Шишкин М.А. Эволюционная теория и научное мышление. // Палеонтологический журнал, 2010. — № 6. — с. 3-17
УДК 576.12:001.8
ЭВОЛЮЦИОННАЯ ТЕОРИЯ И НАУЧНОЕ МЫШЛЕНИЕ
© 2010 г. М. А. Шишкин
Палеонтологический институт им. А.А. Борисяка РАН e-mail: shishkin@paleo.ru Поступила в редакцию 25.12.2009 г.
Принята к печати 29.01.2010 г.
Прогресс теоретической мысли и рост фактического знания в естественных науках не связаны прямой зависимостью. Состояние теории надолго определяется особенностями коллективного мышления, сложившимися в данной области знаний. Это особенно относится к концепции естественного отбора, известной в виде двух полярных интерпретаций — генетической теории эволюции и эпигенетической теории. Предсказуемый итог их противостояния не будет связан с “решающими аргументами” в пользу одной из них. Его наступление зависит от того, насколько быстро эволюционная биология освободится от традиции мозаичного мышления, позволяющего объединять взаимоисключающие концепции. Ключевым в этом плане будет осознание несовместимости идеи корпускулярной детерминации развития с принципом системной обусловленности того же процесса.
ВВЕДЕНИЕ
Прогресс теоретической мысли и рост фактического знания в естественных науках согласуются друг с другом лишь в общих чертах. Более того, экстенсивное накопление эмпирических наблюдений может отбрасывать теоретическую мысль назад. На конкретных исторических этапах решающими для состояния теории могут быть совсем иные факторы. Прежде всего, это особенности и традиции коллективного научного мышления, сложившиеся в данной области знаний. Все это относится и к теории биологической эволюции на основе естественного отбора.
В дарвиновском изложении теории ее базовые понятия (отбор, изменчивость и наследственность) слабо конкретизированы, что было естественно при тогдашнем уровне биологических представлений и отсутствии у Дарвина склонности к умозрительным конструкциям. Им во многом было лишь намечено то, что оказалось осознанным значительно позже. Особенно отметим близость творца теории к интуитивному пониманию роли естественного отбора как механизма стабилизации исторических изменений: “отбор в этих случаях еще не успел осилить наклонности к дальнейшей изменчивости...”; или: “...отбор уже успел сообщить... органу постоянный характер” (см. Дарвин 2003, с. 166). Отсюда же логически следовало и объяснение им изменчивости рудиментарных органов как результата отсутствия селективного контроля (там же, с. 165). Глубинный смысл этих обобщений не мог быть в то время понят по причине отсутствия представлений о системных процессах.
В дальнейшем дарвиновская теория получила две диаметрально полярных интерпретации, исходно связанные с именами А. Вейсмана (неодарвинизм, или генетическая теория эволюции — ГТЭ) и И.И. Шмальгаузена—К. Уоддингтона (учение о стабилизирующем отборе, или эпигенетическая теория — ЭТЭ). Они опираются на альтернативные концепции индивидуального развития, т.е., соответственно, на преформационную (прямо или по умолчанию) и эпигенетическую модели. Первая из них предполагает ту или иную форму упорядоченного соответствия между элементами зародышевой клетки и частями взрослого организма. Напротив, в рамках второй модели эти категории относятся к разным иерархическим уровням системы развития и не связаны прямыми зависимостями.
В представлениях ГТЭ субстратом отбора является мозаика независимых наследственных факторов (генов) в половых клетках. Устойчивость (наследуемость) черт взрослого фенотипа есть выражение имманентных свойств этих факторов, не требующее эволюционного объяснения. Эволюция организма и его онтогенеза есть результат отбора генов; поэтому она протекает, как и сам онтогенез, в направлении от зиготы к взрослому состоянию. Наследственные изменения (мутации) и естественный отбор (в качестве сита для них) взаимодействуют в эволюции как независимые факторы. Наследственные и ненаследственные изменения фенотипа качественно различны.
Основа этого мировоззрения наглядно выражена в словах Вейсмана: “Естественный подбор только по видимости имеет дело с качественными особенностями готового организма; на самом же деле лишь со скрытыми в половых клетках зачатками этих особенностей” (Weissmann, 1883; цит. по Филипченко, 1977, с. 148).
Напротив, для ЭТЭ объектом эволюции является индивидуальное развитие как целое, т.е. онтогенез рассматривается как динамическая система, направленная к конечному равновесию (взрослой норме). Эволюция есть цепь актов репарации такой системы, следующих после каждого нарушения ее устойчивости. Отбор есть механизм самоорганизации, т.е. поиска нарушенной системой нового равновесия путем стабилизации одного из реализуемых уклонений развития. Материал отбора — это гетерогенные особи-изоаберранты, а не носители “отбираемой мутации”. Эволюционные изменения начинаются с взрослого фенотипа и распространяются в направлении генотипа последующих поколений1. Наследственность (устойчивость) — не партнер естественного отбора, а его продукт, выступающий как системное свойство развития. Все изменения фенотипа суть выражения нормы реакции и в этом качестве однородны.
Эти принципы отражены, в частности, в следующих обобщениях: “Хотя изменение генотипа и является необходимой базой для эволюционного процесса, но не оно определяет эволюцию... Наоборот, эволюция организма определяет изменения его генотипа...” (Шмальгаузен, 1940, с. 57). “Теория естественного отбора является ... теорией, объясняющей возникновение и преобразование наследственных механизмов. ...Устойчивость... признака... не свойство гена, а выражение взаимозависимости... частей ... развивающегося организма” (Шмальгаузен, 1982, с. 110, 174) (курсив наш — МШ).
Согласно ЭТЭ, пространство возможных аберраций развития в данной системе составляет ее видоспецифичную характеристику. Любое отклонение от нормального итога развития остается в пределах этого ограниченного пространства и представляет целостный ответ системы на нарушение должной координации событий в ней (рис. 1). Он отражает, таким образом, свойства самой системы, а не специфику того или иного внутреннего или внешнего повреждающего фактора (ср. Goldschmidt, 1938, 1940). В сумме это пространство реагирования описывается как совокупность онтогенетических траекторий, осуществимых в данном типе развития с той или иной вероятностью. Его наглядной трехмерной моделью является эпигенетический ландшафт К. Уоддингтона в виде сети дивергирующих долин, обозначающих возможные варианты развития (рис. 2, а). Демонстрируемая этой моделью относительно простая структура “есть свойство высшего порядка, основанное на подстилающей сети гораздо более сложных взаимодействий” (Waddington, 1957, с. 34, рис. 5), т.е. процессов, протекающих на уровнях генной экспрессии и элементарных морфогенетических актов (рис. 2, б).
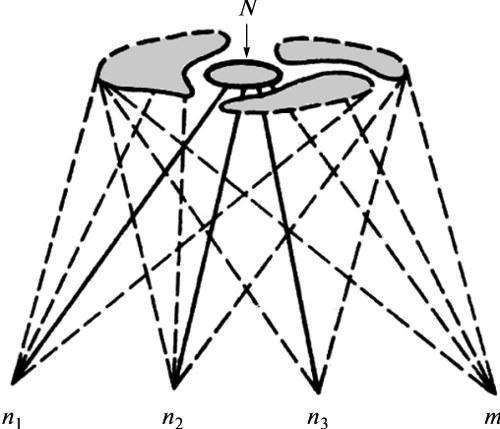
Рис. 1. Видоспецифичная ограниченность пространства итогов развития, не зависящая от природы возмущающих факторов (по Шишкину, 1987, с изменениями). Обозначения: N — область вариаций взрослой фенотипической нормы; поля вокруг нее — области аномального формообразования; n1—n3 — зиготы с высокой помехоустойчивостью нормального развития, m — зигота с дестабилизированным развитием; сплошные линии — устойчивые (для индивидуальных зигот) траектории развития, прерывистые — неустойчивые траектории.

Рис. 2. а — система развития в виде “эпигенетического ландшафта”, показывающего возможные траектории онтогенеза; б — нелинейная природа соотношений между ролью индивидуальных генов, функционирующих в ходе развития, и структурой эпигенетического ландшафта (по Waddington, 1957).
Из сказанного ясно, что в представлениях ЭТЭ не существует фенотипических новшеств, привносимых мутациями. “Естественный отбор... создает эпигенетический ландшафт, который... делает возможными фенотипические эффекты мутаций. В этом свете привычное утверждение, что сырой материал эволюции создается случайными мутациями, является пустым” (Waddington, 1957, с. 188). Иначе говоря, “мутационные” изменения фенотипа реализуют лишь те или иные из существующих возможностей реагирования, свойственных данной системе развития (Шишкин, 2006). Собственно же эволюционное изменение такой системы, согласно ЭТЭ, означает перестройку ее эпигенетического ландшафта, т.е. создание в нем новой канализированной траектории и, соответственно, изменение общего спектра аберраций развития (рис. 3).
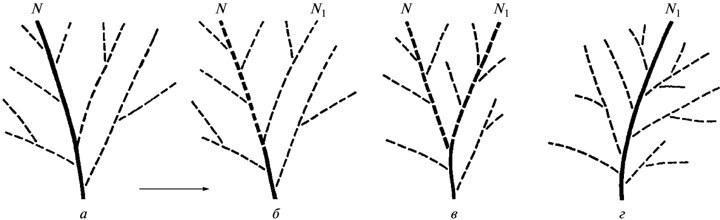
Рис. 3. Элементарное эволюционное изменение (а—г) как перестройка системы развития в ходе стабилизации одной из ее прежних аберративных траекторий (по Шишкину, 1987, с изменениями): а — исходное состояние; б, в — частичная дестабилизация прежней фенотипической нормы и переход к состоянию с двумя преобладающими путями развития; г — закрепление новой нормы в качестве единственной. Обозначения: N — прежняя норма, N1 —вновь возникшая норма; жирными линями выделены нормальные пути развития (креоды) на разных стадиях их стабилизации или утраты.
Суть различия между двумя рассмотренными альтернативами дарвинизма может быть выражена следующим образом. Для ГТЭ (рис. 4, а) каждое элементарное эволюционное событие есть продукт единичного онтогенетического цикла, реализуемого на основе измененной (мутантной) зародышевой клетки. Тем самым это по определению сальтационистская теория. Напротив, для ЭТЭ (рис. 4, б) каждое эволюционное событие — это всегда процесс, охватывающий череду поколений, где начальное фенотипическое уклонение проявляется раньше, чем формируется стабилизированный (наследственно устойчивый) механизм его осуществления.
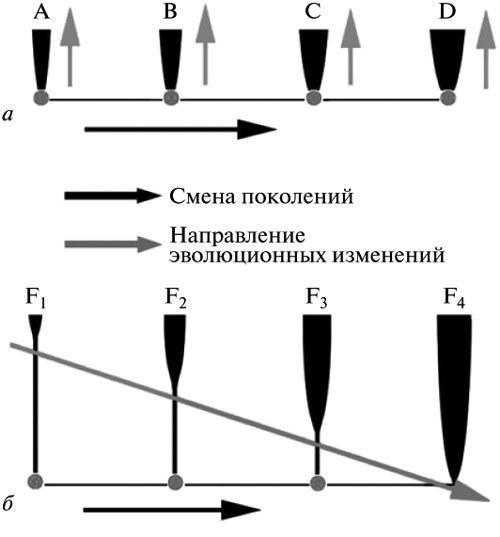
Рис. 4. Механизм эволюции в понимании двух альтернативных версий селекционизма: а — генетическая теория (ГТЭ), б — эпигенетическая теория (ЭТЭ). ГТЭ: элементарное эволюционное событие равнозначно мутации, содержащейся в зародышевой клетке, и осуществляется в ходе одного онтогенетического цикла. ЭТЭ: становление элементарного новшества есть процесс его стабилизации, распространяющийся в поколениях от взрослой стадии к зародышевой клетке. Обозначения: А—D — онтогенетические циклы, связанные с появлением последовательных эволюционных изменений (в ГТЭ); F1—F4 — этапы становления одного эволюционного изменения (в ЭТЭ).
Конечный итог противостояния этих концепций представляется вполне предрешенным, но основой для него с очевидностью не будет служить предъявление “решающих аргументов”, убедительных для обеих сторон. Судьба теорий, как и обстоятельства их возникновения, обычно определяются иначе (см. раздел VI). Рассмотрим некоторые существенные в этом случае факторы.
I. ТЕОРИИ И ИНТЕРПРЕТАЦИЯ ФАКТОВ
Возникновение теории — эвристический акт, не сводимый к логическим операциям. Теории не строятся непосредственно из фактов; наоборот, последние получают освещение в свете теории (Любищев, 1925). От выбора постулатов теории (не обязательно осознанного) зависит, какие факты для нее существенны и предсказуемы, а какие относятся к информационным шумам, затемняющим должный ход событий. Например, все основные “шумы” ГТЭ — это как раз область предсказуемых явлений для ЭТЭ (табл. 1).
Таблица 1. Феномены, предсказуемые для ЭТЭ, и возможности их истолкования в рамках ГТЭ
|
Феномены, предсказуемые для ЭТЭ |
Возможности их истолкования в рамках ГТЭ |
|
Устойчивость нормального (“дикого”) фенотипа |
“генетический гомеостаз”? |
|
Гетерогенность природных популяций |
“частотозависимый отбор”? |
|
Преобладание не-менделевского наследования аномалий в популяциях |
влияние “генотипической среды”? |
|
Возможность изменения “генетической детерминации” признака |
? |
|
Параллелизм мутационной и модификационной изменчивости |
? |
Тот же выбор постулатов предопределяет и рабочие понятия (язык) теории. Вне ее они либо лишены содержания, либо могут иметь совершенно иное значение. Например, понятия отбора мутаций или генетического дрейфа лишены смысла вне рамок ГТЭ; и, наоборот, представление о стабилизации развития для этой теории является беспредметным. Подобным же образом естественный отбор означает для двух версий дарвинизма диаметрально разные вещи: для ГТЭ это сито для мутаций, а для ЭТЭ — механизм преобразования целостных свойств живой организации. Наследственность (см. выше) — в первом случае имманентное свойство материальных частиц, во втором — эпифеномен внутрисистемной регуляции, созданной отбором.
По этой же причине одно из ключевых понятий неодарвинизма — “наследственная изменчивость”, обозначающее вариации, привносимые мутациями, является в рамках ЭТЭ бессмысленным, так как для нее наследственность — это устойчивость, а изменчивость — отсутствие таковой. Вариации здесь наследственны лишь исключительно как целое, т.е. как специфический спектр изменчивости, характеризующий данную норму реакции и поэтому передаваемый в поколениях, с неопределенной вероятностью индивидуального наследования. Но в этом качестве все вариации (онтогенетические траектории) для ЭТЭ равноценны и различаются лишь по частоте своего осуществления.
Наконец, из базовых постулатов теории вытекают и характерные для нее представления о “пространстве возможностей”, мыслимом для альтернативных (отвергаемых ею) теоретических позиций. Так, для ГТЭ принципиальный водораздел проходит между ее представлением, что эволюционные изменения начинаются с зародышевой клетки, и альтернативным мнением, что они начинаются с взрослой организации. Вторая позиция автоматически отождествляется ГТЭ с ламаркизмом, к разновидностям которого, естественно, причисляют в этом случае и теорию Шмальгаузена. Напротив, для этой последней (ЭТЭ) главный водораздел лежит между объяснением органической целесообразности (наследственной устойчивости, ср. Шишкин, 2006) как продукта эволюционного процесса и фактическим отказом от такого подхода. С этой точки зрения ламаркизм и генетическая теория одинаково попадают для ЭТЭ во вторую категорию. Ибо если в глазах ламаркизма наследственность “создается” самой полезностью нового приобретения, то в рамках ГТЭ она сходным же образом тавтологически объясняется действием соответствующего фактора наследственности.
Все эти свойства теории демонстрируют частный пример общего правила, что характер и оценка считываемой информации всегда определяются особенностями восприятия реципиента (в данном случае спецификой его мировоззрения). В этом отношении наглядный пример дают взгляды самого Ч. Дарвина. Если учесть, что он заслуженно считается образцом научной беспристрастности, то восприятие им некоторых биологических обобщений имеет для нас ценность своеобразного “контрольного опыта”. Речь здесь пойдет об оценке Дарвиным двух явлений, известных уже в его время: (1) появления предковых черт при гибридизации и (2) возвращения культурных форм к исходному типу при одичании в природе.
С точки зрения сегодняшних представлений о системе развития, причина этих явлений в конечном счете одна и та же. Она связана с сохранением в эпигенетическом пространстве исследуемых форм не вполне стертых предковых траекторий развития. Их реализация все еще может быть спровоцирована — либо сразу же, благодаря неустойчивости развития у гибрида (1), либо путем повторной стабилизации такой траектории как результат возобновления отбора в пользу прежней природной нормы (2).
Рассмотренные с этих позиций, оба явления одинаково хорошо согласуются с дарвиновской теорией. Но для самого Дарвина, при существовавшем тогда уровне естественнонаучных представлений, это далеко не представлялось очевидным. Он поддерживал эволюционное истолкование первого феномена, видя в нем проявление черт далекого предка, но выражал сдержанные сомнения в достоверности второго, т.е. примеров одичания (Дарвин, 2003, с. 31, 32, 160—164). Одна из причин этого скептицизма заключалась в том, что такие факты в то время использовались против дарвиновской теории (там же, с. 31) — как свидетельство, что процессы изменений организмов в природе не описываются механизмом искусственной селекции (что само по себе, безусловно, верно). Этот пример иллюстрирует сказанное выше: оценка фактов исследователем объективно определяется тем, насколько они кажутся ему совместимыми с собственной системой взглядов.
II. ВЗАИМОВОСПРИЯТИЕ И ЗАМЕЩЕНИЕ АЛЬТЕРНАТИВНЫХ ТЕОРИЙ
В теориях, построенных на качественно различных основаниях, понимание причинных связей между явлениями в общем случае несопоставимо, вследствие чего суждения одной из них не могут быть переведены на язык другой. Поэтому возможность содержательного диалога между ними обычно исключена. В широком смысле это касается не только научных обобщений, но и вообще любых разнокачественных типов мировосприятия, свойственных живым существам. Носители каждого из таких вариантов способны отображать явления окружающего мира лишь в рамках доступных им информационных сигналов. Эта ситуация наглядно передана в афористической притче В. Маяковского (1928) о лошади и верблюде, где каждое из этих животных видит в другом лишь уродливый (“неправильный”) экземпляр собственного вида.
Эта невозможность коммуникации между альтернативными системами восприятия носит объективный характер. То есть, если говорить о теориях, представления одной из них о содержании другой всегда преломляются через специфику ее собственного видения. Непонимание в этом случае может быть и односторонним, если одна из конкурирующих концепций, будучи обобщением более высокого порядка, способна показать на своем языке реальное место причинных связей, абсолютизируемых ее оппонентом. Но именно односторонняя слепота оказывает фатальное влияние на прогресс теоретического знания, если она свойственна господствующей системе взглядов. Примеры такого рода заставляют думать, что прогресс теоретической мысли определяется даже не собственно борьбой идей. Решающую роль здесь играет сама готовность научного сообщества к осознанию того, что господствующие постулаты не являются единственно возможным выбором.
Одной из наглядных иллюстраций сказанного является восприятие эволюционизмом концепции Р Гольдшмидта о системе реагирования (развития) как объекте эволюции. Эта идея лежит в основе представлений ЭТЭ. Ее будущее признание в качестве ключевого положения интегральной эволюционной теории представляется нам неизбежным. По существу Гольдшмидт показал, что анализ развития генетическими методами подтверждает целостность онтогенетического процесса, вскрытую ранее экспериментальной эмбриологией. Это тождество результатов открывает путь к интерпретации генетических обобщений в свете общих законов развития, т.е. к решению задачи, которая представляется наиболее актуальной для эволюционной теории (ср. Шишкин, 1987, 2006).
Тем не менее, указанная идея осталась вне круга интересов доминирующего направления эволюционной мысли ХХ века. Уже несколько поколений исследователей обсуждают гипотезу “системных мутаций” Гольдшмидта, с очевидностью не замечая причины, обусловившей ее появление. Между тем она заключалась именно в осознании, что эволюционные изменения не сводятся к эффектам независимых генов. Но такая позиция несовместима с генетическим мышлением и потому просто не воспринимается им. По той же причине сегодняшние попытки этого мышления найти место для целостных механизмов развития в “новом эволюционном синтезе” мало что изменили в его воззрениях на эволюцию (см. раздел VI).
Эту ситуацию иллюстрирует, в частности, книга Р. Рэффа и Т. Кофмена (1986) “Эмбрионы, гены и эволюция” — одно из первых крупных обобщений в направлении “нового синтеза”. Книга посвящена Р Гольдшмидту, и авторы считают себя последователями его идеи об эволюции как системной трансформации развития. Но они убеждены, что Гольдшмидт подразумевал под этим “изменения в генах, регулирующих онтогенез” (Рэфф, Кофмен, 1986, с. 13), т.е. процессы типа гомеозисных мутаций. Насколько это далеко от позиции Гольдшмидта, можно понять хотя бы из следующих его слов: “Факты генетики, конечно, могут описываться в терминах генов, но теория зародышевой плазмы должна полностью освободиться от концепции генов как единиц” (Goldschmidt, 1938, с. 311). “Зародышевая плазма как целое контролирует определенную реактивную систему, которая есть. единая система развития.” (Goldschmidt, 1940, с. 218). Характерно, что автор этих выводов не питал иллюзий насчет того, как они будут восприняты: “Для многих генетиков явно трудно мыслить в таких понятиях, поскольку большинство их настолько связаны аксиоматической верой в атомистическую генную теорию, что не в состоянии думать иначе.” (там же).
Переход от редукционистского видения законов эволюции к организмической (системной) модели ЭТЭ не был простым даже для создателей этой последней. У И.И. Шмальгаузена и К. Уод-дингтона мы находим немало эклектических суждений, где смешиваются оба эти подхода (ср. Шишкин, 1987, с. 99; 2006, с. 184). Там, где эти противоречия были преодолены, это достигалось далеко не сразу. Характерный пример — понимание Шмальгаузеном стабилизирующей функции отбора. Выдвинутая им концепция с самого начала подразумевала принципиально новое понимание наследственной устойчивости — как целостного выражения корреляций развития, созданных отбором. Этот подход логически исключал разделение фенотипических свойств на продукты мутаций и экзогенных воздействий. И, тем не менее, в ранних работах по этому вопросу Шмальгаузен (1940, 1941) описывает механизм селективной стабилизации именно в этих редукционистских терминах — как накопление “совпадающих мутаций” под покровом ненаследственной “фенокопии”, т.е. в согласии с гипотезой совпадающего (органического) отбора Моргана-Болдуина! Этот пример показывает, что становление качественно нового мировоззрения — процесс многоступенчатый, и что ключевая идея здесь может появляться много раньше, чем ее непротиворечивое обоснование.
Сказанное можно подытожить следующим образом. Устойчивая система научных взглядов, как и всякая эффективно регулируемая система, невосприимчива к радикальным новшествам, т.е. реагирует на вызываемые ими возмущения как на обратимые (релаксируемые) флуктуации, иными словами, “не замечает” их. И лишь снижение способности такой теории к самоподдержанию (по мере исчерпания ее объяснительного потенциала) делает ее все более предрасположенной к изменениям. Иначе говоря, между двумя стабильными состояниями системы в процессе ее качественной трансформации всегда лежит неустойчивая область фазового перехода, где периоды погашения флуктуаций прежнего состояния все более удлиняются, вплоть до того, что становится возможным их “запоминание”. С этим, в конечном счете, связано известное наблюдение, что смена теоретических убеждений обычно происходит лишь со сменой поколений, когда старая парадигма еще не вполне укрепилась в сознании ее новых носителей.
III. ПРОЦЕСС ПОЗНАНИЯ
Специфичность отражения реальности любым реципиентом означает неполноту этого отражения. Для человеческого восприятия эта особенность усугубляется преимущественным использованием языковых символов. Пространство зависимостей в реальном мире бесконечно и взаимосвязано, тогда как наличные средства языка ограничены и дискретны. В итоге любое научное утверждение есть всегда продукт абстрагирования от бесконечного множества не улавливаемых причинных связей, определивших исследуемое событие. Любой вывод лишь условно отражает реальность, в соответствии с утверждением поэта, что “мысль изреченная есть ложь”. Это абстрагирование достигает максимума в выводах, полученных из непосредственного наблюдения или опыта.
Объективно цель познания — постижение общих законов бытия; но именно конкретное опытное знание является необходимой основой этого процесса в естественных науках. При этом кажущаяся неоспоримость причинных зависимостей, напрямую вскрываемых опытом, издавна фетишизировала этот подход в глазах естествоиспытателей как собственно цель познания. Путь к ней понимался как расчленение изучаемого явления на составные элементы и поиски (локализацию) непосредственной причины для каждого из них (Bertalanffy 1969). В частности, с такой детерминистской программы началась история экспериментальной эмбриологии, именовавшейся на рубеже XIX—XX вв. “механикой развития”. Этот подход, в качестве подлинного пути познания движущих сил онтогенеза, противопоставлялся натурфилософским объяснениям типа геккелев-ского биогенетического закона (Gould, 1977). В этой же колее протекали и поиски механизмов наследственности, начиная от умозрительных редукционистских концепций Негели и Вейсмана и кончая появлением хромосомной генетики.
Но, как уже сказано, любые выводы, полученные из опыта, всегда упрощают реальные соотношения. Обычно это обнаруживается уже при расширении или изменении условий эксперимента. Начальная объяснительная схема, в которой для каждого свойства объекта усматривалась отдельная причина (рис. 5, а), теряет свою эффективность по мере того, как выясняется, что одно и то же свойство может иметь разные причины и наоборот (рис. 1; 5, б). Такая асимметрия видимых причин и следствий с очевидностью указывает, что между ними на самом деле нет линейных зависимостей и что эти две категории свойств связаны лишь опосредованно, через системные отношения (Waddington, 1957; Белоусов, 1979). Иными словами, они принадлежат к двум разным иерархическим уровням системы, в которых изменения протекают в разном режиме и прямо не коррелируются. Именно такие соотношения между пространством путей развития и лежащей в его основе структурой генома заложены в представлении об эпигенетическом ландшафте (рис. 2, б).
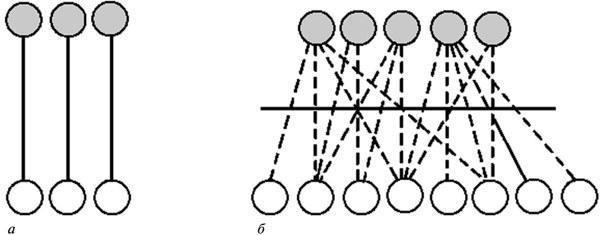
Рис. 5. Этапы оценки причинно-следственных отношений между двумя категориями факторов, наблюдаемыми в эксперименте: а — начальная гипотеза, постулирующая простые линейно-каузальные зависимости; б — обнаружение асимметрии “причин” и “следствий” (по мере расширения опытных данных), ведущее к заключению о их нелинейных отношениях и принадлежности к двум разным системным уровням.
В теоретической биологии главным источником подобных трудностей для линейно-детерминистского мышления всегда было индивидуальное развитие. Их нарастание по мере накопления опытных данных и поиска их непротиворечивого согласования неизбежно подталкивало мысль к идее о целостности механизма онтогенеза. В эмбриологии эта смена парадигмы выразилось в концепции Г. Дриша (Driesch, 1908) о развитии как “гармонической эквипотенциальной системе”, а в физиологической генетике — в затронутой выше идее Гольдшмидта о системе реагирования. Как ключевой аргумент неизменно подчеркивалось отсутствие общего соответствия между конкретным итогом развития и определенным вариантом его начального состояния.
В целом, чем содержательнее опыт науки в осмыслении своих наблюдений, тем больше она склонна отдаляться от редукционистских объяснений в сторону системных интерпретаций. И наоборот, каждый прорыв науки в незнакомую прежде область фактического знания обычно сопровождается на первом этапе возрождением механистических причинных истолкований. В истории взглядов генетики на детерминацию свойств организма подобный ход событий повторялся по меньшей мере дважды, характеризуясь сначала декларированием линейных причинных зависимостей и затем отходом от них (табл. 2). Нынешнее очередное возрождение надежд генетики на возможность редукционистского истолкования морфогенеза, связанное с открытием Нох-генов, очевидно, будет иметь ту же судьбу.
Таблица 2. “Циклическая эволюция” взглядов генетики на детерминацию развития
|
Область исследования |
Оценка причинных зависимостей |
|
|
Исходная |
Последующая |
|
|
I. Хромосомная генетика |
признаки определяются независимыми генами |
признак — продукт взаимодействия всех генов |
|
I. Молекулярная генетика |
“один ген — один фермент” |
все этапы матричного синтеза поливариантны |
|
III. Открытие гомеобокс-последовательностей |
общие Нох-гены управляют гомологичными процессами в разных организмах |
? |
В сумме можно заключить, что ограниченность субъективного отражения реальности не позволяет познавать ее иначе, как путем приближения, т.е. последовательной ревизии временных объяснительных моделей. Это продвинутое знание не может быть получено непосредственно из опыта.
Мыслители прошлого понимали это давно. По словам Галилея (1948, с. 89), если “божественный разум простым восприятием сущности... [предмета] охватывает... всю бесконечность его свойств”, то “наш способ заключается в рассуждениях и переходах от заключения к заключению”.
IV. АХИЛЛЕСОВА ПЯТА ЭВОЛЮЦИОННОГО МЫШЛЕНИЯ
Итак, самоочевидность объяснений, полученных из прямого наблюдения, есть иллюзия. На самом деле их характер всегда обусловлен обобщениями из предшествующего опыта, которые, в свою очередь, опираются на определенную концептуальную установку. Например, если для хромосомной генетики менделевское расщепление стало искомым подтверждением корпускулярной природы наследственности, то для ЭТЭ (и отчасти классического менделизма) оно выражает различие двух целостных генотипов, выявляемое в скрещивании как пороговый эффект развития (Камшилов, 1935; Шишкин, 1987). В конечном счете, понимание действующих причин явления определяется уровнем и способом нашего абстрагирования от полноты реальных взаимосвязей.
Тем не менее, в любом случае первичное представление о причинах есть необходимая ступень познавательного процесса. Как уже сказано, эта начальная объяснительная модель должна служить основой для дальнейшей коррекции и ревизии по мере ее испытания (процедуры верификации — фальсификации по К. Попперу) в свете новых наблюдений. В принципе ее исходные понятия могут и после этого сохранять свое значение как рабочий язык для описания эксперимента. Но в качестве базовых положений, обеспечивающих непротиворечивое истолкование фактов, они могут оказаться уже несостоятельными (Шишкин, 2006, с. 188).
Здесь мы переходим к проблеме, которая представляется ключевой для методологии современного эволюционизма и, в конечном счете, связана с его преимущественной опорой на генетическое мышление. Столкновение теории с закономерностями, не поддающимися в ее рамках объяснению, есть сигнал о том, что уровень упрощения реальности, принимаемый теорией, не отражает полноты нашего фактического знания. То есть принципы теории должны быть так или иначе пересмотрены. Это единственно возможный путь теоретического прогресса.
Между тем оценка главного направления эволюционной мысли последнего столетия позволяет говорить о фундаментальном нарушении этого правила. Путь, который был избран взамен, фактически можно охарактеризовать как двойственность коллективного научного сознания. Она заключается в том, что в биологии для одного и того же феномена (а именно, развития) признается два взаимоисключающих истолкования. При этом использование одного из них генетикой как основы для эволюционной теории не сопровождалось какими-либо попытками доказать несостоятельность альтернативного варианта.
Эту ситуацию можно обобщить в двух основных пунктах. (1) В редукционистской идеологии генетики механизм нормального воспроизведения (наследования) сводится к преформационной модели, где результат есть совокупность действия независимых причин. Напротив, для эмбриологии детерминация развития имеет целостную (системную) природу, не сводимую к элементарным причинам. (2) Собственный опыт генетики в изучении развития также давно вынуждает ее к формальному отказу от преформационной модели. Но именно последняя как раз и лежит в основе ГТЭ (Шишкин 1987, 2006).
То же самое противоречие обнаруживается между указанной декларативной коррекцией взглядов генетики и принципами, лежащими в основе ее реальной практики (табл. 3). Наконец, подобная же неувязка нередко имеет место и в индивидуальных представлениях авторитетов ГТЭ, отражая скрытые расхождения между собственным опытом исследователя и требованиями указанной теории (табл. 4).
Таблица 3. Расхождения между рабочими постулатами генетики (левый столбец) и положениями, признаваемыми ею декларативно или по умолчанию. Первые основаны на абсолютизации типичных экспериментальных зависимостей, вторые — на обобщениях, имеющих более широкую основу
|
Выводы, экстраполированные из гибридного анализа чистых линий |
Разделяемые генетикой положения, основанные на более широком (теоретическом и эмпирическом) знании |
|
Признаки делятся на генетически обусловленные (наследственные) и экзогенные (ненаследственные) |
Все признаки суть выражения генотипически обусловленной нормы реакции |
|
Признаки детерминируются независимыми генами |
Каждый признак определяется всем генотипом |
|
Наследование признаков подчиняется менделевским законам |
Менделевское наследование не характерно для “сырой” природной изменчивости |
Таблица 4. Примеры несоответствий между собственными обобщениями исследователя и поддерживаемой им теорией (ГТЭ)
|
Э. Майр (1968) |
Генетическая теория эволюции |
|
Эволюция начинается с поведения |
Эволюция начинается с изменений в генах |
|
Все мутации с фенотипическим проявлением вредны |
Мутации с полезным проявлением сохраняются отбором |
|
Мутации сами по себе не меняют видовой принадлежности особи |
Мутации лежат в основе видовых изменений |
Такая двойственность в оценке механизмов развития и эволюции обнажает главную проблему генетического мышления, связанную с его редукционистским пониманием феномена наследственности. Как уже не раз указывалось эмбриологами, оно фактически предполагает, что наследственность есть нечто отдельное от онтогенетического осуществления и потому может быть объяснена независимо от него, т.е. вне рамок реального знания закономерностей развития (Gurwitsch, 1912; Гурвич, 1944; Светлов, 1964). Но эти предостережения не оказали влияния на господствующую эволюционную доктрину (ГТЭ), кроме признания, что она допускает значительные упрощения. Современные попытки согласовать генетическое понимание наследственной детерминации с принципом морфогенетических полей, пришедшим из эмбриологии, остаются в пределах все тех же изначальных преформационных представлений (Шишкин, 2006, с. 193). Точно также и попытки достичь “нового эволюционного синтеза” путем объединения ГТЭ с закономерностями эмбриогенеза отнюдь не ставят своей задачей пересмотр постулатов неодарвинизма (см. раздел VI). Исходное противоречие остается неразрешенным.
В итоге важно подчеркнуть следующее. Распространение причинного объяснения за пределы тех условий опыта, в которых оно было получено, само по себе есть неизбежный шаг познавательного процесса. Но с момента, когда обнаружилась неадекватность этого объяснения для новых или расширенных условий, оно должно быть изменено. Идея мозаичной детерминации признаков, составляющая основу генетики, на деле избежала такого пересмотра, несмотря на ее очевидную непригодность для объяснения развития (ср. Светлов, 1978). Это исходное нарушение логики познавательного процесса лежит в основе всех вскрывшихся со временем главных трудностей генетической теории эволюции.
Причины такого хода событий, т.е. затянувшегося неявного сосуществования двух несовместимых концепций развития, в большой мере лежат на поверхности. Реальный отказ генетики от принципа мозаичной детерминации сделал бы по определению невозможным описание эволюции в терминах генов, составляющее основу ГТЭ. Это означало бы слишком радикальный поворот в господствующем эволюционном мышлении, чтобы можно было рассчитывать на его исторически быстрое осуществление.
Другие причины имеют в естествознании еще более давние корни. Это прежде всего традиция редукционистского понимания причинности, отдающая предпочтение механистическим объяснениям и канализирующая поиски решений в их направлении (см. выше, раздел III). И, наконец, надо указать на невысокий уровень требований теоретической биологии к согласованию своих обобщений. Последнее исторически связано с многообразием органического мира и трудностью выявления в нем универсальных закономерностей. Поэтому, в частности, типичным для эволюционизма является не стремление к созданию целостной теории, а “синтез” тех или иных факторов, понимаемых в качестве независимых участников эволюционного процесса (Шишкин, 2006, 2009).
V. ИЛЛЮЗИИ ЭВОЛЮЦИОННЫХ РЕКОНСТРУКЦИЙ
Абсолютизация зависимостей, полученных непосредственно из эксперимента или сравнения, без оценки их реальной значимости в свете других биологических обобщений, составляет типичную практику истолкования эволюционных процессов на основе опытного знания. Этот подход характерен как для эмбриологии, так и для генетики (Шишкин, 2006; 2007, с. 180).
Предметом изучения для обеих названных дисциплин является пространство вариантов развития и контролирующие их механизмы. И обе они чаще всего (в случае генетики — всегда) судят об эволюционном значении наблюдаемых явлений, опираясь по умолчанию на две презумпции. (1) Исторические изменения протекают в том же направлении, что и онтогенетические (от ранних стадий развития к поздним); поэтому вторые могут быть прямым отражением первых. (2) Связи между взрослыми структурами и их причинами в раннем развитии или зародышевой клетке могут быть упрощены до линейных отношений, т.е. до преформационной модели.
Как уже сказано, второе из этих допущений в теории отвергается обеими дисциплинами, т.е. опора на него заведомо нелогична. Что касается допущения (1), то в эмбриологии оно не имеет специального обоснования (кроме лишь расширительного толкования биогенетического закона) и принимается скорее эмпирически — как простейшее из возможных (Шишкин, 2007). Напротив, для генетики это допущение прямо вытекает из ее идеологии, принимающей, что свойства организма детерминируются генами, а изменения этих свойств — мутациями генов. Это обстоятельство, вместе с фактом, что именно генетика является основой господствующих эволюционных представлений, определяет наше главное внимание к практике исторических реконструкций на основе генетических данных.
Как уже говорилось (см. Введение), генетическому пониманию эволюции противостоит позиция ЭТЭ, согласно которой эволюционные изменения распространяются в поколениях от взрослой стадии к началу развития. Иными словами, согласно данной теории, изменения проявляются раньше, чем формируется организация зародышевой клетки, обеспечивающая их стабильное воспроизведение (рис. 4, б: F1—F4). Если ГТЭ прямо проецирует на эволюцию результаты экспериментов, определяющих в ее понимании связи генов и признаков (в классическом случае — на основе гибридного анализа), то позиция ЭТЭ основана на осмыслении общего разнообразия эмпирических зависимостей, наблюдаемых в онтогенезе и не укладывающихся как целое в линейные схемы. Различия этих двух подходов играют решающую роль в оценке возможностей эволюционных реконструкций.
Выше подчеркивалось, что при сопоставлении хода развития в разных условиях эксперимента вскрывается общая асимметрия причин и следствий (рис. 1; 2, б; 5, б). И что особенно важно, обнаруживается ограниченность пространства вторых по отношению к разнообразию первых. Главный вывод из этого феномена (особенно в применении к нормальному фенотипу), известный еще с конца XIX в., сводится к правилу: “результат развития более устойчив, чем способ его осуществления”. Этот вывод заключен в обобщениях, известных в эмбриологии как “правило Ру”, принцип эквифинальности Дриша, принцип нормировки Гурвича, а также во множестве обобщений, обозначающих эволюционное сжатие и упрощение развития при сохранении его прежнего результата, выявляемые сравнительным путем (Шишкин, 2006, с. 182). В генетике эта же закономерность выражается в правиле Четверикова (генетическая гетерогенность под покровом “дикого” фенотипа), в представлениях Гольдшмидта о системе развития, в факте гетерогенности однотипных природных аберраций и в бесчисленных примерах реализации одних и тех же свойств на разной генетической основе (см., например, Шмальгаузен, 1982; Дубинин, 1966). Одним из впечатляющих примеров этого правила является детерминация пола, реализуемая в органическом мире на основе самых различных механизмов — от мужской или женской гетерогаметности до определения факторами среды, причем иногда разными способами у предельно близких таксонов.
В результате, даже безотносительно к постулатам ЭТЭ, весь опыт экспериментального знания о развитии, учитываемый этой теорией, может быть обобщен в виде одного базового принципа, проливающего свет на эволюционную оценку событий онтогенеза. А именно, что программа развития, заключенная в зародышевой клетке, всегда исторически моложе, чем дефинитивный результат, реализуемый этой программой. Последняя характеризует лишь один из вариантов такой реализации, наряду со многими другими — как нередко существующими параллельно, так и необходимо предшествовавшими данному варианту в ходе его становления. Другими словами, история каждой новой элементарной особенности начинается с ее появления на основе несовершенного морфогенеза, который в дальнейшем преобразуется все более глубоко. (В представлениях ЭТЭ этот процесс описывается как стабилизация новшества; см. рис. 4, б: F1—F4). Повторим, что такой взгляд на эволюцию хода развития есть не только одно из ключевых положений ЭТЭ, но и вывод из общей оценки свойств онтогенеза как системы. По заключению эмбриолога Л.Б. Белоусова (1979) эволюция есть усиление и распространение на нижние системные уровни тех изменений (“неустойчивостей”) онтогенеза, которые были изначально предопределены на верхнем (морфологическом) уровне.
Из сказанного понятно, что искать в функциональных свойствах генома или зародышевой клетки прямых указаний на способ исторического возникновения реализуемых ими взрослых особенностей есть предельно наивное ожидание. На деле все, что мы наблюдаем в современном онтогенезе, создающем данный взрослый план строения, есть максимально трансформированная (вплоть до зиготы) модель его развития, прошедшая долгий путь стабилизации. Она далеко отстоит от первоначальной модели (ср. рис. 4, б: F1, F4), которая была относительно неустойчивой, слабо интегрированной (лишенной многих корреляций, выявляемых в современном онтогенезе) и зависимой от внешних факторов. Поэтому особенности развития любой структуры, регистрируемые экспериментатором, являются продуктом всей истории ее существования, а не указанием на механизм ее появления в эволюции. Допускать обратное значит принимать следствия за причины. В действительности, говоря словами С.Г. Крыжановского (1939), развитие непосредственно не содержит в себе метода для своей исторической интерпретации.
Между тем, практически все суждения о природе эволюционных новшеств, исходящие из данных генетики, основаны именно на указанном отождествлении причин и следствий исторического процесса. Таковы, прежде всего, бесчисленные гипотезы о происхождении различных фенотипических свойств за счет эффектов мутаций, понимаемых как одномоментные устойчивые изменения фенотипа (например, в случае меланизма или альбинизма млекопитающих, редукции крыльев у насекомых, торсионного процесса у брюхоногих моллюсков и т.д.). Эти гипотезы, не предусматривающие никакой исторической смены начальных факторов развития по отношению к его конкретному результату (ср. рис. 4, а), неизбежно подразумевают и другие несостоятельные допущения. Во-первых, они опираются на преформационную модель развития, перенося ее механически далее на эволюцию. Во-вторых (в соответствии с первым допущением), они исходят из убеждения, что “генетическая основа” признака, выявляемая гибридным анализом, это и есть его исторический “детерминирующий фактор”. На самом же деле речь идет лишь о пороговой функции этого фактора (при переключении родительских онтогенетических траекторий у гибридов), возникающей исключительно в данной комбинации скрещивания (подробнее см. Шишкин, 1987, с. 95-98, 101).
Такое же понимание эволюционного процесса заключено и в более сложных реконструкциях, например, при освещении эволюции насекомых от предков, близких к многоножкам, в направлении двукрылых (Lewis, 1985; Рэфф, Кофмен, 1986). Эта трансформация описывается как результат последовательных мутаций (или дупликаций с последующей дивергенцией функций генов), приведших сначала к утрате конечностей в абдоминальных сегментах у ранних крылатых насекомых, а позднее (у Diptera) также и к редукции крыльев заднегруди. Вся схема построена на анализе гомеозисных переключений в имагинальных дисках у высших насекомых при мутациях или последовательных делециях в соответствующих генных комплексах. То есть речь идет об особенностях исторически глубоко трансформированного типа морфогенеза. Далее они экстраполируются на ранние этапы эволюции насекомых, на которых не было ни развития с полным превращением, ни свойственного ему упреждающего обособления зачатков взрослых тканей (имагинальных дисков), ни соответственно, высоко интегрированных формативных зависимостей, выявляемых как альтернативные программы развития этих зачатков. Таким образом, формообразовательные гомеозисные реакции, характеризующие высших насекомых, в данной реконструкции событий (а) приписывается специфическим генам, (б) переносится на примитивные типы развития, не имевшие таких свойств, и (в) отождествляется непосредственно с эволюционными механизмами. Это еще один пример смешения причин и следствий эволюционного процесса, основанный на представлении, что причинные связи хода развития, наблюдаемые в эксперименте, и эволюция — это в принципе одно и то же (ср. рис. 4, а: A—D).
В конечном счете, вся логика умозаключений в этих реконструкциях объективно исходит из убеждения, что мозаичная детерминированность развития есть единственно возможный вывод из данных генетики — независимо от того, насколько он совместим с обобщениями эмбриологии.
VI. СИМПТОМЫ КРИЗИСА ТЕОРИИ
Вернемся к вопросу о факторах, определяющих утрату теорией статуса достоверного объяснения реальности. Используя снова эволюционное учение как модельный объект, мы приходим к отчасти неожиданному выводу, что для такого хода событий вообще не требуется появления противоборствующей альтернативной концепции, по крайней мере, в оформленном виде.
Любая модель функционирования, свойственная живым организмам, включая и человеческую деятельность, предполагает определенную систему отражения реальности, с которой сообразуются все выполняемые действия. Научная теория — такая же система, организующая познавательный процесс, независимо от того, сознает ли это ее носитель. Обычно она включает небольшое число исходных допущений. Ценность ее для познания тем выше, чем более широкий круг явлений она позволяет объяснить на этой основе, показывая их общие причинные связи. Таким образом, содержательная теория максимально экономна (рис. 6, б). Она не заменяет опытного знания, но сильно облегчает выбор возможных для него направлений поиска, отсекая пути, заведомо бесперспективные в свете ее представлений. В этом суть известного обобщения, что нет более практичной вещи, чем хорошая теория.
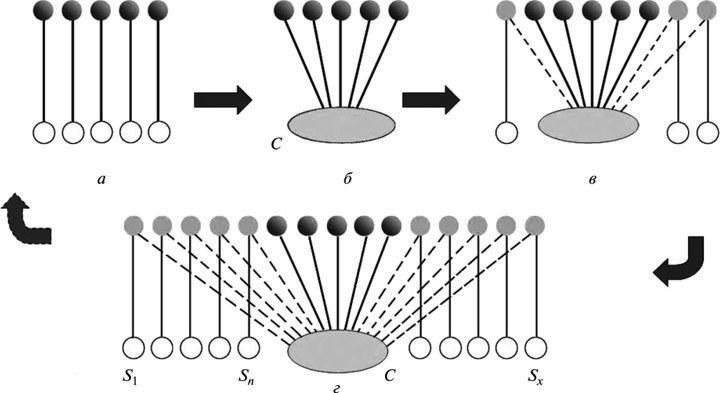
Рис. 6. Симптомы кризиса теории: а — исходный этап познания (отсутствие общего объяснения для наблюдаемых явлений); б — появление теории (переход к истолкованию фактов на единой основе); в — обнаружение фактов, не объяснимых для теории без дополнительных частных допущений; г — нарастание ситуации, возникшей на этапе (в). Объяснительные возможности теории сокращаются, приближая ее к состоянию, существовавшему на этапе (а). Обозначения: С — концептуальная основа теории; S1, Sn, Sx, — дополнительные допущения.
Прогресс в сфере знаний, к которой относится теория, может быть стимулом к ее совершенствованию и конкретизации ее понятий. Например, для неодарвинизма таким шагом было открытие хромосом и их комбинирования вместе с мутационным процессом, а для классического дарвинизма (с большим запозданием) — открытие системных свойств онтогенеза. Эти изменения могут сильно менять язык теории, но не задевают ее постулатов и не нарушают ее объяснительной логики.
Положение становится иным, когда теория сталкивается с фактами и обобщениями, противоречащими ее ожиданиям. Осознание ею таких препятствий не означает, что они не были известны науке задолго до этого. Но способ их восприятия, если говорить о биологии, меняется во времени и зависит от многих причин. Во-первых, потребность в согласовании обобщений, полученных в разных областях биологического знания, мало свойственна коллективному научному сознанию (Шишкин, 2006; ср. выше, раздел IV) — до тех пор, пока она не становится самоочевидной. Во-вторых, в своей оценке “неудобных” фактов теоретическое мышление склонно руководствоваться не столько общими требованиями логики, сколько инерцией своего концептуального выбора, подразумевающего правоту и незыблемость защищаемой теории. Очевидно, по этим причинам радикальное противоречие между принципом мозаичной детерминации и свидетельствами целостности развития оставалось на долгие годы вне поля зрения неодарвинизма (ГТЭ) в после-вейсмановскую эпоху. В ином случае это учение никогда не смогло бы стать доминирующей доктриной в биологии ХХ в. Историки науки называют и более конкретную причину такого хода событий (по сути включающую обе предыдущие). Они указывают, что идея морфогенетического поля, определяющего ход развития, отвергалось моргановской генетикой в качестве опасной альтернативы представлению о гене как факторе детерминации (Гилберт и др., 1997, с. 327—330).
Оценка адекватности ГТЭ стала меняться в коллективном сознании лишь по мере того, как детальный анализ механизмов развития (как в отношении продуктов синтеза, так и морфологических особенностей) стал одной из главных задач генетики. Как и следовало ожидать, он принес новые подтверждения известного правила, что соотношения между причинами и их результатами в развитии неоднозначны на всех его уровнях, начиная с экспрессии генов (ср. табл. 2). Понятно, что для теории, сводящей объяснение эволюции к изменению генного состава популяций, это означает неразрешимые проблемы. Указанная неопределенность стала стимулом к поискам “нового синтеза”, в котором нашлось бы место для причинных соотношений и механизмов, вскрываемых эмбриологией.
Главная трудность обрисовывалась инициаторами этой новой программы следующим образом. Чтобы понять связь между геномными изменениями и эволюцией, необходимо знать, как структура генома преобразуется в определенную морфологию. Но здесь между сигналом и ответом существует “черный ящик”. Процессы, изоморфные геному, заканчиваются на определении первичной структуры белков и находятся далеко от процессов развития, формирующих организм (Evolution..., 1982). Следовательно, объяснения этих последних нужно искать на других путях, т.е. обратившись к эмбриологическому пониманию закономерностей морфогенеза.
Этот план реорганизации теории фактически возлагает надежды на ее усовершенствование за счет альтернативного ей принципа (системное объяснение развития), без пересмотра ее прежней основы. Собственно новый синтез обычно мыслится как “сложение” этих разнородных предпосылок. Принимается, что сегодняшняя ГТЭ по-прежнему адекватна для истолкования микроэво-люционных процессов, но что изменения макро-эволюционного уровня должны описываться на основе знания законов онтогенеза (Gould, 1982; Maderson et al., 1982). Такой же подход заложен и в попытках построения концепции развития на основе синтеза представлений генетики и эмбриологии — как ряда этапов каскадной экспрессии генов в ходе ступенчатой детерминации. Он сводится к идее чередования активности генов и морфогенетических полей, контролируемых генами, причем вся последовательность событий должна инициироваться генами высшего иерархического порядка (Гилберт и др., 1997). По сути это лишь сильно усложненный вариант префор-мационной модели развития, в которой действие системных факторов играет сугубо подчиненную роль (Шишкин, 2006). Суммируя, речь идет о попытках восстановить объяснительную ценность генетической теории путем включения в нее чужеродных принципов, сфера действия которых ограничивается произвольным образом.
Трудности генетического объяснения эволюции не только прямо сфокусированы в его ключевом постулате (отбор мозаичных детерминантов развития), но имеют и множество других связанных с ним проявлений. Многие из них до сих пор не усвоены коллективным сознанием, хотя, казалось бы, лежат на поверхности. Так, не осознается, что генетическая (“эволюционно значимая”) обусловленность признаков, приписываемая неодарвинизмом действию мутаций, есть на самом деле свойство любой реализуемой особенности, поскольку все они суть выражения нормы реакции (табл. 3; ср. Шишкин, 2006, с. 183, 188). Или, что эффект менделевского наследования, наблюдаемый в гибридном анализе, создается собственными усилиями исследователя, стабилизирующего сначала чистые родительские линии. И потому видеть в этом феномене фактор эволюции, управляющий природной изменчивостью, значит принимать продукт отбора за его сырой материал (Шишкин, 1987, с. 117).
Там, где такие трудности очевидны для теории, она пробует объяснить их с помощью дополнительных допущений ad hoc (ср. табл. 1). Но эти попытки лишь обозначают проблему, не позволяя ее устранить. Если, например, эффекты мутаций всегда наследуются хуже, чем норма, то, каким бы путем это ни объяснять, они заведомо не обладают той исторической устойчивостью, которую им приписывает неодарвинизм. Или, если соотношения генома и фенотипа принципиально неоднозначны, то непонятно, как в этом случае можно объяснить механизм отбора мутаций, на котором зиждется указанная теория.
Схематически весь рассмотренный ход событий в истории существования теории описывается следующим образом. Чем больше растет область фактов, трудно с нею совместимых, тем шире она вынуждена прибегать к объяснениям, требующим частных дополнительных гипотез (ср. табл. 1). В результате “объяснительная структура” теории все более приближается к тому состоянию, которое предшествовало ее появлению (рис. 6, а). Иными словами, ранний этап, на котором предлагался общий принципиальный ответ на множество разнородных вопросов (рис. 6, б), сменяется этапами, на которых нарастает симметрия между числом вопросов, возникающих перед теорией, и числом предлагаемых ответов (рис. 6, в, г). В итоге эта объяснительная схема как целое перестает быть экономной и прогностич-ной, т.е. все более утрачивает статус теории. Пытаясь защититься, теория разрушает самое себя.
Если в эволюции теории обозначился такой ход событий, то его финал предсказуем независимо от того, существует ли для данной теории сложившаяся альтернатива. Последняя все равно должна рано или поздно возникнуть в поисках ответа на те вопросы, для которых не нашлось объяснения в рамках существующих представлений. В случае развития идеи эволюции на основе естественного отбора так оно фактически и происходит. Хотя системная альтернатива неодарвинизму в виде эпигенетической теории Шмальгаузена—Уоддингтона (ЭТЭ) существует уже более полувека, ее влияние на коллективную эволюционную мысль было достаточно ограниченным в силу многих причин (ср. Шишкин, 2006, с. 184). Тем не менее, неудовлетворенность сторонников ГТЭ ее нынешним состоянием стимулирует поиски нового, более широкого “синтетического” обобщения на тех же самых путях, на которых сложилась ЭТЭ. Несмотря на оппортунистический характер большинства таких попыток, стремящихся сохранить в новой теории центральную догму неодарвинизма, само их направление показательно. Оно говорит в пользу конечной неизбежности признания принципов, лежащих в основе ЭТЭ.
ЗАКЛЮЧЕНИЕ (ПЕРСПЕКТИВЫ ЭВОЛЮЦИОНИЗМА)
Из сказанного понятен наиболее вероятный прогноз в отношении дальнейшего развития эволюционной теории. Он основан на убеждении, что при всей своей традиционной мозаичности, теоретическое мышление в биологии все же является единой системой с внутренними обратными связями. Поэтому оно в конечном счете способно к постепенной самокоррекции, основанной на критическом сопоставлении и согласовании собственных обобщений. Этот процесс неизбежно должен привести коллективную эволюционную мысль к осознанию несовместимости системных свойств развития с принципом корпускулярной наследственности. В соответствии с этим завтрашняя эволюционная теория будет рассматривать смену органических форм как преобразование целостных систем развития. Все свойства развития, вскрываемые эмбриологией и генетикой, будут интерпретироваться в этом контексте. Попытки фетишизации каких-либо структур живого в качестве самостоятельных факторов эволюции, возникающих и действующих помимо естественного отбора, останутся для теории в прошлом.
СПИСОК ЛИТЕРАТУРЫ
Белоусов Л.В. Целостные и структурно-динамические подходы к онтогенезу // Журн. общ. биол. 1979. Т. 40. № 4. С. 514-529.
Галилей Г. Диалог о двух главнейших системах мира — Птоломеевой и Коперниковой. М.-Л.: ОГИЗ — Гос. изд-во Техн.-теор. лит., 1948. 379 с.
Гилберт С.Ф., Опиц Д.М., Рэф Р.А. Новый синтез эволюционной биологии и биологии развития // Онтогенез. 1997. Т. 28. № 5. С. 325—343.
Гурвич А.Г. Теория биологического поля. М.: Сов. наука, 1944. 156 с.
Дарвин Ч. Происхождение видов путем естественного отбора. М.: Тайдекс Ко, 2003. 495 с.
Дубинин Н.П. Эволюция популяций и радиация. М.: Атомиздат, 1966. 743 с.
Камшилов М.М. Является ли плейотропия свойством гена? // Биол. журн. 1935. Т. 4. № 1. С. 113—144. Крыжановский С.Г. Принцип рекапитуляции и условия исторического понимания развития // Памяти акад. А.Н. Северцова. М.; Л.: Изд-во АН сСсР, 1939. С. 281—366.
Любищев А.А. О природе наследственных факторов // Изв. Биол. научн.-исслед. ин-та при Пермском университете. 1925. Т. 4. Прилож. 1. С. 1—142.
Маяковский В. Стихи о разнице вкусов // Собр. соч. в 3х томах. Т. 2 (1928). М.: Худ. лит-ра, 1978. С. 358.
Майр Э. Зоологический вид и эволюция. М.: Мир, 1968. 597 с.
Рэфф Р., Кофмен Т. Эмбрионы, гены и эволюция. М.: Мир, 1986. 402 с.
Светлов П.Г. О целостном и элементаристическом методах в эмбриологии // Арх. анатомии, гистологии и эмбриологии. 1964. Т. 46. № 4. С. 3—26.
Светлов П.Г. Физиология (механика) развития. Л.: Наука, 1978. Т. 1. 279 с.; Т. 2. 262 с.
Филипченко Ю.А. Эволюционная идея в биологии. М.: Наука, 1977. 227 с.
Шишкин М.А. Онтогенез и эволюционная теория // Эволюция и биоценотические кризисы / Ред. Л.П. Та-таринов, А.П. Расницын. М.: Наука, 1987. С. 76—123.
Шишкин М.А. Закономерности эволюции онтогенеза // Современная палеонтология / Ред. В.В. Меннер, В.П. Макридин. Т. 2. М.: Недра, 1988. C. 169-209.
Шишкин М.А. Индивидуальное развитие и уроки эволюционизма // Онтогенез. 2006. Т. 37. № 3. 179-198.
Шишкин М.А. Морфогенез как объект исторического прочтения: продукт эволюции или ее запись? // Клеточные, молекулярные и эволюционные аспекты морфогенеза. Симпозиум с международным участием / Ред. Л.В. Белоусов. М.: КМК, 2007. C. 179-183.
Шишкин М.А. Эволюционная теория и особенности научного мышления // Научное наследие Шмальгаузена и его развитие. Конф. посв. 125-летию со дня рождения акад. И.И. Шмальгаузена. М.: ИПЭЭ РАН им. А.Н. Северцова, 2009. C. 46-50.
Шмальгаузен И.И. Пути и закономерности эволюционного процесса. М.-Л.: Изд-во АН СССР, 1940. 231 с.
Шмальгаузен И.И. Стабилизирующий отбор и его место среди факторов эволюции // Журн. общ. биол. 1941. Т. 2. № 3. С. 307-354.
Шмальгаузен И.И. Организм как целое в индивидуальном и историческом развитии. М.: Наука, 1982. 228 с.
Bertalanffy L. General system theory. N.Y.: Braziller, 1969. 289 p.
Driesch H. The science and philosophy of the organism. L.: Black, 1908. V. 1. 329 p.; V. 2. 381 p.
Evolution and development // Report of the Dahlem Workshop / Ed. J.T Bonner. Berlin, Heidelberg, N.Y.: Springer, 1982. 357 p.
Goldschmidt R. Physiological genetics. N.Y.: L. McGraw Hill Book Co., 1938. 375 p.
Goldschmidt R. The material basis of evolution. New Haven: Yale Univ. Press, 1940. 436 p.
Gould S.J. Ontogeny and phylogeny. Cambridge: Belknap Press, 1977. 501 p.
Gould S.J. The meaning of punctuated equilibrium and its role in validating a hierarchical approach to macroevolution // Perspectives in evolution / Ed. R. Milkman. Sunderland, MA: Sinauer, 1982. P. 83-104.
Gurwitsch A. Die Vsrerbung als Vsrwirklichungvorgang // Biol. Zbl. 1912. Bd 32. S. 458-486.
Lewis E.B. Regulation of the genes of the bithorax complex in Drosophila // Cold Spring Harbor Symp. Quant. Biol. 1985. У 50. P. 155-164.
Maderson P.F.A., Alberch P., Goodwin B.C. et al. The role of development in macroevolutionary change // Evolution and development. Dahlem Konferenzen / Ed. J.T. Bonner. Berlin: Springer, 1982. P. 279-312.
Waddington С.Н. The strategy of the genes: a discussion on some aspects of theoretical biology. L.: Allen and Unwin, 1957. 262 p.
Evolutionary Theory and Scientific Thinking M. A. Shishkin
In natural sciences, the advance of evolutionary thought and growth of empirical knowledge are not strictly correlated. The state of theory primarily tends to be controlled by a mode of collective thinking that historically dominates a given branch of science. This particularly holds true for the natural selection concept, which has two alternative interpretations known as the genetic and epigenetic theories of evolution. The final result of their competition, albeit predictable, will not be based upon any kind of “crucial evidence” giving advantage to either of them. The above result will be in fact attained as soon as the evolutionary biology can overcome the tradition of mosaic thinking which enables the incompatible concepts to be combined. In this respect, the key point to be realized is that the idea of corpuscular determination of the ontogeny is incompatible with understanding the development as a systemically controlled process.
Key words: evolution, genetics, epigenetic theory, ontogeny.
1Такой же путь эволюции постулируется для провизорных адаптаций, реализуемых в ходе развития (Шишкин, 1988, с. 183-186).Обратно к тексту
Коментарі
Как хорошо, что это появилось на Батрахосе!
Удивительно, но последние статьи М.А. Шишкина, известного громоздкостью и нечитабельностью иных своих текстов, очень ясны и привлекательны для чтения. Я бы даже сказал, что эти статьи логичнее и проще для понимания, чем "научно-популярные" заметки другого автора на букву "Ш", часто пишущего на сходные темы.
Эмоционально статья очень убедительна - особенно в разделах V и VI. Хочется согласиться буквально с каждым словом. Обобщения прекрасны и будоражат ум. Всё на своём месте. Но анализа по-настоящему свежих данных эмбриологии и генетики здесь, конечно, недостаёт. Вот прям хотелось бы их тут побольше - а нету. Все ссылки - на что-нибудь, опубликованное, в лучшем случае, лет 15 назад. Это вполне понятно и извиняемо, но на научной (не эмоциональной) убедительности отражается плохо. Не зря сквозь общий оптимистический тон статьи таки просматривается признание того, что в ближайшем времени ЭТЭ широкого признания не получит. "Рано или поздно ..." - пишет Шишкин - "... в конечном итоге ..." Ну, да.
Не пнуть - не мог!
...да еще с прибауточками...
Товарищу на букву Ш.
Товарищу на букву Ш. досталась непростая роль популяризатора. Не будем его судить строго. Если люди сюда пришли и почитали, значит он делает свою работу неплохо)
Шишкин - один из умнейших
Шишкин - один из умнейших людей нашего времени, после Красилова. А может даже и перед.
А вот комментаторы в ЖЖ удивляют. Нет бы пройти и ознакомиться. Не хотят. Увы.