Українська мова (найновіша версія) / Русский язык (обновление прекращено)
4.15. Экологические стратегии
Как определить ценность особи для популяции?
«Естественный отбор признает только один вид «валюты» — благополучное потомство» (Э. Пианка, 1981).
Мы говорили, что популяция — потенциально бессмертная сущность, состоящая из смертных особей. Чтобы поддержать существование популяции, особь должна выжить сама и оставить потомков, которые тоже смогут выжить. Обратите внимание на двойственность этой задачи. Вероятно, наибольшие шансы на выживание будет иметь та особь, которая вообще не будет тратить ресурсы и полученную из них энергию на производство потомства. Но пройдет немного времени — и такая особь без следа исчезнет из популяции. На противоположном «полюсе» находится гипотетическая особь, которая сразу после своего появления начинает всю свою энергию направлять на производство потомков. Такое существо погибнет само и, если его потомки унаследуют столь же неэффективный способ распределения ресурсов, произведет потомков, которые не будут иметь шансов на выживание.
Значит, наибольшую ценность для популяции должна иметь особь, сочетающая затраты на собственное выживание и на производство потомков в оптимальном сочетании. Оценить, насколько это сочетание оптимально, можно. Для этого нужно высчитать, при каком сочетании в данных условиях особь оставит наибольший возможный вклад в будущее поколение. Мера, которая используется для этого в математической популяционной биологии, называется репродуктивной ценностью. Репродуктивная ценность — обобщенная мера выживаемости и плодовитости, учитывающая относительный вклад организма в будущие поколения.
«Легко описать гипотетический организм, имеющий все признаки, необходимые для достижения высокой репродуктивной ценности. Он размножается почти сразу же после рождения, дает многочисленное, крупное, защищенное потомство, о котором заботится; он размножается многократно и часто на протяжении долгой жизни; он побеждает в конкурентной борьбе, избегает хищников и легко добывает пищу. Описать такое существо легко, но представить трудно...» (Бигон и др., 1989).
Вы понимаете, что такая невозможность вытекает из противоречивости задач самоподдержания и размножения (рис. 4.15.1). Одним из первых это осознал в 1870 г. английский философ Герберт Спенсер, говоривший об альтернативности поддержания организмом собственного существования и продолжения себя в потомках. На современном языке можно сказать, что эти параметры связаны отрицательными корреляциями, отношением, при котором улучшение системы по одному параметру должно сопровождаться ее ухудшением по другому.
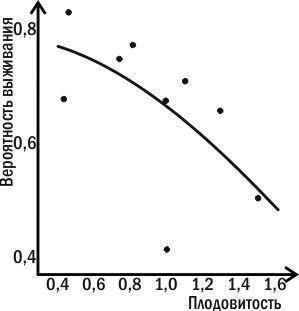
Рис. 4.15.1. У коловратки Asplanchna шансы на выживание уменьшаются по мере роста плодовитости (Пианка, 1981)
Разные виды (и разные популяции) по-разному перераспределяют энергию между самоподдержанием и размножением. Можно говорить о видовой стратегии, выражающейся в том, как представители вида добывают ресурсы и как они их тратят. Успешной может быть только та стратегия, при которой особи получают достаточное количество энергии, чтобы они могли расти, размножаться и компенсировать все потери на активность хищников и разнообразные несчастья.
Признаки, относящиеся к разным адаптивным стратегиям, могут быть связаны отношением трейдоффа, то есть непреодолимыми отрицательными корреляциями (отношением или-или). Так, отношением трейдоффа связаны число потомков и их выживаемость, скорость роста и устойчивость к стрессу и т.д. Американские экологи Р. Мак-Артур и Е. Уилсон описали в 1967 году два типа видовых стратегий, которые являются результатом двух разных типов отбора и связаны отношением трейдоффа. Принятые обозначения этих стратегий (r- и K-) взяты из логистического уравнения.
Согласно логистической модели, в росте популяции можно выделить две фазы: с ускоряющимся и с замедляющимся ростом (рис. 4.15.2). Пока N невелико, на прирост популяции основное влияние оказывает сомножитель rN и рост популяции является ускоряющимся. На этой фазе (r-фазе) рост популяции ускоряющийся, и ее численность тем выше, чем выше способность особей к размножению. Когда N становится достаточно высоким, на численность популяции начинает оказывать основное влияние сомножитель (K-N)/K. На этой фазе (К-фазе) рост популяции замедляющийся. Когда N=K, (K-N)/K=0 и рост численности популяции прекращается. На K-фазе численность популяции тем выше, чем выше параметр K. Он тем выше, чем более конкурентоспособны особи.
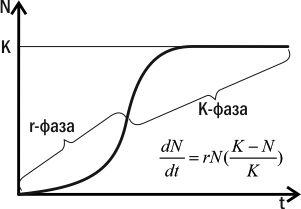
Рис. 4.15.2. r- и K-фазы популяционного роста в соответствии с логистической моделью
Можно предположить, что популяции некоторых видов подавляющую часть времени находятся на r-фазе. У таких видов максимальную репродуктивную ценность имеют особи, способные быстро размножаться и захватывать пустующую среду своими потомками. Иными словами, на этой фазе отбор будет способствовать повышению параметра r — р епродуктивного потенциала. Такой отбор называется r-отбором, а возникающие в его результате виды —г-стратегами.
У видов, популяции которых подавляющую часть времени находятся на K-фазе, ситуация совсем иная. Максимальная репродуктивная ценность в этих популяциях будет присуща особям, которые окажутся настолько конкурентоспособными, что смогут заполучить свою долю ресурса даже в условиях его недостатка; только в этом случае они смогут размножиться и сделать свой вклад в следующее поколение. Популяция, состоящая из таких особей, будет иметь более высокое значение параметра K — емкости среды, чем такая, которая состоит из особей, не «умеющих» бороться за недостающие ресурсы. На этой стадии на популяцию действует К-отбор, результатом которого является появление видов — К-стратегов. K-отбор направлен на увеличение затрат на развитие каждой особи и повышение ее конкурентоспособности.
Между этими стратегиями возможны переходы, но они носят промежуточный характер, а не объединяют типичные выражения двух форм.
«Нельзя быть одновременно салатом и кактусом» (Э. Пианка).
Важное значение для определения того, какой отбор (r- или K-) будет действовать на вид, имеет динамика изменения количества доступного ресурса и острота конкуренции за него. При резком неизбирательном сокращении численности популяций, вызванным обусловленным внешними причинами недостатком ресурса, преимущество получают r-стратеги, а при конкурентной борьбе за недостающий ресурс — K-стратеги.
Выбор между r-стратегией (повышением плодовитости) и K-стратегией (повышением конкурентоспособности) представляется достаточно простым, однако он затрагивает множество параметров организмов и их жизненных циклов. Сравним эти стратегии в их типичной форме (табл. 4.15.1).
Таблица 4.15.1. Особенности r- и K- отбора и стратегий
|
Характеристики |
r-отбор и r-стратеги |
K-отбор и K-стратеги |
|
Климат |
Изменчивый, непредсказуемый |
Постоянный, предсказуемый |
|
Смертность |
Катастрофическая, независимая от плотности популяции |
Вызванная конкуренцией, зависящая от плотности популяции |
|
Кривая смертности |
Обычно типа III |
Обычно типа I или II |
|
Размер популяций |
Изменчивый, неравновесный |
Постоянный, близкий к предельной емкости среды |
|
Свободные ресурсы |
Появление свободных ресурсов, за¬полнение «экологического вакуума» |
Свободных ресурсов почти не бывает, они заняты конкурентами |
|
Внутри- и межвидовая конкуренция |
Слабая |
Острая |
|
Размер тела |
Относительно мелкий |
Относительно крупный |
|
Развитие |
Быстрое |
Медленное |
|
Половозрелость |
Ранняя |
Поздняя |
|
Скорость размножения |
Высокая |
Низкая |
|
Размножение в течение жизни |
Часто однократное |
Неоднократное |
|
Потомков в выводке |
Много |
Мало, часто один |
|
Количество ресурса на одного потомка |
Низкое |
Высокое |
|
Продолжительность жизни |
Короткая |
Долгая |
|
Приспособления |
Примитивные |
Совершенные |
|
Оптимизируется |
Продуктивность |
Эффективность |
Может удивить, почему для r-стратегов характерно однократное размножение, а для K-стратегов — неоднократное. Эту особенность проще пояснить примером. Представьте себе мышей, заселяющих амбар с зерном (ресурса вдоволь, конкуренции никакой). Рассмотрим стратегии двух видов.
Вид № 1. Половозрелость в 3 месяца, количество потомков в выводке — 10, самка живет год и способна размножаться каждые три месяца.
Вид № 2. Половозрелость в 3 месяца, количество потомков в выводке — 15, выкормив их, самка гибнет от истощения.
В первом случае через три месяца к размножению приступит 10 потомков и их родители (всего 12 голов), а во втором — целых 15 потомков. Более высокую скорость захвата свободных ресурсов сможет обеспечить второй вид. Типичная r-стратегия заставляет особей выкладываться в размножении как можно раньше и как можно сильнее, и поэтому r-стратеги зачастую ограничены одним-единственным сезоном размножения.
С другой стороны, легко понять, почему типичные K-стратеги размножаются много раз. В конкурентной среде выживет только тот потомок, на развитие которого потрачено много ресурса. С другой стороны, чтобы выжить и размножаться, взрослая особь должна тратить значительное количество энергии на собственное поддержание и развитие. Поэтому в предельном случае K-стратеги приносят по одному потомку за один раз (как, например, слоны и киты, а также, в большинстве случаев, и люди). Но как бы ни были совершенны эти животные, пара родителей со временем погибнет. Чтобы популяция не пресеклась, пара родителей должна оставить пару выживших потомков, а родить, следовательно, должна более двух. Раз так, необходимым условием выживания K-стратегов оказывается многократность размножения составляющих их особей.
В 1935 г. советский ботаник Л.Г. Раменский выделил три группы растений, которые он назвал ценотипами (понятие о стратегиях еще не было сформировано): виоленты, патиенты и эксплеренты. В 1979 году их эти же группы (под другими названиями) открыл заново английский эколог Дж. Грайм (рис. 4.15.3). Эти стратегии таковы.
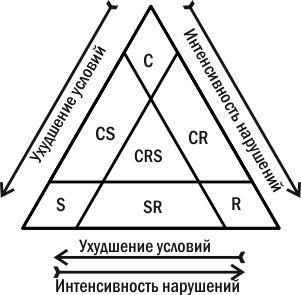
Рис. 4.15.3. «Треугольник Грайма» — классификация видовых стратегий
— Тип C (competitor, конкурент), виолент по Раменскому; затрачивает большую часть энергии на поддержание жизни взрослых организмов, доминирует в устойчивых сообществах. Среди растений к этому типу чаще всего относятся деревья, кустарники или мощные травы (например, дуб, тростник).
— Тип S (stress-tolerant, стресс-толерант); патиент по Раменскому; благодаря специальным адаптациям выносит неблагоприятные условия; использует ресурсы там, где с ним за них почти никто не конкурирует. Обычно это медленно растущие организмы (например, сфагнум, лишайники).
— Тип R (от лат. ruderis, рудерал), эксплерент по Раменскому; замещает виолентов в разрушенных сообществах или использует временно невостребованные другими видами ресурсы. Среди растений это однолетники или двухлетники, которые производят множество семян. Такие семена образуют банк семян в почве или способны эффективно распространяться на значительное расстояние (например, одуванчик, иван-чай). Это позволяет таким растениям дожидаться момента высвобождения ресурсов или вовремя захватывать свободные участки.
Многие виды способны совмещать разные типы стратегий. Сосна относится к категории CS, так как она хорошо растет на бедных песчаных почвах. Крапива — CR стратег, так как она доминирует на нарушенных местообитаниях.
Стратегия вида может быть пластичной. Черешчатый дуб — виолент в зоне широколиственных лесов и патиент в южной степи. Японская технология бонсай (выращивание карликовых деревьев в горшках) может быть представлена как способ превращения виолентов в патиенты.
Интересной задачей оказывается сравнение стратегий по Мак-Артуру–Уилсону и по Раменскому–Грайму. Понятно, что r-стратегам соответствуют организмы R-типа, эксплеренты. А вот K-стратегам соответствуют не только организмы C-типа, виоленты, но и те, кто относятся к S-типу, патиентам. Виоленты максимизируют свою конкурентоспособность (и емкость среды) в условиях острой конкуренции за благоприятные для потребления ресурсы, а патиенты — в условиях затрудненного потребления ресурсов. Иными словами, в задачах, которые решает дуб, конкурирующий за свет в условиях густого леса, и папоротник, выживающий при тусклом освещении в глубине пещеры, есть много общего: необходимость оптимизировать потребление ресурса, совершенствовать индивидуальную приспособленность особи.