Михайлова О. В., Кечеджи А. Е., Шабанов Д. А. Изучение сперматогенеза у диплоидных Pelophylax esculentus (Amphibia, Anura) при помощи кариоанализа в раздавленных препаратах // Праці Українського герпетологічного товариства. – 2011. – № 3. – С. 120-127.
УДК 597.851(576.354.4)
Изучение сперматогенеза у диплоидных Pelophylax esculentus (Amphibia, Anura) при помощи кариоанализа в раздавленных препаратах
О. В. Михайлова, А. Е. Кечеджи, Д. А. Шабанов
Харьковский национальный университет им. В. Н. Каразина,
пл. Свободы, 4, Харьков, 61077 Украина
E-mail: d.a.shabanov@gmail.com
Изучение сперматогенеза у диплоидных Pelophylax esculentus (Amphibia, Anura) при помощи кариоанализа в раздавленных препаратах. Михайлова О. В., Кечеджи А. Е., Шабанов Д. А. – Представлены результаты изучения сперматогенеза 14 половозрелых диплоидных самцов Pelophylax esculentus из Северско-Донецкого центра разнообразия зеленых лягушек (Украина, Харьковская обл.). Использован кариоанализ в препаратах раздавленных мацерированных семенников. Типичный ход сперматогенеза соответствует этапам гаметогенеза, описанным для P. esculentus из Западной Европы, причем у многих особей регистрируется значительное количество анеуплоидных клеток. У одной диплоидной особи в семенниках зарегистрированы как диплоидные клетки, из которых должны образоваться гаплоидные сперматозоиды, так и тетраплоидные клетки, которые должны дать диплоидные сперматозоиды. У этой же особи в семенниках зарегистрированы клетки с хромосомным набором, значительно превышающим тетраплоидный.
Ключевые слова: мейоз, Pelophylax esculentus, сперматогенез, гемиклональное наследование, диплоидный и тетраплоидный набор.
A Study of Spermatogenesis in Diploid Pelophylax esculentus (Amphibia, Anura) using Karyoanalysis in Squashed Preparations. Mykhailova O. V., Kechedzhi A. E., Shabanov D.A. – The results for gametogenesis of 14 mature diploid Pelophylax esculentus males from Seversko-Donetskiy center of diversity of green frogs (Ukraine, Kharkiv region) are shown. For the research caryoanalysis in squashed macerative testicles was used. Typical way of spermatogenesis is similar to the same in ovogenesis, which for P. esculentus from Western Europe were shown. Many individuals have a big amount of aneuploid cells. For the one of all examined diploid males in testicles, both diploid, giving the rise to haploid sperm, and tetraploid cells were registred. Cells with number of chromosomes largely exceeding tetraploid one were shown for the same individual.
Keywords: meiosis: Pelophylax esculentus, spermatogenesis, hemiclonal inheritance, diploid and tetraploid chromosome set.
Первое сообщение о межвидовой гибридизации у зеленых лягушек было опубликовано почти полвека назад (Berger, 1964). На протяжении всего этого времени зеленые лягушки вызывают непреходящий интерес исследователей. Уже через 10 лет после открытия Л. Бергера Х. Тюннер показал, что существование гибридных зеленых лягушек поддерживается благодаря гибридогенезу, клональному наследованию отдельных геномов (Tunner, 1974). Используя современную номенклатуру (Frost et al., 2006) и терминологию, можно сказать, что съедобная лягушка, Pelophylax esculentus (Linnaeus, 1758), является гемиклональным гибридом прудовой лягушки, Pelophylax lessonae (Camerano, 1882) и озерной лягушки, Pelophylax ridibundus (Pallas, 1771). И хотя съедобная лягушка не является видом, но в силу ряда причин для нее используется название, аналогичное видовому. Различные представители гибридогенного комплекса зеленых лягушек способны обитать совместно, образуя системы, которые предложено называть гемиклональными популяционными системами, ГПС (Шабанов, Литвинчук, 2010). В ряде ГПС кроме диплоидных гибридов встречаются полиплоидные (Plötner, 2005).
Существование ГПС зеленых лягушек поддерживается специфическим характером гаметогенеза у гибридных особей. У диплоидных гибридов в гаметы переходит один из геномов, клональный; второй геном в типичном случае элиминируется. Цитогенетические исследования механизмов этого наследования начал Х. Тюннер, открыватель необычного способа воспроизводства гибридных зеленых лягушек. В одной из публикаций, подытоживающей работу Х. Тюннера и его соавтора (Tunner, Heppich-Tunner, 1991), подчеркивается, что длительное независимое существование межвидовых гибридов возможно лишь при помощи аберрантных (по отношению к типичному половому размножению) механизмов воспроизводства. К их числу относятся партеногенез (развитие зиготы из яйцеклетки без сперматозоида), гиногенез (активация сперматозоидом яйцеклетки без слияния ядер) и гибридогенез (слияние ядер с последующей элиминацией одного из родительских геномов).
Используя флуоресцентную микроскопию и электрофорез при изучении гаметогенеза гибридных лягушек из Западной Европы (преимущественно самок), Х. Тюннер и С. Хеппих-Тюннер показали, что неклональный геном удаляется из клеток зародышевой линии ещё до начала мейоза (Tunner, Heppich-Tunner, 1991 и др.). Удивительный результат, полученный этими австрийскими исследователями, заключается в том, что гемиклональное наследование поддерживается благодаря двум различным аномалиям гаметогенеза. Удаление неклонального генома происходит во время митотических делений клеток зародышевой линии. После этого клональный геном удваивается, причем это удвоение может происходить или в ходе митотических делений оогониев, или уже в ходе мейоза, до стадии диплотены профазы I.
В 2004 г. на территории Восточной Украины были описаны ГПС зеленых лягушек, в которых существенную долю гибридов составляют триплоиды лягушек и зарегистрированы единичные тетраплоидные гибриды (Borkin et al, 2004). Регион их распространения был назван Северско-Донецким центром разнообразия зеленых лягушек (Шабанов, Литвинчук, 2010). В этом центре отсутствуют половозрелые P. lessonae, и все геномы этого вида передаются у гибридов из поколения в поколение клонально. Среди диплоидных самцов зарегистрированы такие, которые производят гаметы P. ridibundus, гаметы P. lessonae и, наконец, особи, которые производят смесь гамет P. ridibundus и P. lessonae в соотношении, характерном для каждого индивидуума.
Сказанное означает, что изучение гаметогенеза самцов гибридных лягушек из Северско-Донецкого центра разнообразия представляет значительный интерес. Данная публикация отражает результаты изучения гаметогенеза диплоидных гибридов. Для этого нами был использован метод кариоанализа по давленным препаратам В. В. Клименко (Klymenko , 2001) с некоторыми изменениями.
Материалы и методы
В работе изучены семенники 14 половозрелых диплоидных самцов Pelophylax esculentus из Северско-Донецкого центра разнообразия, пойманных в 2010-2011 гг. в окрестностях Биологической станции Харьковского национального университета им. В. Н. Каразина (с. Гайдары Змиевского р-на Харьковской обл.).
Предварительное определение зеленых лягушек проводили по комплексу внешних признаков (Шабанов и др., 2006). Окончательное определение основывалось на данных проточной ДНК-цитометрии, которая выполнялась С. Н. Литвинчуком и Ю. М. Розановым в ЦИН РАН (г. Санкт-Петербург). Для нескольких особей определение плоидности проводили на основании измерения размеров их эритроцитов (Бондарева, Шабанов, 2011); в последних случаях определение перепроверяли по описанной ниже методике в соматических тканях (тонком кишечнике).
Мы использовали методику кариоанализа в раздавленных препаратах по В. В. Клименко (Klymenko, 2001) с некоторыми модификациями.
Образцы ткани семенников помещали не менее чем на 2 сут в фиксатор из одной объемной части ледяной уксусной кислоты и трех частей 96%-ного этанола, с протравой из уксуснокислого железа (III). При необходимости образцы хранили в описанном фиксаторе.
Фрагмент ткани (около четверти семенника) переносили в 40%-ный раствор гематоксилина на ледяной уксусной кислоте для окрашивания. Образец выдерживали в красителе в течение 1,5 ч при t = 62-64 °С. Зачастую в ходе окрашивания объект распадался на части.
После окрашивания еще более мелкий образец ткани (на кончике препаровальной иглы) переносили на часовое стекло в небольшое количество ацетохлоралгидрата (40%-ный раствор хлоралгидрата в ледяной уксусной кислоте). В этом растворе происходит вымывание излишков краски и набухание клеток. В течение 5-10 мин после начала промывания ткань становится более светлой и рыхлой, что является необходимым условием для приготовления давленного препарата. На полное вымывание краски до обесцвечивания хромосом уходит около 3 ч, после этого хромосомы практически не видны.
При визуальном контроле под бинокулярным микроскопом, находящийся в капле ацетохлоралгидрата образец ткани разрезали на мельчайшие фрагменты (по несколько сотен клеток), используя микрохирургические инструменты (препаровальные иглы, заточенные на мелкозернистом точильном камне). Полученный фрагмент ткани помещали в каплю ацетохлоралгидрата на предметном стекле и накрывали 1/4 или 1/16 частью покровного стекла. На покровное стекло слегка надавливали кончиком препаровальной иглы, добиваясь раздавливания образца до слоя в одну клетку. Раздавливание образца проводили под микроскопом при увеличении в 160 раз. На приготовленном препарате выбирали клетки, содержащие хромосомы на разных стадиях митотического или мейотического деления. Выбранные пластинки микроскопировали с увеличением в 160, 640 и 1600 раз с масляной иммерсией и фотографировали с использованием цифровой USB-камеры для микроскопа.
Результаты и обсуждение
Типичный сперматогенез диплоидных P. esculentus. При изучении кариотипов из семенников диплоидных самцов мы наблюдали хромосомы на разных стадиях, соответствующих течению гаметогенеза, описанному для гибридных лягушек из Западной Европы.
Образование половых клеток начинается с митотического деления сперматогониев, диплоидных клеток зародышевой линии (рис. 1). Вероятно, как и у лягушек, которых изучал Х. Тюннер, элиминация неклонального генома происходит во время многочисленных митотических делений сперматогониев. Этот этап на нашем материале не изучен.
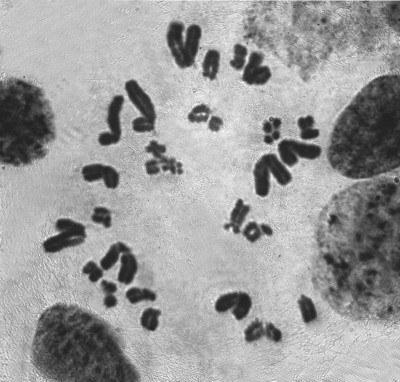
Рис. 1. Митотическое деление диплоидной клетки в семеннике P. esculentus. Стадия метафазы с 26 хромосомами
Fig. 1. Mitotic fission of the diploid cell in a P. esculentus testicle. Metaphase, showing 26 chromosomes
Сперматогонии проходят стадию роста и превращаются в сперматоциты I порядка. К метафазе I мейоза предшественники половых клеток гибридов содержат уже двойной набор хромосом (2n = 26), который несет две копии одного из родительских геномов.
Мейотическое деление начинается с профазы, которую разделяют на 5 стадий: лептотена, зиготена, пахитена, диплотена, диакинез. Профаза I мейоза характеризуется тем, что на этой стадии происходит конъюгация гомологичных хромосом и кроссинговер, обмен участками между ними (Айала, Кайгер, 1987).
Для лептотены (стадии тонких нитей) характерно начало спирализации и уплотнения хромосом; они приобретают нитевидную форму. Зиготена знаменуется конъюгацией сначала отдельных участков гомологичных хромосом, которая завершается по всей их длине к концу стадии. Соединение гомологичных хромосом друг с другом называется синапсисом. Для этой стадии характерно появление синаптонемного комплекса, входящего в состав бивалента — пары конъюгирующих хромосом.
Стадия пахитены (стадия толстых нитей) характеризуется гаплоидным числом бивалентов, т. е. фигур, образуемых конъюгирующими хромосомами. Хромосомы утолщаются и укорачиваются, и между ними происходит кроссинговер.
На стадии диплотены (стадия двойных нитей) наиболее четко видна структура бивалентов. У гомологичных хромосом остаются одна или несколько зон контакта, которые называются хиазмами. Заметна относительно большая спирализация хромосом, чем на стадии пахитены.
Диакинез характеризуется максимальным утолщением и спирализацией хромосом. Хиазмы перемещаются в направлении от центромер к концам хромосом и исчезают (рис. 2); этот процесс называется терминализацией хиазм (Айала, Кайгер, 1987).
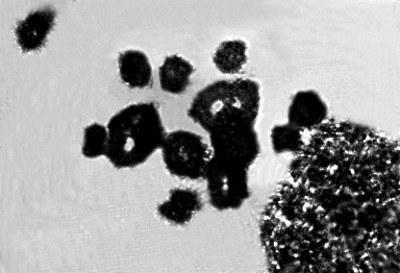
Рис. 2. Мейоз в семеннике P. esculentus. Стадия профазы I: диакинез. Видны 13 кольцевых структур: гомологичных хромосом, соединеных попарно концевыми участками на стадии терминализации хиазм
Fig. 2. Meiosis in a P. esculentus testicle. Late prophase – diakinesis, showing the terminalization of 13 ring-shaped pairs of homologous chromosomes, connected with their ends
У изученных нами гибридов зеленых лягушек хорошо заметны проявления кроссинговера: хиазмы на стадии диплотены, терминализация хиазм во время диакинеза. Важно подчеркнуть, что поскольку кроссинговер происходит между копиями одного родительского генома, он не должен приводить к генетической рекомбинации.
В течение профазы I точный подсчет количества хромосом возможен лишь на стадии диакинеза, и очень редко — пахитены.
После профазы I следует стадия метафазы I мейоза, на которой гомологи максимально сближены и образуют 13 бивалентов. Затем в анафазе биваленты распадаются, и гомологичные хромосомы расходятся к полюсам. Вокруг каждого набора гомологов образуется ядерная мембрана, и клетка делится на две дочерние. Это стадия телофазы I. В результате образуются две идентичные гаплоидные клетки — сперматоциты II порядка, которые, минуя стадию интерфазы, сразу вступают во второе деление мейоза.
Профаза II мейоза очень коротка, и клетки на этой стадии встречаются крайне редко. В метафазе II клетка содержит гаплоидный набор, который у зелёных лягушек в норме составляет 13 хромосом. Обычно две такие клетки, образовавшиеся после редукционного деления, лежат рядом (рис. 3).
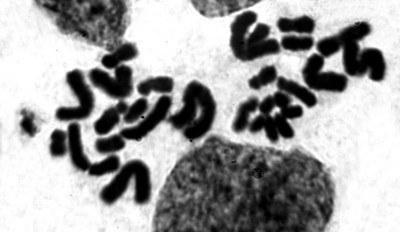
Рис. 3. Мейоз в семеннике P. esculentus. Стадия метафазы II. Видны 2 пластинки по 13
хромосом в каждой (1n)
Fig. 3. Meiosis in a P. esculentus testicle. The second metaphase of meiosis. Two plates with 13 chromosomes in each are shown
В анафазе II центромера каждой хромосомы делится, и она распадается на две хроматиды (дочерние хромосомы), которые расходятся к полюсам клетки. В результате в ранней телофазе видны «пучки» по 13 дочерних хромосом. Получившиеся гаплоидные клетки называют сперматидами. Далее они дифференцируются в спермии.
Таким образом, в результате мейотического деления диплоидной клетки у самцов образуются 4 гаплоидные гаметы. В случае гемиклонального размножения P. esculentus все гаметы одной особи несут идентичный клональный геном.
Аномалии сперматогенеза. Выше описан типичный ход сперматогенеза, от которого в ряде случаев удается наблюдать разнообразные отклонения. В семенниках гибридных зеленых лягушек регистрируется значительное количество анеуплоидных клеток, т. е. клеток, имеющих количество хромосом, некратное гаплоидному набору. Такие аномалии свидетельствуют о нестабильности процессов образования половых клеток у гибридных форм зеленых лягушек.
Особый интерес представляет ход мейоза в семенниках у одного из самцов P. esculentus, отловленного весной 2011 г. в окрестностях с. Гайдары. Эта особь была диагностирована как диплоид на основании размера ее эритроцитов. В тонком кишечнике у этой лягушки мы наблюдали исключительно диплоидные клетки. В семенниках у этого самца, наряду с типичными митотическими метафазами с диплоидным набором хромосом, найдены клетки, в которых количество хромосом превышает двойной набор. В частности нами были зарегистрированы тетраплоидные метафазные пластинки (рис. 4).
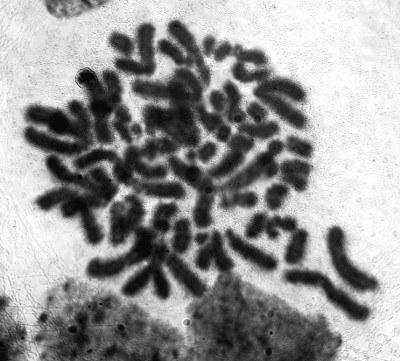
Рис. 4. Митоз в семенниках P. esculentus. Метафаза. Тетраплоидный набор хромосом (4n)
Fig. 4. Meiosis in a P. esculentus testicle. Metaphase I, showing a tetraploid chromosome set (4n)
Тетраплоидные хромосомные наборы зарегистрированы также на стадиях пахитены и метафазы I мейоза (рис. 5). Появление тетраплоидных клеток в зародышевой линии диплоидных P. esculentus можно объяснить несколькими способами. Можно предположить, что элиминация не произошла и оба родительских генома удвоились, или же после удаления неклонального генома удвоение клонального произошло дважды.
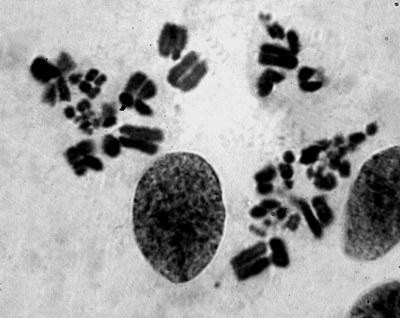
Рис. 5. Мейоз в семенниках P. esculentus. Поздняя метафаза I. Виден двойной набор бивалентов (4n). Гомологичные хромосомы хорошо различимы и обособлены, но еще соединены попарно перед самым расхождением
Fig. 5. Meiosis in a P. esculentus testicle. Late metaphase I, showing diploid number of bivalents (4n). Homologous chromosomes are detached and well observable, but still connected in pairs
Итак, у данной особи мейоз проходят диплоидные клетки, из которых должны появиться гаплоидные сперматозоиды и тетраплоидные клетки, которые должны привести к образованию диплоидных сперматозоидов. Вероятно, в случае с описанной особью можно говорить о явлении хромосомной мозаичности семенников. Это явление определяют (Ригер, Михаэлис, 1967) как наличие в ткани какого-либо органа секторов, генетически отличающихся от нормы, в связи с изменением в них количества хромосом.
В семенниках той же диплоидной особи P. esculentus, где были найдены тетраплоидные клетки, были обнаружены клетки с еще большим количеством хромосом. Эти клетки пребывали на стадии интерфазы и метафазы (рис. 6). Наличие подобных гигантских клеток рядом с тетраплоидными сперматоцитами служит, по нашему мнению, косвенным аргументом в пользу того предположения, что тетраплоидные клетки образуются вследствие избыточного удвоения клонального генома, оставшегося в клетке после элиминации неклонального генома.
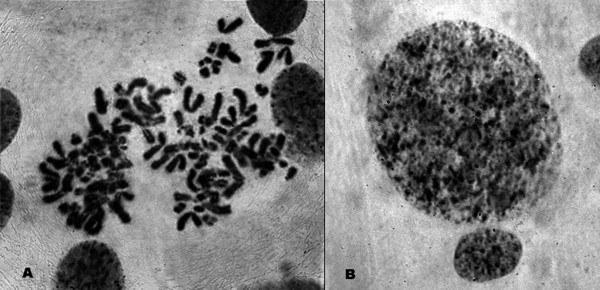
Рис. 6. Гаметогенез в семенниках P. esculentus. Полиплоидные клетки с хромосомным набором, превышающим тетраплоидный: А - Клетка на стадии метафазы; В - Интерфазные ядра: рядом с гигантским ядром располагается нормальное
Fig. 6. Gametogenesis in a P. esculentus testicle. Poliploid cells with the number of chromosomes exceeding tetraploid one are shown: A - Metaphase; B - Interphase, showing normal and giant nucleus
По мнению авторов, дальнейшее изучение гаметогенеза самцов P. esculentus из Северско-Донецкого центра разнообразия зеленых лягушек с применением кариоанализа в давленых препаратах и иных методов сохраняет научный интерес.
Благодарности
Выполнение данной работы оказалось возможным благодаря помощи коллег авторов, участвующих в исследованиях зеленых лягушек. Идея использования кариоанализа в давленных препаратах для изучения сперматогенеза гибридных лягушек принадлежит В. В. Клименко и А. И. Зиненко. Первые препараты мейоза у зеленых лягушек, полученные с применением описанной методики, были получены Г. А. Мазепой, а позже — Д. В. Дедухом. В ходе отработки методики авторы получили ценные консультации от В. В. Клименко и Е. А. Киоси. В определении плоидности зеленых лягушек неоценимую помощь оказали Ю. М. Розанов и С. Н. Литвинчук, которые исследовали предоставленные нами образцы с использованием проточной ДНК-цитометрии, а также А. А. Бондарева, которая проводила микроморфометрию эритроцитов. Авторы выражают всем названным коллегам глубокую признательность.
Айала Ф., Кайгер Дж. Современная генетика: в 3-х т. – М. : Мир, 1987. – Т. 1. – 295 с.
Бондарева А.А., Шабанов Д.А. Сравнение размера и формы эритроцитов у диплоидных и триплоидных гибридов зеленых лягушек (Pelophylax esculentus) // Всеукр. конк. студ. наук. роб. з біол. наук. – Мелітополь: «Люкс», 2011. – С. 11-14.
Ригер Р., Михаэлис А. Генетический и цитогенетический словарь. – М. : Колос, 1967. – 607 с.
Шабанов Д. А., Зиненко А. И., Коршунов А. В. и др. Изучение популяционных систем зеленых лягушек (Rana esculenta complex) в Харьковской области: история, современное состояние и перспективы // Вісн. Харків. нац. ун-ту ім. В. Н. Каразіна. Сер. : Біологія. – 2006. – Вип. 3, № 729. – С. 208–220. https://dspace.univer.kharkov.ua/handle/123456789/763.
Шабанов Д. А., Литвинчук С. Н. Зеленые лягушки: жизнь без правил или особый способ эволюции // Природа. – 2010. – № 3 (1135) . – С. 29–36. https://dspace.univer.kharkov.ua/handle/123456789/755.
Berger L. Is Rana esculenta lessonae Camerano a distinct species? // Ann. Zool. PAN. – 1964. – 22, N 13. – P. 245-261.
Borkin L. J., Korshunov A. V., Lada G. A. et al. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian Journal of Herpetology. - 2004. – 11, N 3. – P. 194–213.
Frost D. R., Grant T., Faivovich J. N, Bain R. H. et al. The Amphibian tree of life. – Bull. Amer. Mus. Nat. Hist. 2006. – 297. – 370 p.
Klymenko V.V. Parthenogenesis and cloning in the silk worm Bombyx mori: problems and prospects // J. Insect Biotechnol. аnd Sericol. – 2001. – 70. – P. 155–165.
Plötner J. Die westpaläarktichen Wasserfrösche // J. Plötner – Bielefeld : Laurenti – Verlag, 2005. – 161 s.
Tunner H.G. Die Klonale Struktur einer Wasserfroschpopulation // Z. zool. Syst. und Evolut.-forsch. – 1974. – 12, N 4. – P. 309–314.
Tunner H.G., Heppich-Tunner S. Genome exclusion and two strategies of chromosome duplication in oogenesis of a hybrid frog / // Naturwissenschaften. – 1991. – 78. – P. 32 – 34.
Коментарі
A pochemy reshili, chto eto
A pochemy reshili, chto eto kletki zarodyshevogo puti s 4n, a ne somaticheskie? Dolzhny bili by registrirovat' 26 bivalentov v diakineze, ne tak li? Na risunke 6a mogut bit' neskolko plastinok iz ongogo tsitofora.
Дело в том, что
Дело в том, что у этой конкретной особи стадия диакинеза вообще не была представлена. Ни у одной лягушки в семенниках мы не находили полного спектра стадий. У разных преобладают одна две стадии. Поэтому мы довольствовались 26-ю бивалентами в метафазе I мейоза (рис.5) и многочисленными 4n метафазами митоза. 4n клетки найденные в семенниках, не были найдены в соматических тканях этой лягушки (кишечник, кровь). А показанная на рисунке 2 пластинка с 13 бивалентами принадлежит нормальной диплоидной особи с типичным гаметогенезом (у этой лягушки было много клеток на стадии диакинеза, все по 13).