Студенческие работы после практики II курса - 2009
Студенческие работы после практики II курса - 2011
Студенческие работы после практики II курса - 2012 (I часть)
Студенческие работы после практики II курса - 2012 (II часть)
Студенческие работы после практики II курса - 2013
Студенческие работы после практики II курса - 2014
Студенческие работы после практики II курса - 2015
Студенческие работы после практики II курса - 2016
Студенческие работы после практики II курса - 2017
Афоничева Я. В., Бондарева А. А., Баланюк Е. В., Бибик Ю. С.
Харьковский национальный университет им. В.Н. Каразина, биологический факультет, пл. Свободы, 4, г. Харьков, Украина
В ходе нормального развития головастиков зеленых лягушек (Pelophylax esculentus complex) одним и тем же стадиям развития задних конечностей могут соответствовать разные типы ротовых аппаратов. Это можно объяснить при помощи двух альтернативных гипотез: (1) определенной стадии развития задних конечностей соответствует несколько «подстадий», отличающихся по строению ротовых аппаратов; (2) каждый головастик имеет характерный для него тип ротового аппарата, с которым он и проходит все стадии развития конечности. Мы провели эксперимент для проверки этих гипотез.
Для эксперимента использовали головастиков, пойманных на левом берегу р. Северский Донец ниже по течению г. Змиева Харьковской обл. Из пойманных головастиков были составлены пары с одинаковым развитием задних конечностей (со 2-й по 7-ю стадии; см. тезисы Т. Виноградской и др. в настоящем сборнике) и одинаковыми ротовыми аппаратами (типами A и D; см. тезисы Ю. Бибик в настоящем сборнике). Мы содержали 15 пар головастиков в емкостях с 330 мл воды в течение 6 дней. Раз в сутки воду сменяли, а головастиков кормили избыточным количеством сухого аквариумного корма. Первой из проверяемых гипотез должен был бы соответствовать результат, при котором типы ротовых аппаратов головастиков изменялись бы чаще (в крайнем случае - с той же скоростью), что и стадии развития конечностей. Второму предположению соответствовало бы изменение стадий конечностей у экспериментальных при неизменности их ротовых аппаратов.
В начале эксперимента A-тип ротового аппарата был зарегистрирован только у головастиков, находящихся на относительно ранних стадиях (у всех 4- х головастиков, находящихся на 2-й стадии развития задней конечности, и у 2-х из 16 головастиков, находившихся на 4-й стадии развития конечностей). За время эксперимента эти головастики изменили тип A на D (за исключением одного погибшего и одного травмированного). Все 24 головастика с типом ротового аппарата D, находившиеся на стадиях от 3 до7, сохранили свои ротовые аппараты неизменными. Развитие задних конечностей у большинства головастиков продвинулось на одну-две стадии. Наиболее интенсивно развивались конечности, находящиеся на стадии 4 (у некоторых особей развитие продвинулось на три-четыре стадии), хотя у головастиков, сменивших ротовой аппарат, задняя конечность не изменилась.
Таким образом, мы зарегистрировали два процесса: (1) развитие задних конечностей от начальных стадий к последующим, происходящее с разной скоростью; (2) перестройку ротового аппарата от Aтипа к D(происходящую на разных стадиях развития конечностей у разных особей). Существенно, что смена типа ротового аппарата происходит в одном направлении и только на относительно ранних стадиях развития конечности. Этот результат не соответствует ни одной из тестировавшихся гипотез, хотя в целом ближе ко второй. Вероятно, характерный для каждого головастика тип ротового аппарата формируется не сразу, а проходит несколько этапов на ранних стадиях развития.
Summary. It has been investigated hind limbs and oral apparatus development character in the course of normal ontogenesis of water frogs tadpoles to examine the fits between certain stages of hind limbs development and the types of oral apparatus. According to the results of our experiment it has been suggested that the kind of oral apparatus which is typical for each tadpole is not formed at once but it passes several phases in the early stages of development.
Описанный эксперимент выполнен в ходе УИРС на учебной практике по зоологии позвоночных. Авторы благодарны доц. Шабанову Д.А. за руководство данной работой.
Афоничева Я. В., Бондарева A. A., Баланюк Е. В., Бибик Ю. С. Изучение соответствий между стадиями развития задних конечностей и ротовых аппаратов в ходе нормального развития головастиков зеленых лягушек.// "Біологія : від молекули до біосфери". Матеріали V Міжнародної конференції молодих науковців. - Харьков: Оперативна поліграфія, 2010. - с. 337-338.
РАЗЛИЧИЕ В ПРОПОРЦИЯХ И РАЗМЕРАХ ГОЛОВАСТИКОВ ЗЕЛЕНЫХ ЛЯГУШЕК, ПРОХОДЯЩИХ РАЗВИТИЕ В РАЗНОЕ ВРЕМЯ
Белкина И. О., Брилева О. П.
Харьковский национальный университет имени В. Н. Каразина, биологический факультет, пл. Свободы 4, г. Харьков, Украина
Европейские зеленые лягушки (Pelophylax esculentus complex) - интересная для изучения группа животных, в которой наблюдаются межвидовая гибридизация, полиплоидия и полуклональное наследование. Для изучения факторов, которые влияют на состав популяционных систем зеленых лягушек, необходимо изучить развитие их головастиков в естественных условиях. Мы изучили 3 выборки головастиков, собранные на левом берегу р. Северский Донец ниже по течению г. Змиева: выборка №1 (24.06.2010) состояла из 130 головастиков, выборка №2 (1.07.2010) - из 232 головастиков, выборка №3 (10.07.2010) - из 139 головастиков.
Для измерения длины тела и длины туловища был использован штангенциркуль (измерения проводились с точностью до 0,1 мм); для измерения веса - электронные весы (с точностью измерений до 0,001 г). Стадии развития головастиков были определены по задним конечностям с помощью стереоскопического микроскопа (см. тезисы Виноградской и др. в данном сборнике).
Данные обрабатывали с помощью программы Statistica for Windows 6.5
Мы зарегистрировали, что головастики, которые проходят развитие позже, имеют относительно более длинное туловище (рис. 1). Дисперсионный анализ (в котором рассматривается влияние даты сбора и стадии развития на относительную длину тела) показал, что разница между выборками высоко значима (p<0,000001)).
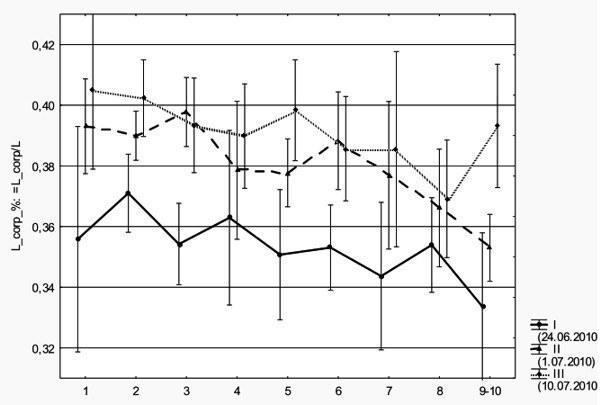
Рис. 1. Относительная длина туловища (отношение длины туловища к общей длине тела) головастиков трех выборок, собранных в разное время. По оси абсцисс отложены стадии развития задних конечностей
Summary. At comparison of tadpoles of the green frogs caught at various times (on July, 24th, on July, 1st and on July, 10th 2010) differences in their proportions are registered. At tadpoles which educe later, the attitude of length of a trunk to body total length significantly increases.
Описанные результаты получены в ходе учебно-исследовательской работы на учебной практике по зоологии позвоночных. Авторы выражают благодарность Ю. С. Бибик за помощь в выполнении работы и доц. Д. А. Шабанову за научное руководство.
Белкина И. О., Брилева О. П. Различие в пропорциях и размерах головастиков зеленых лягушек, проходящих развитие в разное время.// "Біологія : від молекули до біосфери". Матеріали V Міжнародної конференції молодих науковців. - Харьков: Оперативна поліграфія, 2010. - с. 341-342.
КАКИЕ ФАКТОРЫ ОПРЕДЕЛЯЮТ ТИП РОТОВОГО АППАРАТА ГОЛОВАСТИКОВ ЗЕЛЕНЫХ ЛЯГУШЕК?
Бибик Ю. С.
Харьковский национальный университет им. В.Н. Каразина биологический факультет пл. Свободы, 4, г. Харьков, Украина. e-mail: yulia.bibik@gmail.com
Гибридогенный комплекс европейских зеленых лягушек (Pelophylax esculentus complex) представлен двумя родительскими видами и несколькими формами полуклональных гибридов. Особенности развития головастиков разных форм зеленых лягушек до сих пор почти не описаны. Мы изучали головастиков из 7 выборок. Выборки № 1 и № 2 собраны на левом берегу р. Северский Донец ниже г. Змиева 24 июня и 1 июля 2010 г. соответственно. Выборки № 3 и № 4 выращены из кладок икры, собранных 6 мая 2010 г в пойме Северского Донца в окрестностях биостанции ХНУ, а выборка № 5 получена от пары лягушек (самка Pelophylax ridibundus, самецP. esculentus) пойманных там же. Вероятно, в состав выборок 1-5 входили как P. ridibundus,так и P. esculentus. Выборки № 6 и № 7 были собраны 18 июня 2010 г. в Донецкой области, в водоемах, населенных исключительно P. ridibundus.
Фиксированных в формалине головастиков рассматривали под стереоскопическим микроскопом, определяли тип строения ротового аппарата (рис. 1) и стадию развития задних конечностей (см. работу Виноградской и др. в этом же сборнике). Результаты определения типов ротовых аппаратов приведены в табл. 1.
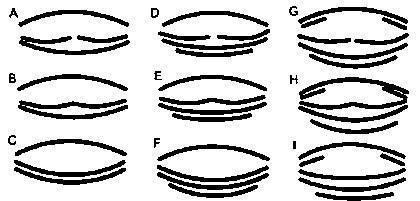
Рис. 1. Классификация типов ротовых аппаратов головастиков зеленых лягушек (условно показано взаиморасположение скоблящих роговых пластин)
Таблица 1. Разнообразие типов ротовых аппаратов в изученных выборках головастиков
|
Выборки |
К-во |
Стадии развития задних конечностей |
Типы ротовых аппаратов |
||||||||
|
A |
B |
C |
D |
E |
F |
G |
H |
I |
|||
|
1 и 2 |
352 |
1–10 |
0 |
0 |
0 |
31 |
0 |
16 |
242 |
5 |
58 |
|
3–5 |
419 |
1–8 |
81 |
18 |
30 |
216 |
12 |
14 |
48 |
0 |
0 |
|
6 и 7 |
96 |
2–8, 10 |
0 |
0 |
0 |
0 |
0 |
0 |
82 |
0 |
14 |
Выборки 3-5, развивавшиеся в одинаковых условиях, но вероятно отличавшиеся генотипу, значимо отличались по соотношениям семи зарегистрированных в них типов ротовых аппаратов (при сравнении по критерию х2 p<0,00001). Видимо, на разнообразии ротовых аппаратов отражаются генотипические особенности лягушек. В выборках 1 и 2, полученных из естественного местообитания, не найдены типы A, B, C и E, но зато найдены типы H и I. Наконец, в выборках 6 и 7, состоящих только из головастиков P. ridibundus, зарегистрировано лишь 2 типа ротовых аппаратов: G и I. Наименьшее разнообразие типов ротовых аппаратов характерно для выборок одного вида из естественного местообитания, а наибольшее — для смешанных выборок, развивавшихся в искусственных условиях. Итак, на разнообразие ротовых аппаратов головастиков влияет и их вид, и генотип, и условия их развития, но никакой из этих факторов не может исчерпывающе объяснить наблюдаемый характер изменчивости их ротовых аппаратов.
Харьковский национальный университет им. В.Н. Каразина, биологический факультет, пл. Свободы, 4, г. Харьков, Украина. e-mail: yulia.bibik@gmail.com
Задача данной работы — установить, влияет ли пребывание в составе разновозрастной группы на развитие ротовых аппаратов и конечностей головастиков представителей Pelophylax esculentus complex. Как известно (Мина, Клевезаль, 1976), групповое содержание водных животных изменяет их онтогенез по сравнению с одиночным.
Мы использовали головастиков из реки Северский Донец, собранных на левом берегу ниже г. Змиева 24.06.2010 г. Рассматривая живых головастиков под стереоскопическим микроскопом, мы выбрали две группы головастиков с одинаковым строением задних конечностей. 24 «мелких» головастика находились на 3-ей стадии развития конечностей (см. тезисы Виноградской и др.), 12 крупных — на 7 стадии. В обеих группах были представлены головастики с типами ротовых аппаратов (см. тезисы Бибик) G, D, H и I. Головастиков взвешивали и размещали в емкости (поллитровые банки), содержащие по 0,3 л речной воды. В 6 емкостей поместили по 1 «мелкому» головастику, в 6 — по 1 «крупному», и еще в 6 — по 3 «мелких» и 1 «крупному». Раз в сутки воду сменяли и кормили головастиков избыточным количеством сухого аквариумного корма. Через 10 дней провели повторные измерения массы, определили стадии развития конечностей и типы ротовых аппаратов.
У «мелких» головастиков-одиночек стадии развития конечностей изменились с 3-ей до 6-й, а в группах — с 3-ей до 4-й. У «крупных» головастиков независимо от условий содержания произошло изменение с 7-ой до 9-й стадии. Групповое содержание значимо (p=0,004) уменьшило относительный прирост массы в обеих группах, практически одинаково для «крупных» и «мелких». Таким образом, групповое содержание затормозило рост (оцененный по возрастанию массы) в обеих группах, а развитие (оцененное по состоянию конечностей) — только у «мелких».
Тип ротового аппарата у «мелких» головастиков сменялся значимо (при сравнении по Манну-Уитни p=0,005) чаще, чем у «крупных». Групповое содержание не повлияло на частоту смен типов ротовых аппаратов у «крупных» головастиков и незначимо (p=0,32) снизило ее у «мелких». У «мелких» головастиков зарегистрированы следующие сочетания начального и конечного типов ротовых аппаратов: D→D (13 случаев); D→E (1); D→G (3); G→G (2); G→H (2); H→H (1); I→G (2). Для «крупных» головастиков зарегистрированы сочетания G→G (2 случая); G→H (5); G→редукция ротового аппарата (5 случаев). Последняя группа случаев напоминает события, происходящие при метаморфозе (когда скоблящий ротовой аппарат редуцируется и развивается ротовой аппарат, предназначенный для захватывания добычи), но эти процессы происходят на более поздних стадиях развития.
Summary. Development of hind limbs and oral apparatus as well as mass growth have been studied in small and big tadpoles of water frogs(Pelophylax esculentus complex) maintained solitary or in groups (1 big and 3 small tadpoles in each group). Maintenance in groups inhibits growth both in small and big tadpoles, whereas hind limbs development is inhibited only in small tadpoles. Alteration of structure in various types of oral apparatus is also described.
Описанный эксперимент выполнен как учебно-исследовательская работа на учебной практике II курса. Авторы благодарят доцента Д. А. Шабанова за научное руководство данной работой.
Бибик Ю. С.. Коваленко М. С., Кучкова А. Г. Влияние эффекта группы на развитие головастиков зеленых лягушек. // "Біологія : від молекули до біосфери". Матеріали V Міжнародної конференції молодих науковців. Харьков: Оперативна поліграфія, 2010. -с. 338-339.
РОСТ И РАЗВИТИЕ ГОЛОВАСТИКОВ ЗЕЛЕНЫХ ЛЯГУШЕК В ЕСТЕСТВЕННОМ МЕСТООБИТАНИИ
Виноградская Т. И.,Толокнеева А. А , Комарова Е. В., Хлопонина В. В., Бибик Ю. С.
Харьковский национальный университет им. В.Н. Каразина; пл. Свободы, 4, г. Харьков, Украина
Европейские зеленые лягушки (Pelophylax esculentus complex) — гибридогенный комплекс, привлекающий внимание многих исследователей (Шабанов, Литвинчук, 2010). Для комплексного изучения этой группы животных нужно установить, как проходит их онтогенез, в том числе — личиночное развитие. После описания развития смешанных выборок головастиков, взятых из естественных ГПС, можно будет сравнить особенности личиночного развития отдельных форм лягушек.
Мы изучили головастиков, собранных на левом берегу Северского Донца ниже по течению г. Змиева в зарослях водной растительности на мелководье. Выборка № 1, собранная 24 июня 2010 года, состоит из 130 особей, а выборка № 2, собранная 1 июля 2010 года — из 232 особей.
Мерами роста головастиков служили общая длина тела с хвостом и длина туловища (которые мы измеряли штангенциркулем с точностью до 0,1 мм), а также вес (который измеряли электронными весами с точностью до 0,001 г.). Стадии развития определяли по состоянию задних конечностей. Выделяли такие стадии: 1) отсутствие конечностей; 2) зачатки задних конечностей имеют вид круглого бугорка; 3) зачатки конечностей начинают изгибаться; 4) дистальный конец конечности принимает форму лопаточки; 5) зачатки 3-х пальцев; 6) зачатки 4-х пальцев; 7) зачатки 5-ти пальцев, начинает развиваться коленный сустав; 8) сформирован коленный сустав; 9) сформированы коленный сустав и сустав между голенью и вторичной голенью, перепонки между 3-мя пальцами; 10) полностью сформирована межпальцевая перепонка. Данные обрабатывали с помощью программы Statistica for Windows 6,5.
В целом зарегистрированная связь длины (L, мм) и веса (W, г) головастиков может быть экстраполирована выражением W= 0,26 - 0,27 × L+ 0,098 × L. Значения длины и веса для отдельных стадий показаны в табл. 1.
Табл. 1. Средние значения длины и веса головастиков на разных стадиях их развития
|
Ста дия |
Кол-во особей (выборки №1+№2) |
Длина (L),мм |
Вес (W), г |
|
Стадия |
Кол-во особей (выборки №1+№2) |
Длина (L),мм |
Вес (W), г |
|
1 |
4 + 24 |
2.02 |
0,107 |
|
6 |
22 + 20 |
4,22 |
1 |
|
2 |
37 + 74 |
2,74 |
0,265 |
7 |
19 + 8 |
4,76 |
1,261 |
|
|
3 |
23 + 22 |
3,24 |
0,408 |
8 |
11 + 17 |
5,3 |
1,698 |
|
|
4 |
5 + 12 |
3,84 |
0,657 |
9 |
1 + 11 |
5,52 |
1,788 |
|
|
5 |
7 + 29 |
3,88 |
0,659 |
10 |
1 + 15 |
6,42 |
2,597 |
Как видно из табл. 1, выборки отличаются по относительной численности головастиков, находящихся на разных стадиях развития. При сравнении зарегистрированного распределения с теоретическим, построенным на основании предположения об отсутствии связи выборки и стадии развития, критерий Пирсона j2= 34,4, что при данном числе степеней свободы соответствует высоко значимым различиям (p = 0,000075). Преобладание в выборке № 2 головастиков на поздних стадиях, вероятно, связано с более длительным временем их развития.
Описанные результаты получены в ходе учебно-исследовательской работы на учебной практике по зоологии позвоночных. Авторы выражают благодарность доц. Д. А. Шабанову за руководство данной работой.
Харьковский национальный университет им. В.Н. Каразина, биологический факультет, пл. Свободы, 4, г. Харьков, Украина
Определение уровня флуктуирующей асимметрии, ФА (ненаправленных случайных отклонений от билатеральной симметрии) — один из способов оценки стабильности онтогенеза. Развитие у межвидовых гибридов обычно менее устойчиво, чем у родительских видов. В окрестностях биостанции ХНУ в с. Гайдары (Змиевской р-н Харьковской области) из числа европейских зеленых лягушек обитают представители родительского вида, Pelophylax ridibundus (озерные лягушки) и гемиклональные межвидовые гибриды, Pelophylax esculentus (съедобные лягушки). В 2008 году ФА их окраски изучали по общепринятым признакам — асимметричности дорзомедиальной полосы, количеству пятен на спине и пятен-полос на задних конечностях (Деряженцева и др., 2008). Как ни странно, ФА гибридов в этом исследовании не превышал ФА родительского вида. Мы применили для измерения ФА лягушек новый метод, предполагая, что он окажется более чувствительным к отклонениям от симметрии окраски.
В пойме р. Северский Донец в окрестностях с. Гайдары было поймано 186 зеленых лягушек. Их определяли по внешним признакам: низкий и косой пяточный бугор, отсутствие желтоватого оттенка на бедрах, темные резонаторы у самцов соответствует P. ridibundus; высокий бугор, присутствие желтого оттенка на бедрах, светлые резонаторы у самцов соответствует P. esculentus. Мы определили 107 особей (46 самок, 36 самцов и 25 полувзрослых) как P. esculentus, 55 особей (31+12+12 соответственно) — как P. ridibundus, и 24 особи (12+6+6) остались не определенными вследствие нехарактерного сочетания их признаков.
Спинную сторону туловища и конечностей каждой лягушки фотографировали (для этого лягушке давали уцепиться передними конечностями за опору и растягивали ее в длину за пальцы задних конечностей), а затем выпускали в места поимки. В программе CorelDraw на каждую фотографию была наложена сетка, разбивающая ее вдоль оси тела лягушки, от рыла до пяточных бугров, на 20 зон. Для всех зон, для которых это было возможно, определяли, симметрична в ней окраска (S), слегка асимметрична (H) или сильно асимметрична (A). Подсчитывали количество зон каждого типа: NS, NHи NA. Результаты обрабатывали в программе Statistica. Асимметричность особи вычисляли как ФА = (NA+ NH/2) / NS.
Признаки ФА, учитывающие знак отклонения от симметрии, имеют нормальное распределение со средним значением 0. Наша мера ФА принимает только положительные значения. Самыми многочисленными оказались значения, близкие к 0, с ростом показателя ФА частота встречаемости значений уменьшалась. Такое распределение подтверждает, что использованная мера характеризует ФА и может применяться для изучения устойчивости развития лягушек, и, вероятно, некоторых других животных. P. esculentus оказались значимо (при сравнении по Манну-Уитни p=0,031) асимметричнее, чем P. ridibundus. Медиана использованной меры ФА для P. esculentus составляет 0,4, для P. ridibundus — 0,33, а для неопределенных особей — 0,39. Влияние пола и возраста на ФА окраски оказалось незначимым.
Summary. For study fluctuating asymmetry (FA) tinge in water frogs, their photographed from back side. Photography was dividing along axis body on 20 zones. We counted the symmetrical, weak and hard asymmetrical zones. The tinge of hybrids form water frogsP. esculentus was found a more asymmetrical, then tinge of representatives by generation species water frogsP. ridibundus.
Работа выполнена как УИРС на учебной практике по зоологии позвоночных. Авторы благодарят доцента Д. А. Шабанова за идею работы и научное руководство ею.