, какими были пути их расселения (рис. 7). Гипотеза, согласно которой триплоидные гибриды спускались по течению Северского Донца из IV субрегиона в V, постепенно теряя определенные формы, намного более вероятна, чем ее альтернатива, предполагающая их подъем против течения Донца. В ходе расселения P. esculentus не поднялись против течения Оскола, левого притока Северского Донца. Лягушки II субрегиона, расположенного в бассейне Днепра, имеют другое происхождение. III субрегион, вероятно, был заселен диплоидными гибридами, происходящими из IV субрегиона.
Чтобы установить биогеоценотические факторы, влияющие на распространение отдельных форм P. esculentus complex, следует анализировать их разнообразие внутри субрегионов, где распространена определенная эти формы. Обсуждаются факторы, влияющие на наличие диплоидных P. esculentus в III и IV субрегионах и триплоидных P. esculentus в IV субрегионе.
СЕВЕРСКО-ДОНЕЦКИЙ ЦЕНТР РАЗНООБРАЗИЯ PELOPHYLAX ESCULENTUS COMPLEX И ОСОБЕННОСТИ РАСПРОСТРАНЕННЫХ В НЕМ ЗЕЛЕНЫХ ЛЯГУШЕК
III и IV субрегион (см. Рис. 7) образуют зону, названную нами (Шабанов и др., 2009) Северско-Донецким центром разнообразия Pelophylax esculentus complex. Это соответствует международному определению понятия «центр разнообразия», который применяется для регионов, где в естественных условиях распространена большее количество определенных форм, чем на других территориях.
При изучении характера гаметогенеза P. esculentus из названного центра разнообразия были получены парадоксальные и, в определенной степени, противоречивые данные. В табл. 3 приведены результаты изучения сперматогенеза P. esculentus с помощью проточной ДНК-цитометрии, которую выполняли С. М. Литвинчук и Ю.М. Розанов (ЦИН РАН, г. Санкт-Петербург).
Таблица 3. Сперматогенез самцов P. esculentus с Северско-Донецкого центра разнообразия зеленых лягушек по данным проточной ДНК-цитометрии (rid — геном P. ridibundus, les — геном P. lessonae, rec — гаметы с рекомбинантным геномом, «-» — отсутствие гамет)
|
Тип популяционной системы |
Генотип |
К-во |
Типы гамет (% особей с опр. типом сперматогенеза) |
|||||
|
rid |
les |
rid + rec + les |
rid + les |
rec + les |
— |
|||
|
R-E- |
LR |
28 |
38% |
— |
4% |
29% |
|
29% |
|
R-E-Ep- |
LR |
99 |
25% |
9% |
1% |
33% |
1% |
31% |
|
LRR |
18 |
61% |
— |
— |
6% |
— |
33% |
|
|
LLR |
2 |
— |
100% |
— |
— |
— |
— |
|
Характер сперматогенеза определялся по размеру генома гаплоидных клеток. Суспензию спермы исследовали под микроскопом, чтобы проверить наличие диплоидных спермиев. Особей, которые производили диплоидные сперматозоиды, не найдено.
Кроме того, характер сперматогенеза P. esculentus с Северско-Донецкого центра разнообразия изучался путем электрофореза белковых маркеров (С.Ю. Морозов-Леонов, ИЗШ НАНУ). Его результаты показаны в табл. 4.
Таблица 4. Сперматогенез самцов P. esculentus из Северско-Донецкого центра разнообразия по данным электрофореза белковых маркеров (Наследование ..., 2009)
|
Генотип |
К-во |
rid |
les |
Нередуцированные гаметы (LR) |
rid + les |
|
LR |
10 |
1 |
5 |
1 |
3 |
|
LRR |
1 |
1 |
0 |
0 |
0 |
Результаты, приведенные в табл. 4, подтверждают наличие значительного количества особей с гибридной амфиспермией, но демонстрируют большее количество особей, продуцирующих гаметы les, чем данные табл. 3. Вероятно, эта разница отражает неоднородность групп исследуемых особей.
Характер оогенеза изучали с помощью электрофореза белковых маркеров и путем изучения хромосом типа ламповых щеток (Cytological maps ..., 2011). Большинство самок LR и LRR производит гаметы rid. Самки LLR продуцируют гаметы L. Зарегистрирована гибридная амфиспермия у самок LRR и продуцирования диплоидных яйцеклеток с двумя геномами P. ridibundus самками LR. Самки LR могут производить ооциты с генотипами LR, а самки LRR - ооциты с генотипом LRR. Такой характер гаметогенеза может объяснить появление триплоидия LRR, LLR и тетраплоидов.
Впрочем, имеющиеся данные о характере гаметогенеза с P. esculentus с Северско-Донецкого центра разнообразия оставляют больше вопросов, чем дают ответов. Поскольку в бассейне Северского Донца отсутствуют P. lessonae и распространены систем R-E- и R-E-Ep-типов, следует ожидать, что P. esculentus преимущественно продуцируют гаметы с геномом P. lessonae (рис. 3). Но у исследованных особей преобладает передача через гаметы генома P. ridibundus. Вызывает удивление большое количество особей с гибридной амфиспермией и особей с нарушенным сперматогенезом. Одним из путей исследования того, как воспроизводятся исследуемые нами популяционные системы, может быть имитационное моделирование.
Для экспресс-определения плоидности зеленых лягушек в наших исследованиях используется измерение параметров эритроцитов. В связи с этим мы провели исследование размера и формы эритроцитов различных P. esculentus (Бондарева и др., 2012). Установлено, что триплоидия приводит к увеличению эритроцитов и их ядер по всем параметрам, а ядерно-цитоплазматическое отношения остается постоянным. Относительная поверхность клеток у триплоидов уменьшается, что частично компенсируется их более эллиптической формой.
Нами были найдены две особи P. esculentus с аномально мелкими эритроцитами, которые по своим размерам должны были бы соответствовать гипотетическим гаплоидным клеткам. Удалось установить, что такие клетки являются диплоидными, а уменьшение ядер и клеток в целом является следствием более высокой степени конденсации гетерохроматина. Эритроциты триплоидов также отличаются от аналогичных клеток диплоидов более высокой степенью компактизации гетерохроматина.
Уменьшение относительной поверхности транспортных клеток должно влиять на особенности процессов обмена. Поэтому мы провели сравнение нескольких гематологических показателей ди- и триплоидных P. esculentus (Бондарев и др., 2013). Установлено, что у триплоидов уменьшено количество эритроцитов и содержание гемоглобина, а также повышены активность лактатдегидрогеназы в эритроцитах по сравнению с диплоидами. Последнее свидетельствует о том, что триплоидные лягушки имеют специфические механизмы компенсации увеличения размера эритроцитов, в частности интенсификации анаэробного расщепления глюкозы.
СПЕЦИФИКА БИОСИСТЕМ, В КОТОРЫХ ПРОИСХОДИТ ГЕМИКЛОНАЛЬНАЯ ГИБРИДИЗАЦИЯ И ВОСПРОИЗВОДСТВО ГЕМИКЛОНАЛЬНИХ ГИБРИДОВ
Популяционные системы, в которых воспроизводятся P. esculentus, принципиально отличаются от популяций видов с типичным половым размножением (рис. 2 Б.). Для них нами (Шабанов и др., 2009) предложено понятие гемиклональных популяционных систем (ГПС). Для ГПС характерно совместное проживание и воспроизводство особей родительского вида и различных форм гемиклональных гибридов, а также передача как клональных, так и, в типичном случае, рекомбинантных геномов.
Для ГПС Pelophylax esculentus complex присуща специфическая форма динамики, отличающаясяся и от демографических процессов, и от сукцессий — трансформации их состава, выражающиеся в изменении соотношения различных кариогенетических форм. Одним из способов изучения этой формы динамики является имитационное моделирование.
Отличия ГПС от обычных (менделевских) популяций является следствием отличий в характере популяционного воспроизводства. Для обобщения всех стадий процесса изменения форм популяционного воспроизводства, нами (Шабанов, 2010) предложена классификация его типов (табл. 5). Например, клональное воспроизводство может быть обеспечено благодаря бесполому размножению, гиногенезу или партеногенезу. Биосистемы, порождаемые различными способами воспроизводства, отличаются по своим свойствам (табл. 5).
Чтобы установить закономерности, определяющие состав ГПС зеленых лягушек и его динамику, необходимы долгосрочные наблюдения. Самую длительную историю изучения, начатую в 1995 году, имеет ГПС Иськова пруда, расположенного в окрестностях НПП «Гомольшанские леса» и Биостанции ХНУ имени В. Н. Каразина. В 1995 г. в пруду была зарегистрирована жизнеспособная популяционная система, состоящая из диплоидов P. esculentus (Лада, 1998). После осушения в 2000 г. пруд был вновь частично заполнен в 2001 г. (Ачкасова и др., 2001). Впоследствии его ГПС оказалась в критическом состоянии. По нашим предположениям, в 2006 г. в пруду большинство P. esculentus были носителями мужского клонального генома P. lessonae. Во время нереста там находились почти исключительно самцы; с ними скрещивались единичные самки P. ridibundus, которые могли подходить из соседних местообитаний (Шабанов и др., 2006). В результате появлялись снова исключительно самцы P. esculentus: ♀XRXR × ♂XR(YL) → ♂XR(YL). Имитационное моделирование (Кравченко, 2013) показало, что для выживания ГПС необходимо попадание в нее других клональных геномов.
Таблица 5. Классификация типов популяционного воспроизводства и связанные с ними особенности биосистем (разнообразие форм генетической рекомбинации, показанное в двух последних сроках таблицы, рассматривается в последнем разделе работы)
|
Биосистемы и их особенности |
Популяции |
ГПС |
||||
|
клональные |
менделевские |
|||||
|
с горизонтальным переносом |
без переноса |
гермафродитные |
раздельнополые |
|||
|
без полового диморфизма |
с половым диморфизмом |
|||||
|
Ia |
Ib |
II |
IIIa |
IIIb |
IV |
|
|
Особи |
Однотипны |
Однотипны, два пола |
Два ярко отличающихся пола |
Два пола, разные формы |
||
|
Генеалогия |
Линейная |
Сетчатая |
Фактически линейная |
|||
|
Уникальность особи |
Нет |
Есть |
Ненаследуемая (гемиклоны и клоны) |
|||
|
Генофонд популяции |
Нет |
Есть |
Нет (набор гемиклонов и клонов) |
|||
|
«Двойная цена пола» |
Нет |
Есть |
||||
|
Половой отбор |
Нет |
Малоэффективный |
Эффективный |
Есть |
||
|
Внутривидовая рекомбинация |
Негомологичная (в меньшей степени — гомологичная) |
Нет |
Гомологичная |
Нет или ограниченная гомологичная (у триплоидов) |
||
|
Межвидовая рекомбинация |
Негомологичная |
Нет |
Гомеологичная |
|||
В 2010-2011 гг. в описываемой ГПС появились многочисленные одно- и двухлетние лягушки. С 2010 мы начали ее исследования методом мечения и повторного отлова (Популяционная ..., 2012). Состав этой ГПС продолжает меняться. В 2012-2013 гг. количество половозрелых особей в составе нерестового стада в этой популяционной системе составляло около 500 особей, а в 2014 г. оно значительно снизилась. Зарегистрирован значимый рост доли триплоидных P. esculentus (p <0,02) и уменьшение доли самок (составляющих около 20% нерестового стада). Вероятно, ГПС Иськова пруда не нашла стабильное состояние и вновь оказалась в кризисе (Meleshko et al, 2014). Наблюдение за ее будущим представляют исключительный интерес.
С 2012 начато мечение еще в двух ГПС: Нижний Добрицкий и Жовтневое (рис. 6). В ГПС зеленых лягушек, населяющих пруд в с. Жовтневое, зарегистрирована система R-E-Epf-типа, в которой триплоиды представлены самками LRR и LLR. Результаты экспресс-анализа плоидности с помощью измерения размера эритроцитов соответствуют результатам проточной ДНК-цитометрии. Эта ГПС расположена на границе IV и III субрегионов и демонстрирует возможность частичной потери геномов, приводящих к появлению триплоидов.
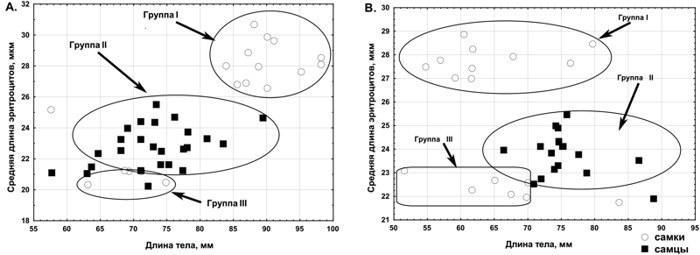
Рис. 8. Зависимость длины эритроцитов от длины тела зеленых лягушек из пруда с. Жовтневое. А. — выборка 2012 г; В. — выборка 2013. Группа I — триплоидные самки, группа II — диплоидные самцы, группа III — диплоидные самки
ПОПУЛЯЦИОННО-ЭКОЛОГИЧЕСКИЕ ПАРАМЕТРЫ ПОПУЛЯЦИОННЫХ СИСТЕМ PELOPHYLAX ESCULENTUS COMPLEX, СУЩЕСТВЕННЫЕ ДЛЯ ИМИТАЦИОННОГО МОДЕЛИРОВАНИЯ ИХ ТРАНСФОРМАЦИЙ
Рассмотрены особенности годового цикла зеленых лягушек и их онтогенеза, которые необходимо учитывать при имитационном моделировании. Перечислены параметры жизнеспособности, которыми охарактеризована жизнеспособность различных кариогенетических форм. Смертность лягушек разделена на зависимую от плотности популяции (связанную с конкуренцией) и независимую от плотности.
Описаны упрощения, принятые при построении имитационной модели. Приведены разностные уравнения, использованные при разработке концептуальной модели. Проведено сравнение различных реализаций имитационной модели: предыдущей, созданная в Microsoft Excel (Кравченко, Шабанов, 2010; Исследование…, 2011; Кравченко, 2013), и примененной в этой работе, консольного приложения для Java, разработанного А. О. Леоновым. Логика работы модели условно показана на рис. 9.
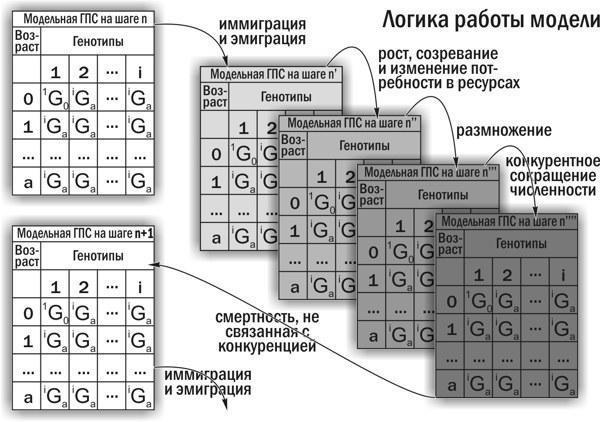
Рис. 9. Пояснение логики работы имитационной модели
В модели принято, что потребление ресурсов пропорционально биомассе особи; динамику биомассы оценили с применением скелетохронологии. Приведены объяснения выбора параметров, используемых при моделировании. Охарактеризован равновесный состав модельной популяции P. ridibundus, порождаемой при принятых умолчаниях.
Рассмотрены примеры динамики модельной ГПС: при попадании в популяцию родительского вида (P. ridibundus) женского гетероспецифичного генома (рис. 10), а также женского конспецифичного генома, который добавлялся в ГПС после увеличения численности носителей гетероспецифичного генома (рис. 11).
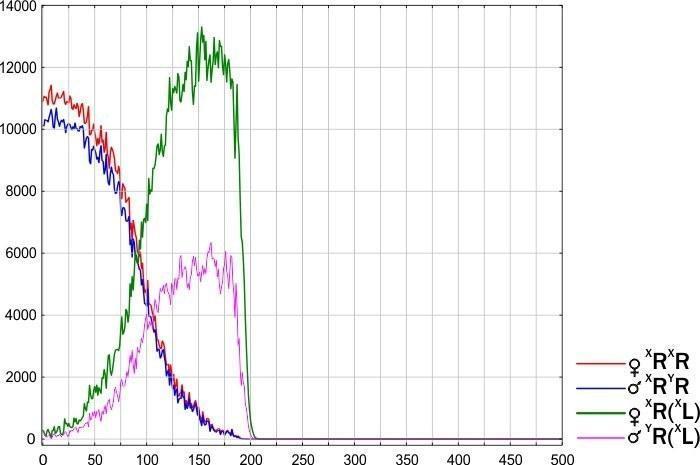
Рис. 10. Гибель ГПС R-E-типа, в которой передаются гетероспецифичные клональные геномы, (XL). Этот процесс начался с того, что на первом шаге имитации в популяцию P. ridibundus попала одна самка P. esculentus XR(XL)
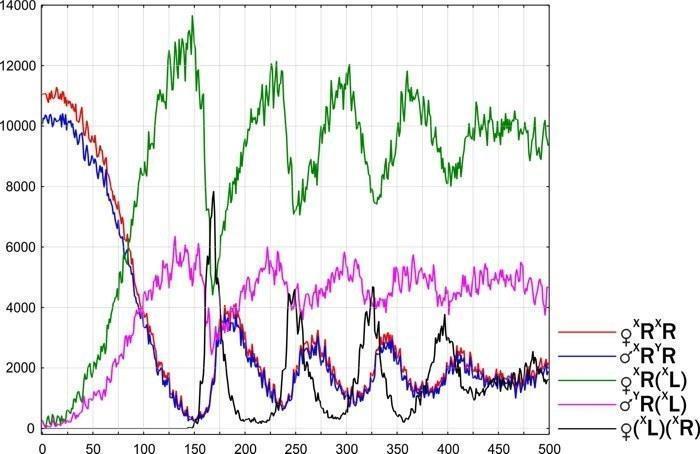
Рис. 11. Переход к стабильному сосотоянию ГПС R-E-типа, в которой передаются геномы (XR) и (XL). До 144-го шага динамика показаной тут ГПС идентична предыдущему случаю. На 145-м шаге в ГПС добавляется одна самка P. esculentus (XL)(XR), что приводит к переходу ГПС в стабильное состояние
Особенностью примененной модели является то, что результат ее работы является вероятностным. Выживание и размножение каждой особи — случайные процессы, вероятность которых определяется параметрами жизнеспособности каждой кариогенетической формы и каждой возрастной группы. Для каждого варианта начальных условий эксперимента можно определить определенное распределение вероятностей конечных состояний имитации.
На рис. 12 показано распределение вероятностей в случае имитаций, построенных следующим образом. На первом этапе модели в ГПС добавлялось 10 самок XR(XL). В большинстве случаев в этой ГПС начинался процесс, показанный на рис. 10. После этого в ГПС добавлялась одна самка XL(XR). Время, когда она прилагалась, менялось от 1-го до 300-го шага модели; для каждого варианта срока добавления самки XL(XR) было выполнено по 200 имитаций.
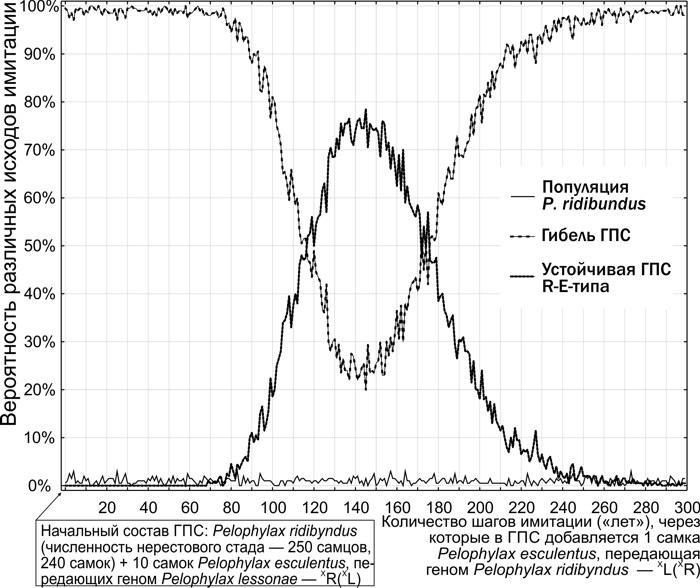
Рис. 12. Зависимость вероятности перехода к стабильному состоянию ГПС R-E-типа, в которой наростает доля геномів (XL), от времени попадания в нее геномов (XR)
В описанном эксперименте были возможны три конечные состояния:
— в модельной ГПС исчезает геном (XL) и она становится популяцией P. ridibundus;
— модельная ГПС погибает, как на рис. 10;
— модельная ГПС переходит в стабильное состояние, как на рис. 11.
В ходе экспериментов с моделью зарегистрированы неустойчивые состояния модельной ГПС, которые со временем под влиянием случайных причин переходят к другим состояниям. Пример такого перехода от нестабильного к стабильному состоянию показано на рис. 13.
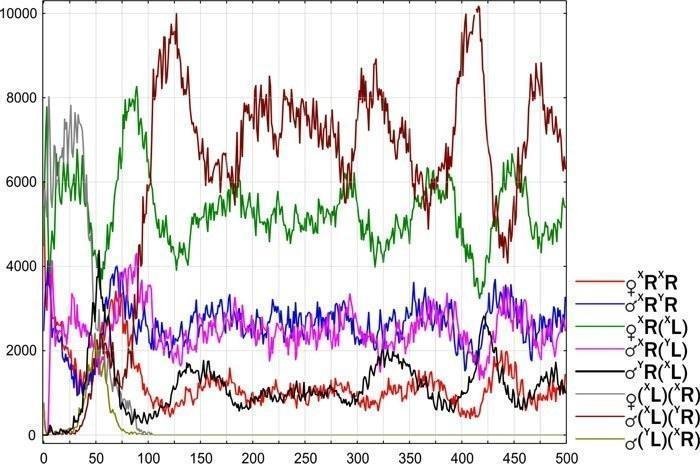
Рис. 13. Модельная ГПС пребывала в нестабильном состоянии (который в следующей главе обозначено как Wandering R-E-type). После случайной потери генома (XR) и, соответственно, с исчезновением генотипов (XL)(XR) і (YL)(XR), ГПС перешла в устойчивое состояние
Обсуждается возможность проверки адекватности результатов моделирования. С помощью моделирования невозможно доказать, что процессы в реальных ГПС происходят именно определенным образом. Но в случае регистрации существенных расхождений результатов моделирования и эмпирических данных можно отклонить предположения, проверяемые при моделировании. Для сравнения можно использовать распределение состояний, которые принимают модельные ГПС, и эмпирически зарегистрированное разнообразие природных ГПС (рис. 14).
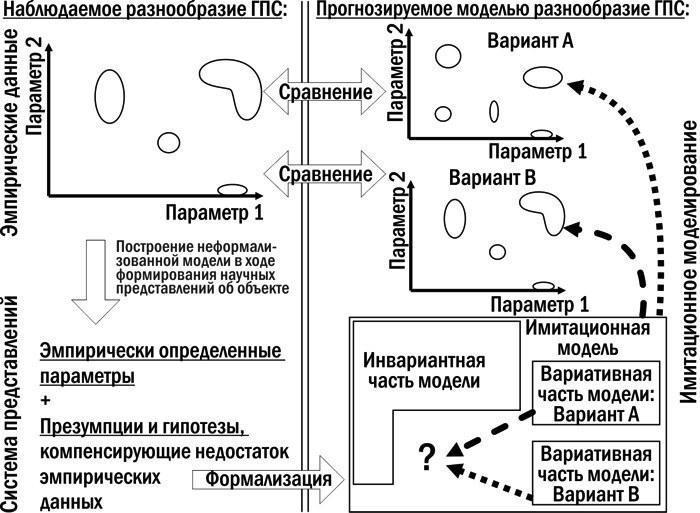
Рис. 14. Использование имитационной модели для проверки гипотез о механизмах функционирования ГПС. Распределение модельных ГПС, которое соответствует эмпирическому, порождается при выборе варианта В вариативной части. Это не доказывает вариант В, но позволяет отбросить вариант А в его пользу
ДИНАМИЧЕСКАЯ ТИПОЛОГИЯ И УСТОЙЧИВОСТЬ ПОПУЛЯЦИОННЫХ СИСТЕМ, ВКЛЮЧАЮЩИХ ДИПЛОИДНЫХ ПРЕДСТАВИТЕЛЕЙ PELOPHYLAX ESCULENTUS COMPLEX
Проведено сравнение трех типов типологий. Ассоциативные типологии связывают объекты с определенными образцами. Аналитические классифицируют их по имеющимся признакам. Динамические распределяют объекты на типы по прогнозам их будущей динамики.
Для построения динамической типологии ГПС зеленых лягушек, состоящих из диплоидных представителей, перебраны 5 895 начальных составов ГПС, равномерно распределенных в пространстве их возможных состояний. Вариативная часть предположений при моделировании (рис. 14) соответствует результатам изучения R-E-ГПС Северско-Донецкого центра разнообразия. Для 5 895 начальных точек проведено по 10 имитаций из 500 шагов, а затем для тех 1 117 точек, имитации для которых закончились различными финалами, — еще по 10 имитаций. Полученную совокупность из 70 120 финалов распределили на 5 груп (рис. 15):
— Death — 37 946 финалов, закончившихся гибелью ГПС;
— R-E-HPS — 22 204 финала, соответствующих различным ГПС R-E-типа;
— HPS E-type I — 5 884 финала, соответствующих «чистым» ГПС E-типа; в состав таких ГПС входят ♀(XL)(XR) и ♂(XL)(YR);
— HPS E-type II — 3 892 финала, соответствующих иному возможному типу «чистых» ГПС E-типа, в состав которого входят ♀(XL)(XR) и ♂(YL)(XR);
— Dying — 124 финалы в окрестностях точки Dead, движущихся к гибели (эта группа по своему составу может быть разделена еще на 13 типов);
— R-population - 70 финалов, соответствующие популяции P. ridibundus.
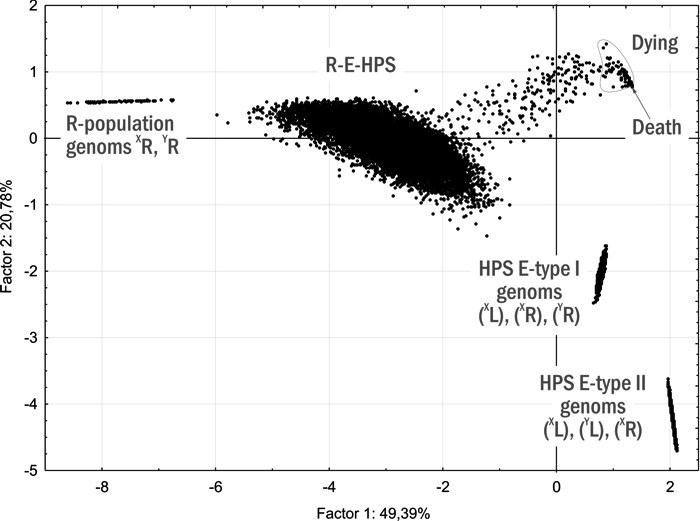
Рис. 15. Ординация на плоскости двух первых главных компонент результатов 70 120 имитаций. Анализируются данные о составе нерестового стада модельной ГПС после 500 шагов моделирования. Начало координат является центром равновесия для размещенных вокруг него точек; асимметричность графика связана с тем, что в правом верхнем квадранте находится точка Dead, которой соответствуют 56% всех имитаций
В составе группы R-E-HPS выделены (рис. 16) три пересекающиеся подгруппы:
— Stable RE-type I — 18 852 исхода, соответствующие стабильному состоянию на рис. 11.
— Stable RE-type II — 2 847 финалов, соответствующие стабильному состоянию на рис. 13;
— Wandering RE-type — 505 случаев, в которых, кроме P. ridibundus, присутствуют P. esculentus, передающие оба гетероспецифичных генома и женский конспецифичный геном.
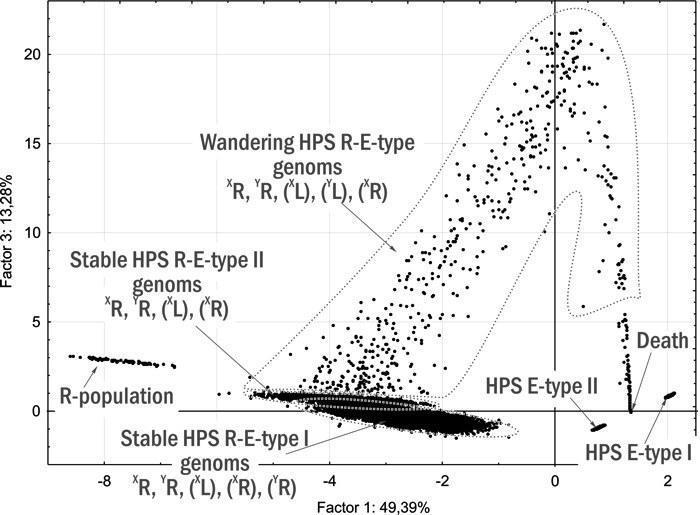
Рис. 16. Ординация на плоскости І и ІІІ главных компонент финалов тех же имитаций, что и на рис. 15
Чтобы определить, к каким типам по характеру их устойчивости принадлежат выделенные динамические типы ГПС, проанализированы 162 580 пар из состояний, в которых находилась модельная ГПС на определенном этапе, и состояний, в которые она перешла через 100 шагов имитации. Все зарегистрированные состояния ГПС были классифицированы по типам устойчивости, показанным на рис. 17).
Таким образом, в пространстве возможных состояний ГПС, состоящих из диплоидных особей и соответствующих по характеру передачи геномов особенностям Северско-Донецкого центра разнообразия Pelophylax esculentus complex, существует шесть бассейнов устойчивости. Один из них соответствует популяции родительского вида, два — ГПС, состоящим исключительно из P. esculentus, два — ГПС, содержащим P. ridibundus и P. esculentus и еще один — гибели ГПС.
Проведено сравнение прогнозов, полученных в результате моделирования, и имеющихся данным о составе естественных ГПС и характера гаметогенеза P. esculentus. Показано, что в настоящее время в регионе исследования отсутствуют ГПС E-типа, но, вероятно, две такие системы существовали в 1995 году (Лада, 1998). Вариант Stable RE-type II не соответствует имеющимся данным. Предположение, что в регионе возможно существование Stable RE-type I, а большинство зарегистрированных систем находятся в переходных состояниях, не противоречит эмпирическим данным.
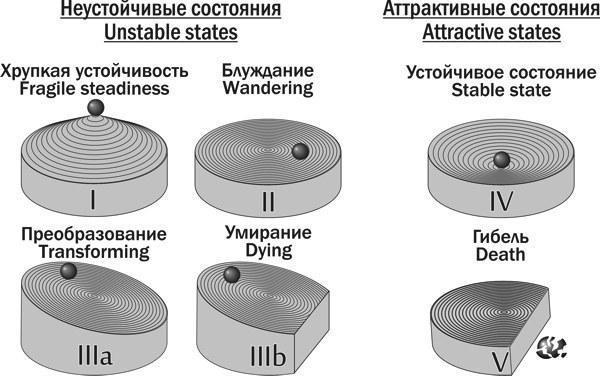
Рис. 17. Типы устойчивости биосистем, зарегистрированные в экспериментах с имитационной моделью
МНОГОУРОВНЕВЫЙ ОТБОР В PELOPHYLAX ESCULENTUS COMPLEX
Зеленые лягушки — удачная модель для изучения многоуровневого отбора (табл. 6; Шабанов, 2012).
Геномный уровень отбора. Нежизнеспособность гибридолизного потомства принято объяснять накоплением летальных мутаций в клональных геномах вследствие действия «храповика Мёллера» (Plötner, 2005). Этому противоречит тот факт, что жизнеспособность потомков, которые получили клональные геномы одного вида, снижается в равной степени, копии это одного клона или разных (Plötner, 2005). Это противоречие снимает выдвинутая нами гипотеза о подборе клональных геномов на способность обеспечивать элиминацию альтернативного генома (Шабанов, 2006). Ее косвенным подтверждением является широкое распространение гибридной амфиспермии в Северско-Донецком центре разнообразия зеленых лягушек. Согласно этой гипотезе причина гибридной амфиспермии — конкуренция двух геномов, измененных длительной клональной передачей в линиях зародышевых клеток.
Таблица 6. Уровни многоуровневого отбора в Pelophylax esculentus complex
|
Уровень |
Примеры вызванных процессов |
Зарегистрированные в работе проявления |
|
Генный уровень: «эгоистичный ген» |
Изменение частот аллелей вследствие отбора их фенотипических проявлений. |
В данной работе не рассматривалось, кроме регистрации отличия региональных групп популяций по частотам аллелей |
|
Геномный уровень: «эгоистичный геном» |
Целостная передача одного генома при гемиклональном наследовании как предельный случай генного драйва; конкуренция клональных геномов в одном гаметоците |
Феномен гемиклонального наследования. Выдвинута гипотеза, объясняющая гибридную амфиспермию конкуренцией клональных геномов, измененных отбором на способность элиминировать альтернативный геном) |
|
Клеточный уровень: «эгоистичная клеточная линия» |
Отбор клеток зародышевой линии, приводящий к формированию жизнеспособных гамет с определенным клональным геномом. |
Наличие механизма «фильтрации» результатов неправильного сперматогенеза. Различная устойчивость гаметогенеза у разных форм |
|
Организменный уровень: «эгоистичный индивид» |
Конкуренция гемиклонов и клонов по их жизнеспособности и способности к воспроизводству. |
Различия особей по развитию гонад и фертильности. Различия особей по жизнеспособности. Изменение соотношения разных форм в разных генерациях. Различные стратегии развития |
|
Групповой уровень: «эгоистичная ГПС» |
Различия ГПС по их устойчивости. Расселение успешных ГПС, гибель или преобразование неустойчивых |
В экспериментах с моделью показана различная устойчивость разных типов, что соответствует наблюдениям за ГПС |
Клеточный уровень отбора. Установлено, что гаметогенез у P. esculentus значимо менее устойчив, чем у P. ridibundus. В семенниках самцов P. esculentus найдены клетки с кариотипом, отсутствующим в сперме (Михайлова и др., 2011; Кечеджи и др., 2011). Это свидетельствует о наличии механизма элиминации клеточных линий, развитие которых не привело к формированию полноценных гамет.
Организменный уровень отбора. Нарушение сперматогенеза у гибридов первого поколения известны давно (Berger, 1971), но зарегистрированная нами степень распространенности таких нарушений оказалась неожиданным (Нарушение .., 2014). Нарушения фертильности обнаружены у 85% P. esculentus (n = 46) и 29% P. ridibundus (n = 17) из Северско-Донецкого центра разнообразия зеленых лягушек.
Скелетохронологические исследования демонстрируют различия особей по скорости роста, времени наступления половой зрелости и продолжительности жизни. В разнообразии внутрипопуляционных онтогенетических стратегий зеленых лягушек (и ряда других групп) можно предположить существование двух типичных случаев, скороспелости и тугорослости (Внутрипопуляционные ..., 2014).
Групповой уровень отбора. Неодинаковая устойчивость разных типов ГПС — результат моделирования, соответствующий наблюдениям. Различные типы ГПС погибают, изменяются или находятся в устойчивом состоянии. Они могут «размножаться» (расселяться), передавая определенный набор клональных геномов.
Филогенетический уровень отбора. Известно три пути преодоления гибридной стерильности, позволяющие сохранить гибридную композицию геномов (Шабанов, 2006): переход к клональному воспроизводству (от вегетативного размножения до партеногенеза); полиплоидия; гемиклональность. Из них именно гемиклональное наследование требует действия сложного кариогенетического механизма, обеспечивающего элиминацию и эндоредупликацию геномов. Поскольку причиной целесообразности биосистем является отбор, необходимо установить, отбор какого уровня поддерживает гемиклональное наследование.
В ряде работ показано, что в результате нарушений гемиклональности при воспроизводстве гибридов возникают частичные рекомбинанты, которые при скрещивании с представителями родительских видов могут переносить в их генофонды фрагменты генетической информации другого родительского вида (Наследование ... 2009; The ploidy ... 2010 и др. работы). Это проявления гомеологической рекомбинации (табл. 5), то есть рекомбинации между соответствующими хромосомами, принадлежащих разным видам. Нами высказано предположение, что ограниченная (уровнем нарушений при передаче клонального генома) межвидовая гомеологичная рекомбинация дает эволюционное преимущество тем видам, скрещивание которых приводит к появлению гемиклональных гибридов. Это объяснение соответствует концепции отбора видов (Eldredge, Gould, 1972), который был доказан в ряде случаях (Goldberg et al, 2010). Мы предполагаем, что способность к скрещиванию с образованием гемиклональных гибридов в роде Pelophylax поддерживается отбором видов. Этим объясняется быстрая эволюция рода и наличие в нем нескольких гибридогенних комплексов (Plötner, 2005).
ЗАКЛЮЧЕНИЕ
Обсуждаются перспективы дальнейших исследований ГПС зеленых лягушек Северско-Донецкого центра разнообразия Pelophylax esculentus complex.
ВЫВОДЫ
1. На территории Левобережной лесостепи Украины представители гибридогенного комплекса Pelophylax esculentus существуют не только в составе популяций родительских видов Pelophylax lessonae (Camerano, 1882) и Pelophylax ridibundus (Pallas, 1771), но и в составе популяционных систем, включающих диплоидных и, в ряде случаев, полиплоидных межвидовых гибридов Pelophylax esculentus (Linnaeus, 1758). На состав и динамику таких систем влияют биогеоценотические особенности местообитаний, специфика гемиклонального воспроизводства Pelophylax esculentus и ограниченное распространение отдельных форм Pelophylax esculentus в речных бассейнах в регионе исследования.
2. В регионе исследования выделены пять субрегионов, отличающихся по составу распространенных в них популяционных систем и популяций представителей Pelophylax esculentus complex. Популяционные системы, включающие Pelophylax esculentus, встречаются в четырех из этих субрегионов, триплоиды Pelophylax esculentus зарегистрированы в двух из них.
3. Внутри выделенных субрегионов состав популяционных систем Pelophylax esculentus complex определяется биогеоценотическими условиями, в первую очередь — характером растительности в окрестностях нерестового водоема и уровнем антропогенной трансформации территории. Различия субрегионов связаны частично с различием характерных для них биогеоценотических условий, а частично — с изоляцией бассейнов разных рек.
4. Для двух субрегионов, расположенных в водосборном бассейне реки Северский Донец, зарегистрировано уникально высокое разнообразие представителей Pelophylax esculentus complex, а также необычный, по сравнению с окружающими территориями, состав их популяционных систем. Этот регион назван Северско-Донецким центром разнообразия Pelophylax esculentus complex.
5. Уникальность Северско-Донецкого центра разнообразия Pelophylax esculentus complex связана со следующими его особенностями:
— на части его территории широко распространены триплоидные Pelophylax esculentus обоих полов с генотипами обеих возможных геномных композиций (LLR и LRR);
— в нем преобладают ГПС R-Ep- и R-E-типа;
— в нем отсутствуют половозрелые Pelophylax lessonae; особи Pelophylax lessonae, возникающие в результате гибридолиза (скрещивания гибридов), гибнут на начальных етапах онтогенеза;
— его характеризует высокая доля особей Pelophylax esculentus с гибридной амфиспермией (производством половых клеток обоих родительских видов);
— значительная часть особей Pelophylax esculentus демонстрирует нарушения фертильности.
6. Характерные свойства биосистем различных уровней организации тесно связаны со способом рекомбинации (на генном уровне), гаметогенеза или сравнимых процессов (на клеточном уровне), размножения (на организменном уровне) и популяционного воспроизводства. Гемиклональное наследование у Pelophylax esculentus связано с глубоким изменением всей цепочки этих процессов и порождает специфичные биосистемы разных уровней организации, отличающиеся от таковых у организмов с типичным половым размножением.
7. Существование межвидовых гибридов Pelophylax esculentus, в силу особенностей их воспроизводства, возможно не в составе популяций, а в составе гемиклональных популяционных систем (ГПС). ГПС гибридогенных комплексов видов являются особой категорией биосистем, принципиально отличающейся от популяций (тем, что состоят из особей различных видов и межвидовых гибридов, а также тем, что в них передаются не только рекомбинантные, но и клональные геномы), а также от гильдий и сообществ (тем, что объединены совместным воспроизводством). ГПС характеризуются не только половой и возрастной структурой, но и составом кариогенетических форм (групп особей, которые отличаются по геномной композиции и составу геномов, передаваемым в гаметах), который определяет состав клонально передаваемых геномов. ГПС способны к трансформациям, т.е. изменению состава и соотношения различных кариогенетических форм в их составе.
8. Для исследований механизма трансформаций ГПС Pelophylax esculentus complex разработана и создана имитационная модель. Эта модель дает возможность получать распределения вероятностей исходов трансформаций ГПС в зависимости от исходного состава ГПС, а также принятых предположений о характере наследования различных геномов и относительной жизнеспособности разных кариогенетических форм Pelophylax esculentus complex в условиях определенного биогеоценоза.
9. Использование имитационной модели, созданной в работе, позволяет тестировать гипотезы, относящиеся к неизученным аспектам функционирования ГПС, на соответствие комплексу накопленных эмпирических данных.
10. Разработана динамическая (объединяющая рассматриваемые объекты в группы на основе прогноза их будущей динамики) типология ГПС, включающих диплоидных представителей Pelophylax esculentus complex. Показано, что такие ГПС могут существовать всего в четырех устойчивых состояниях, в которых в их состав входят представители Pelophylax esculentus. Во всех этих четырех устойчивых состояниях в ГПС передаются геномы обоих родительских видов.
11. Обоснована теоретическая возможность существования гемиклональных популяционных систем, состоящих исключительно из диплоидных Pelophylax esculentus. Показано, что эти ГПС будут устойчивыми в том случае, если будут состоять из особей с гибридной амфиспермией, относящихся к двум разным клонам (совокупностям генетически идентичных особей).
12. Pelophylax esculentus complex предоставляет исключительные возможности для изучения многоуровневого отбора. В нем зарегистрирован отбор на нескольких уровнях:
— генный, соответствующий модели «эгоистичного гена» и проявляющийся в различной успешности в распространении в популяциях или ГПС отдельных аллелей;
— геномный (предельный случай генного в том случае, если все гены одного генома наследуются как единое целое), соответствующий модели «эгоистичного генома» и проявляющийся в конкуренции отдельных гемиклонов;
— клеточный, соответствующий модели «эгоистичной линии зародышевых клеток» и проявляющийся в конкуренции различных линий зародышевых клеток в половых железах Pelophylax esculentus; образованием жизнеспособных гамет заканчивается развитие только тех линий, в которых успешно происходит элиминация рекомбинантного генома и эндоредупликация клонального (в случае диплоидных гибридов);
— организменный, соответствующий модели «эгоистичного организма» и проявляющийся в чистом виде в конкуренции гибридных особей внутри клона;
— групповой, соответствующей модели «устойчивой ГПС» и проявляющийся в различии вероятности выживания и расселения ГПС различного состава.
Динамика изменений каждой отдельной ГПС определяется взаимодействием эффектов отбора на разных уровнях.
13. Выдвинута гипотеза, объясняющая феномен гибридной амфиспермии конкуренцией в клетках зародышевого пути клональных геномов, измененных вследствие отбора на способность вызывать элиминацию альтернативных геномов
14. Выдвинута гипотеза, согласно которой способность двух относительно близких видов к гемиклональной гибридизации является приспособлением к гомеологичной межвидовой рекомбинации, которое поддерживается на уровне отбора видов.
БЛАГОДАРНОСТИ
Автор выполнял данную работу в сотрудничестве со многими коллегами и получил ценную консультационную и организационную помощь от многочисленных специалистов. На разных этапах научного пути его направляли В. П. Кудокоцев та В. М. Эпштейн, благодарную память о которых хранит диссертант. Автор исполнен признательности к В. Л. Булахову, который был руководителем его кандидатской диссертации, и А. П. Расницыну, предоставившему ценные консультации во время подготовки докторской. Важную роль в выполнении этой работы сыграло сотрудничество з С. Н. Литвинчуком, Ю. М. Розановым, Л. Я. Боркиным, Г. А. Ладой и С. Ю. Морозовым-Леоновым. Особо важной для автора была работа А. В. Коршунова и М. А. Кравченко, которые подготовили свои кандидатские диссертации под его руководством. Всем названным и многим неназванным коллегам автор висказывает искреннюю и глубокую благодарность.
ПУБЛІКАЦИИ ПО ТЕМЕ ДИССЕРТАЦИИ
Статьи в изданиях, которые входят в наукометричные базы данных
Наследование родительских геномов гибридной формой Rana “esculenta” (Amphibia, Ranidae) / С. Ю. Морозов-Леонов, С. В. Межжерин, О. Д. Некрасова, Д. А. Шабанов, А. В. Коршунов, Ф. Ф. Куртяк // Генетика. – 2009. – Том 45, № 4. – С. 488–495. (Диссертант принимал участие в сборе материала и интерпретации результатов).
The ploidy and genetic structure of hybrid population of water frogs Pelophylax esculentus complex (Amphibia, Ranidae) of Ukraine fauna / S. V. Mezhzherin, S. Yu. Morozov-Leonov, O. V. Rostovskaya, D. A. Shabanov, L. Yu. Sobolenko // Cytology and Genetics. – 2010. – Vol. 44, № 4. – P. 212–216. (Диссертант принимал участие в планировании сборов в Левобережной лесостепи Украины, собирал материал и участвовал в интерпретации результатов).
Influence of environmental conditions on the distribution of Central Asian green toads with three ploidy levels / S. N. Litvinchuk, G. O. Mazepa, R. A. Pasynkova, A. Saidov, T. Satorov, Yu. A. Chikin, D. A. Shabanov, A. Crottini, L. J. Borkin, J. M. Rosanov, M. Stöck // Journal of Zoological Systematics and Evolutionary Research. – 2011. – Vol. 49, № 3. – P. 233–239. (Диссертант принимал участие в сборе материала и интерпретации результатов).
Кечеджи А. Е. Сперматогенез у триплоидных Pelophylax esculentus (Amphibia, Anura) из Северско-Донецкого центра разнообразия зеленых лягушек (Украина, Харьковская область) / А. Е. Кечеджи, О. В. Михайлова, Д. А. Шабанов // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2011. – Вип. 14, № 971. – С. 112–116. (Диссертант принимал участие в постановке задания для исследования, выборе и отработке методики, проведении полевых работ, а также в интерпретации результатов).
Бондарева А. А. Цитогенетические особенности эритроцитов зеленых лягушек из Северско-Донецкого центра разнообразия Pelophylax esculentus complex / А. А. Бондарева, Ю. С. Бибик, С. М. Самило, Д. А. Шабанов // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2012. – Вип. 15, № 1008. – С. 116–123. (Диссертант принимал участие в постановке задания для исследования, выборе и отработке методики, проведении полевых работ и в интерпретации результатов).
Cytological maps of lampbrush chromosomes of European water frogs (Pelophylax esculentus complex) from the Eastern Ukraine / D. Dedukh, G. Mazepa, D. Shabanov, Ju. Rosanov, S. Litvinchuk, L. Borkin, A. Saifitdinova, A. Krasikova // BMC Genetics. – 2013. – Vol. 14. – P. 26–46. (Диссертант принимал участие в сборе материала и в интерпретации результатов).
Внутрипопуляционные онтогенетические стратегии скороспелости и тугорослости: определение на примере бесхвостых амфибий / Д. А. Шабанов, А. В. Коршунов, Е. В. Мелешко, А. В. Шабанова, Е. Е. Усова // Вестник Харьковского национального университета имени В. Н. Каразина, серия "Биология". – 2014. – Вып. 22, № 1126. – С. 115 – 124 (Диссертант принимал участие в постановке задания для исследования, анализе накопленного материала и в интерпретации результатов).
Meleshko O. V. The study of three hemiclonal population systems Pelophylax esculentus complex from the Seversko-Donetskiy center of green frogs diversity / O. V. Meleshko, O. V. Korshunov, D. A. Shabanov // Вісник Харківського національного університету імені В. Н. Каразіна. Серія «Біологія». – 2014. – Вип. 20, № 1100. – С. 153–158. (Диссертант принимал участие в постановке задания для исследования, выборе и отработке методики, проведении полевых работ и в интерпретации результатов).
Статьи в профильных изданиях Украины и других стран
Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine / L. J. Borkin, A. V. Korshunov, G. A. Lada, S. N. Litvinchuk, J. M. Rosanov, D. A. Shabanov, A. I. Zinenko // Russian Journal of Herpetology. – 2004. – Vol. 11, № 3. – P. 194–213. (Диссертант принимал участие в сборе материала и в интерпретации результатов).
Різноманіття зелених жаб (Rana esculenta complex) в Харківській області: морфологічний аспект вивчення / О. В. Коршунов, Т. В. Бабініч, О. І. Зіненко, Д. А. Шабанов // Біологія та валеологія. – 2004. – Вип. 6. – С. 24–30. (Диссертант поставил задание для исследования, принимал участие в обработке литературных данных и в интерпретации результатов).
Кравченко М. А. Оценка уникальности популяций Bufo viridis Laurenti, 1768 (Amphibia, Anura) с точки зрения рациональной экологической этики / М. А. Кравченко, Д. А. Шабанов // Вісник Дніпропетровського університету. – 2005. – № 3/2. – С. 91–95. (Диссертант поставил задание для исследования, предоставил эмпирический материал для его выполнения и принимал участие в интерпретации результатов).
Мазепа Г. О. Формалізований опис рострального рисунку зелених ропух (Bufo viridis) / Г. О. Мазепа, Д. А. Шабанов // Біологія та валеологія. –2005. – Вип. 7. – С. 77 –83. (Диссертант поставил задание для исследования, принимал участие в выборе и отработке методики, сборе материала, а также в интерпретации результатов).
Изучение популяционных систем зеленых лягушек (Rana esculenta complex) в Харьковской области: история, современное состояние и перспективы / Д. А. Шабанов, А. И. Зиненко, А. В. Коршунов, М. А. Кравченко, Г. А. Мазепа // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2006. – Вип. 3, № 729. – С. 208–220. (Диссертант поставил задание для исследования, принимал участие в обработке литературных и эмпирических данных полевых исследованиях и в интерпретации результатов).
До реорганізації зонування Національного природного парку «Гомільшанські ліси» / О. Ю. Акулов, Т. А. Атемасова, О. Ф. Бартенєв, С. Г. Вітер, А. С. Влащенко, О. І. Зіненко, О. В. Коршунов, Н. Б. Саідахмедова, Є. В. Скоробогатов, Д. А. Шабанов, А. Ю. Утєвський // Заповідна справа в Україні. – 2006. – Т. 12, Вип. 2. – С. 73–79. (Диссертант принимал участие в сборе материала и формулировании выводов работы в части, касающейся зеленых лягушек).
Эволюционно-генетические аспекты полуклонального воспроизводства гибридной формы Rana kl. esculenta (Amphibia, Ranidae) / С. В. Межжерин, С. Ю. Морозов-Леонов, О. Д. Некрасова, Ф. Ф. Куртяк, Д. А. Шабанов, А. В. Коршунов // Науковий вісник Ужгородського університету: Серія: біологія. –2007. – Вип. 21. – С. 79–84. (Диссертант принимал участие в сборе материала и в интерпретации результатов).
A case of natural triploidy in European diploid green toad (Bufo viridis), with some distributional records of diploid and tetraploid toads / L. J. Borkin, D. A. Shabanov, O. V. Brandler, O. V. Kukushkin, S. N. Litvinchuk, G.A. Mazepa, J. M. Rosanov // Russian Journal of Herpetology. – 2007. – Vol. 14, № 2. – P. 121–132. (Диссертант принимал участие в сборе материала, его обработке и в интерпретации результатов).
О южной границе распространения гибридной Rana esculenta (Ranidae, Anura, Amphibia) на территории Украины и Молдовы: данные проточной ДНК-цитометрии / Л. Я. Боркин, О. С. Безман-Мосейко, Г. А. Мазепа, А. И. Зиненко, А. В. Коршунов, Г. А. Лада, Д. А. Шабанов, С. Н. Литвинчук, Ю. М. Розанов // Праці Українського герпетологічного товариства. – 2008. – № 1.– С. 3–8. (Диссертант принимал участие в сборе материала и в интерпретации результатов).
Маро А. Н. Могут ли условия развития головастиков Bufo bufo определять темпы постметаморфического роста и созревания жаб? / А. Н. Маро, А. В. Шабанова, Д. А. Шабанов // Вопросы герпетологии. Мат-лы III съезда Герпетологического общ-ва им. А. М. Никольского. – СПб: 2008. – С. 274–280. (Диссертант поставил задание для исследования, принимал участие в обработке данных и в интерпретации результатов).
Кравченко М. А. Возможные пути трансформации популяционных систем Pelophylax esculentus cоmpleх (Ranidae, Anura, Amphibia) / М. А. Кравченко, Д. А. Шабанов // Праці Українського герпетологічного товариства. – № 1, 2008. – С. 15–20. (Диссертант принимал участие в постановке задания для исследования, обработке эмпирических данных и в интерпретации результатов).
Шабанов Д. А. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми / Д. А. Шабанов, О. В. Коршунов, М. О. Кравченко // Біологія та валеологія. – 2009. – Вип. 11. – С. 116–125. (Диссертант поставил задание для исследования, принимал участие в обработке литературных данных и в интерпретации результатов).
Коршунов А. В. Экологические факторы, влияющие на распространение представителей Pelophylax esculentus complex в Харьковской области / А. В.Коршунов. Д. А. Шабанов // Биологический вестник. – 2009. – Т. 13, № 1–2. – С. 76–83. (Диссертант принимал участие в постановке задания для исследования, выборе и отработке методики, сборе материала, а также в интерпретации результатов).
Кравченко М. А. Моделирование трансформаций гемиклональных популяционных систем зеленых лягушек (Pelophylax esculentus complex; Amphibia, Ranidae) с помощью рекуррентных разностных уравнений / М. А. Кравченко, Д. А. Шабанов // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2010. – Вип. 12, № 920. – С. 70–82. (Диссертант принимал участие в постановке задания для исследования, выборе методов для его исполнения и в интерпретации результатов).
Бондарєва А. А. Чи впливають розмірні ефекти клітин диплоїдних та триплоїдних їстівних жаб на їх стійкість до гіпоксії? / А. А. Бондарєва, Т. І. Махній, Д. А. Шабанов // Біологія та валеологія. – 2010. – Вип. 12. – С. 10–15. (Диссертант поставил задание для исследования, принимал участие в выборе и отработке методики, а также в интерпретации результатов).
Михайлова О. В. Як оцінити популяційний вантаж, що пов’язаний з геміклональною гібридизацією в популяційних системах Pelophylax esculentus complex? / О. В. Михайлова, О. Є. Усова, Д. А. Шабанов // Біологія та валеологія. – 2011. – Вип. 13. – С. 44–50. (Диссертант поставил задание для исследования, принимал участие в отработке методики и в интерпретации результатов).
Исследование устойчивости гемиклональных популяционных систем гибридогенного комплекса зеленых лягушек при помощи имитационного моделирования / М. А. Кравченко, Д. А. Шабанов, М. В. Владимирова, Г. Н. Жолткевич // Вісник Дніпропетровського університету. Біологія. Екологія. – 2011. – Т. 1, Вип. 19. – С. 51–64. (Диссертант принимал участе в постановке задания для исследования, выборе и отработке методики, а также в интерпретации результатов).
Михайлова О. В. Изучение сперматогенеза у диплоидных Pelophylax esculentus (Amphibia, Anura) при помощи кариоанализа в раздавленных препаратах / О. В. Михайлова, А. Е. Кечеджи, Д. А. Шабанов // Праці Українського герпетологічного товариства. – 2011. – № 3. – С. 120–127. (Диссертант принимал участе в постановке задания для исследования, выборе и отработке методики, а также в интерпретации результатов).
Шабанов Д. А. Гібридогенний комплекс зелених жаб як модель для вивчення багаторівневого добору / Д. А. Шабанов // Вісник Прикарпатського національного університету імені Василя Стефаника. Серія – Біологія. – 2012. – Вип. XVII. – С. 90–94.
Популяционная система зеленых лягушек (Pelophylax esculentus complex) Иськова пруда в Змиевском районе Харьковской области: история изучения и оценка численности / Е. В. Мелешко, А. Д. Суворова, М. А. Кравченко, Д. А. Шабанов // Биологический вестник. – 2012. – Т. 14, № 1. – С. 63–68. (Диссертант поставил задание для исследования, принимал участие в выборе и отработке методики, а также в интерпретации результатов).
Вивчення флуктуючої асиметрії річкового окуня (Perca fluviatilis L., 1758) / К. П. Виноградова, Ю. В. Сакун, К. М. Белоусова, Г. Л. Гончаров, Д. А. Шабанов // Біологія та валеологія. – 2012. – Вип. 14. – С. 9–17. (Диссертант поставил задание для исследования, принимал участие в отработке методики, а также в интерпретации результатов).
Бондарєва А. А. Порівняння деяких гематологічних показників диплоїдних та триплоїдних Pelophylax esculentus / А. А. Бондарєва, К. В. Сєдова, Д. А. Шабанов // Праці українського герпетологічного товариства. – 2013. – № 4. – С. 22–26. (Дисертант поставив завдання для дослідження, збирав матеріал та брав участь в інтерпретації результатів) (Диссертант поставил задание для исследования, собирал материал и принимал участие в интерпретации результатов).
Порушення фертильності у міжвидових гібридів зелених жаб із Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex / А. А. Боброва, Р. М. Макарян, В. П. Шейко, Д. А. Шабанов // Біологія та валеологія. – 2014. – Вип. 16. – С. 7–15. (Диссертант поставил задание для исследования, принимал участие в отработке методики, сборе материала, а также в интерпретации результатов).
Статьи в других изданиях
Массовая полиплоидия в гибридогенном комплексе Rana esculenta (Ranidae, Anura, Amphibia) на Востоке Украины / Л. Я. Боркин, А. И. Зиненко, А. В. Коршунов, Г. А. Лада, С. Н. Литвинчук, Ю. М. Розанов, Д. А. Шабанов // Матеріали Першої конференції Українського Герпетологічного Товариства. – К.: Зоомузей ННПМ НАНУ, 2005. – С. 23–26.
Маро А. Н. Механизмы расселения серой жабы (Bufo bufo (L. 1758); Amphibia, Anura) и особенности ее популяций в недавно заселенных местообитаниях / А. Н. Маро, Д. А. Шабанов // Матеріали Першої конференції Українського Герпетологічного Товариства. – К.: Зоомузей ННПМ НАНУ, 2005. – С. 107–110.
Исследование целостных свойств биосистем на примере имитационного моделирования гибридогенных популяционных систем зеленых лягушек / М. В. Владимирова, Г. Н. Жолткевич, А. А. Луцик, Д. А. Шабанов // Вісник Харківського національного університету. Серія «Математичне моделювання. Інформаційні технології. Автоматизовані системи управління» – 2007. – № 780. – C. 61–70.
Шабанов Д. А. Региональная сводка, учебник, справочник и научная монография. Рецензия на книгу В. Л. Булахова, В. Я. Гассо, А. Е. Пахомова «Биологическое разнообразие Украины. Днепропетровская область. Земноводные и пресмыкающиеся (Amphibia et Reptilia)» / Д. А. Шабанов, А. И. Зиненко // Вісник Дніпропетровського університету. Біологія. Екологія. – 2008. – Вип. 16, т. 2. – С. 209–211.
Шабанов Д. А. Почему раздельнополые организмы вытесняют перекрестнооплодотворяющихся гермафродитов: раздельнополость как равновесие по Нэшу / Д. А. Шабанов // Мат. Междунар. науч. конф. посв. 80-летию со дня рожд. проф. А. П. Крапивного. – Х.: ХНУ им. В. Н. Каразина, 2009. – С. 38–49.
Шабанов Д. А. Зеленые лягушки: жизнь без правил или особый способ эволюции / Д. А. Шабанов, С. Н. Литвинчук // Природа. – 2010. – № 3 (1135). – С. 29–36.
Vlasova T. Simulation of the Expanded Iterated Version of the Prisoner's Dilemma Game Recognisers / T. Vlasova, M. Vladymyrova, D. Shabanov // 7-th Int. Conf. ICTERI, 2011. –Vol. 716. – P. 82–86.
Тезисы в материалах конференций
Шабанов Д. А. Уникальность популяций бесхвостых амфибий как основание для их охраны / Д. А. Шабанов, А. В. Коршунов, М. А. Кравченко // Современные проблемы зоологии и экологии. – Одесса: Феникс, 2005. – С. 328–329.
Кравченко М. А. К разработке методов оценки ценности популяций с точки зрения рациональной экологической этики / М. А. Кравченко, Д. А. Шабанов // Биоразнообразие и роль зооценоза в естественных и антропогенных экосистемах: Мат. III Междунар. научн. конф. – Днепропетровск: Изд-во ДНУ, 2005. – С. 78–80.
Коршунов А. В. Преобразование популяционных систем зеленых лягушек как особая категория процессов естественного развития / А. В. Коршунов, М. А. Кравченко, Д. А. Шабанов // Современные проблемы популяционной экологии. Мат. IX Междунар. научно-практ. экол. конф. – Белгород: Изд-во ПОЛИТЕРРА, 2006. – С. 101–102.
Шабанов Д. А. Гибридные лягушки Rana esculenta, производящие гаметы обоих родительских форм, – результат независимой клональной эволюции геномов? / Д. А. Шабанов // Материалы конференции «Современные проблемы биологической эволюции. К 100-летию Государственного Дарвиновского музея». – Москва, Изд-во ГДМ, 2007. – С. 199–201.
Кравченко М. А. Иерархия уровней биоразнообразия на примере гибридогенного Rana esculenta complex / М. А. Кравченко, Д. А. Шабанов // Zoocenosis-2007. Біорізноманіття та роль тварин в екосистемах. – Дніпропетровськ, ДНУ, 2007. – С. 381–383.
Кравченко М. А. Что нового можно узнать о зеленых лягушках при помощи имитационного моделирования? (Пленарный доклад на открытии конференции) / М. А. Кравченко, А. А. Луцик, Д. А. Шабанов // «Біологія: від молекули до біосфери». Матеріали III Міжнародної конференції молодих науковців. – Харьков: СПД ФО Михайлов Г. Г., 2008. – C. 9–11.
Усова Е. Е. Об оптимизации методики ретроспективной оценки динамики размеров тела представителей Pelophylax esculentus complex (Amphibia, Ranidae) при помощи скелетохронологии / Е. Е. Усова, Д. А. Шабанов // Zoocenosis-2009. Біорізноманіття та роль тварин в екосистемах. – Дніпропетровськ, ДНУ, 2009. – С. 278–280.
Материалы по гельминтофауне зеленых лягушек (Rana esculenta complex) в Харьковской области области / М. В. Резванцева, Г. А. Лада, Д. С. Аксенов, Д. А. Шабанов, А. В. Коршунов, И. В. Чихляев, Л. Я. Боркин, С. Н. Литвинчук, Ю. М. Розанов // Теоретические и практические проблемы паразитологии. – Материалы Международной научной конференции. – М., 2010. – С. 308 –312.
Шабанов Д. А. К разработке классификации типов популяционного воспроизводства / Д. А. Шабанов // Видовые популяции и сообщества в антропогенно трансформированных ландшафтах: состояние и методы его диагностики. — Мат. XI Междунар. научно-практич. экологич. конф. – Белгород, 2010. – С. 228–229.
Кравченко М. А. Уровни биоразнообразия Pelophylax esculentus complex / М. А. Кравченко, Д. А. Шабанов // Биоразнообразие и устойчивое развитие. Тезисы Междунар. научно-практ. конф. – Симферополь: КНЦ, 2010. – С. 68–71.
Власова Т. Имитационное моделирование расширенного итерированного варианта игры «дилемма заключенного» / Т. Власова, М. Владимирова, Д. Шабанов // ICTERI 2011. ІКТ в освіті, дослідженнях та індустріальних додатках: інтеграція, гармонізація та трансфер знань. – Херсон: ХДУ, 2011. – С. 57–58.
Бондарева А. А. Сравнение размера и формы эритроцитов у диплоидных и триплоидных гибридов зеленых лягушек (Pelophylax esculentus) / А. А. Бондарева, Д. А. Шабанов // Всеукраїнський конкурс студентських наукових робіт з біологічних наук: матеріали підсумкової науково-практичної конференції (Мелітополь, 29-31 березн. 2011 р.). – Мелітополь: «Люкс», 2011. – С. 11–14.
Cytological maps of lampbrush chromosomes of European water frogs / D. Dedukh, G. Mazepa, D. Shabanov, S. Litvinchuk, L. Borkin, A. Saifitdinova, A. Krasikova // 18th International Chromosome Conference (Manchester, 29 August – 2 September 2011). Speaker abstracts. – P. 48–49.
АНОТАЦІЯ
Шабанов Д. А. Еволюційна екологія популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) Лівобережного лісостепу України. — На правах рукопису.
Дисертація на здобуття наукового ступеня доктора біологічних наук за спеціальністю 03.00.16 — екологія. — Дніпропетровський національний університет імені Олеся Гончара. — Дніпропетровськ, 2015.
Наведено результати багаторічних досліджень Pelophylax esculentus complex у Лівобережному лісостепу України. У регіоні дослідження виділено п’ять субрегіонів, що відрізняються за складом популяційних систем і популяцій представників Pelophylax esculentus complex, що поширені у них. Популяційні системи, що включають Pelophylax esculentus, зустрічаються в чотирьох із цих субрегіонів, триплоїди Pelophylax esculentus зареєстровані у двох із них. Описано Сіверсько-Донецький центр різноманіття Pelophylax esculentus complex.
Геміклональне спадкування в Pelophylax esculentus є причиною появи геміклональних популяційних систем (ГПС). Для дослідження трансформацій ГПС Pelophylax esculentus complex створена імітаційна модель. З її використанням розроблено динамічну типологію ГПС, що включають диплоїдних представників Pelophylax esculentus complex. Показано, що Pelophylax esculentus complex надає виняткові можливості для вивчення багаторівневого відбору. Запропоновано гіпотези, що пояснює феномен гібридної амфіспермії та пояснює добір на здатність до геміклональної гібридизації.
Ключові слова: екологія, еволюція, Pelophylax esculentus complex, P. ridibundus, P. lessonae, P. esculentus, геміклональні популяційні системи, імітаційне моделювання, багаторівневий добір, стійкість.
АННОТАЦИЯ
Шабанов Д. А. Эволюционная экология популяционных систем гибридогенного комплекса зеленых лягушек (Pelophylax esculentus complex) Левобережной лесостепи Украины. — На правах рукописи.
Диссертация на соискание научной степени доктора биологических наук по специальности 03.00.16 — экология. — Днепропетровский национальный университет имени Олеся Гончара. — Днепропетровск, 2015.
Приведены результаты многолетних исследований Pelophylax esculentus complex в Левобережной лесостепи Украины. В регионе исследования выделены пять субрегионов, отличающихся по составу распространенных в них популяционных систем и популяций представителей Pelophylax esculentus complex. Популяционные системы, включающие Pelophylax esculentus, встречаются в четырех из этих субрегионов, триплоиды Pelophylax esculentus зарегистрированы в двух из них. Различия субрегионов связаны частично с различием характерных для них биогеоценотических условий, а частично — с изоляцией бассейнов разных рек. Два субрегиона, расположенных в водосборном бассейне реки Северский Донец, образуют Северско-Донецкий центр разнообразия Pelophylax esculentus complex.
Показано, что характерные свойства биосистем различных уровней организации тесно связаны со способом рекомбинации, гаметогенеза, размножения и популяционного воспроизводства. Гемиклональное наследование у Pelophylax esculentus связано с глубоким изменением этой цепочки процессов. Воспроизводство Pelophylax esculentus возможно лишь в составе гемиклональных популяционных систем (ГПС), являющихся особой категорией биосистем.
Для исследования механизма трансформаций ГПС Pelophylax esculentus complex создана имитационная модель, позволяющая тестировать гипотезы, относящиеся к неизученным аспектам функционирования ГПС, на соответствие комплексу накопленных эмпирических данных. С ее использованием разработана динамическая (объединяющая рассматриваемые объекты в группы на основе прогноза их будущей динамики) типология ГПС, включающих диплоидных представителей Pelophylax esculentus complex. Обоснована теоретическая возможность существования гемиклональных популяционных систем, состоящих исключительно из диплоидных Pelophylax esculentus.
Показано, что Pelophylax esculentus complex предоставляет исключительные возможности для изучения многоуровневого отбора. В нем зарегистрированы проявления отбора на генном, геномном, клеточном, организменном и групповом уровнях. Выдвинута гипотеза, объясняющая феномен гибридной амфиспермии конкуренцией в клетках зародышевого пути клональных геномов, измененных вследствие отбора на способность вызывать элиминацию альтернативных геномов. Выдвинута гипотеза, согласно которой способность двух относительно близких видов к гемиклональной гибридизации является приспособлением к гомеологичной межвидовой рекомбинации, поддерживаемым на уровне отбора видов.
Ключевые слова: экология, эволюция, Pelophylax esculentus complex, P. ridibundus, P. lessonae, P. esculentus, гемиклональные популяционные системы, имитационное моделирование, многоуровневый отбор, устойчивость.
Annotation
Shabanov D. A. Evolutionary ecology of population systems of green frogs’ hybridogenetic complex (Pelophylax esculentus complex) from Left Bank forest-steppes of Ukraine. — The Rights of the manuscript.
Dissertation for a degree of Doctor of Biological Sciences by speciality 03.00.16. – Ecology. – Oles’ Gonchar Dnipropetrovsk National University. – Dnipropetrovsk, 2015.
The results of long-term research of Pelophylax esculentus complex in Left Bank forest-steppes of Ukraine are represented. The 5 sub-regions were distinguished in the study region. They differ in composition of widespread population systems and populations of Pelophylax esculentus complex representatives. Population systems, which include Pelophylax esculentus, were found in four of these regions, Pelophylax esculentus triploids were registered in two of them. The Seversko-Donetskiy center of green frogs' diversity was described.
The hemiclonal inheritance is the reason of hemiclonal population systems (HPS) origin. The simulation model was developed for studying the transformation of HPS of Pelophylax esculentus complex. The dynamic typology of HPS was developed using this model. It was shown, that Pelophylax esculentus complex gives exceptional opportunities for studying the multilevel selection. The hypothesis explaining the hybrid-amphispermy and selection for hemiclonal hybridization ability were proposed.
Key words: ecology, evolution, Pelophylax esculentus complex, P. ridibundus, P. lessonae, P. esculentus, hemiclonal population systems, simulation modelling, multilevel selection, stability.