Ранняя эволюция и ископаемая история
Появление позвоночных с конечностями - стегоцефалы
Корни земноводных и амниот (пресмыкающихся, птиц и млекопитающих) следует искать среди стегоцефалов (то есть четвероногих позвоночных с пальцами), которые появились около 370 миллионов лет назад в девоне. Группа костных позвоночных, от которых возникли четвероногие (группа, которая включает в себя сохранившиеся стегоцефалов), неформально известная как костные рыбы, включает в себя две группы позвоночных лучеперых и лопастеперых. Лучеперые - группа, содержащая почти всех костных рыб, которые известны сегодня. Лопастеперые содержат три группы: современную латимерию, двоякодышащих и четвероногих. (Авторы энциклопедии придерживаются здесь кладистического подхода, при котором каждая группа включает в себя всех своих потомков, и, следовательно, млекопитающие и иные четвероногие относятся к лопастеперым рыбам; это не единственная возможная трактовка. – Прим. Д.Ш.) Наши ближайшие известные родственники, которые сохранили парные плавники – это пандерихтиды. Предки пандерихтид и четвероногих не имели спинного и анального плавников, характерных для других лопастеперых рыб, и, в отличие от них, их череп не была разделен на переднюю и заднюю части внутричерепным суставом. Кроме того, череп содержал новые покровные кости, лобные.
Пандерихтиды (Panderichthys и Elpistostege) были крупными лопастеперыми рыбами около 39 дюймов (1 м) в длину из позднего девона Европы и Северной Америки. В отличие от своих остеолепиформных родственников (например, Eusthenopteon), пандерихтиды имели массивное, уплощенное голову и тело, и весьма выступающее рыло с одним внешним носовым отверстие у края челюсти с каждой стороны черепа – этой особенностью они поделились с ранними стегоцефалами. Базальные Лучеперые (например, многопер и ильная рыба), двоякодышащие и четвероногие обладают легкими как средством дыхания, поэтому предполагается, что все ранние костные рыбы обладали легкими. В линии, ведущий к современным лучеперым, легкие были преобразованы в плавательный пузырь (орган плавучести), но у лопастеперых рыб легкие сохранились. Задачей, стоявшая перед ранними водными предками четвероногих была разработка механизма вентиляции легких в наземной среде.
Самые ранние известные стегоцефалы являются животными умеренных размеров: Ichthyostega (61 дюйм, 1,5 м. в длину) и Acanthostega (19,7 дюйма, 0,5 м. в длину) из Гренландии и Tulerpeton (около 26,4 дюймов, 0,67 м. в длину) из России. На основании останков других животных, связанных с этими стегоцефалами и особенностей их анатомии, вполне вероятно, что они вели водный образ жизни. Acanthostega и Ichthyostega, вероятно, жили в пресной или солоноватой среде, в то время как Tulerpeton жил в морской среде. На протяжении большей части двадцатого века считалось, что стегоцефалы возникли в пресной воде, но все большее число ранних амфибий находят в прибрежных, предположительно, солоноватых и морских водах. Некоторые классические пермо-карбоновые (Гарнетт, Гамильтон, и Робинсон, Канзас, Соединенные Штаты) и девонские (Мигуаша, Квебек, Канада) места ископаемых находок, которые ранее интерпретировались как пресноводные, как установлено сейчас, были прибрежными, лагунными, дельтовыми или устьевыми.
Ранние стегоцефалы имели много особенностей, связанных с водной жизни. Все они сохраняли систему боковой линии, серия сенсорных рецепторов в коже, воспринимающих механические колебания в воде и типичные для рыб, а также личинок и водных представителей современных амфибий. Acanthostega, кажется, сохранила функциональные внутренние жабры в дополнение к легким. И Acanthostega, и Ichthyostega имели хвост с плавником наподобие Panderichthys и, вероятно, так же использовали его для обеспечения движущей силы при плавании. Позвоночники этих стегоцефалов были плохо развитыми и, вероятно, неспособными поддерживать вес животного на суше. Ichthyostega имела крепкую грудную клетку. Перекрывающиеся ребра обеспечивали защиту внутренних органов, но не было достаточно подвижны, чтобы способствовать вентиляции легких в наземной среде. Конечности были короткими и толстыми, передние конечности значительно крупнее задних конечностей. Конечности, кажется, были способны только к ограниченным движениям и, как полагают, располагались скорее сбоку от тела тела, чем под ним. И передние и задние конечности были ластообразными, имея от шести до восьми пальцев у разных видов. Таким образом, представляется вероятным, что эти девонские позвоночные использовали конечности для ходьбы по погруженному субстрату, возможно, в приливной зоне или в препятствующих средах, таких как мангровые болота. Опорно-двигательный аппарат этих животных можно рассматривать как преадаптированный для наземного образа жизни. Наличие конечностей, пальцев, грудного и тазового поясов и грудной клетки, которые были полезны в водной среде обитания, обусловило морфологические признаки, которые их потомки смогли использовать для успешного освоения наземной среды обитания.
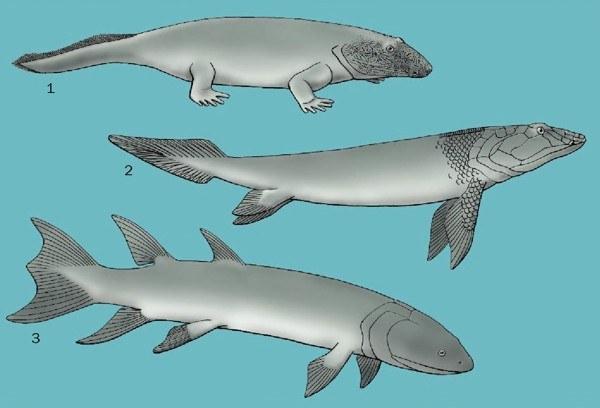
Ранние ископаемые: 1. Ichthyostega, 2. Panderichthys и 3. Eusthenopteron
Архаичные амфибии и другие ранние стегоцефалы
Есть три группы ранних стегоцефалов: стволовые тетраподы, амфибии и рептилиоморфы. Стволовые тетраподы включают всех стегоцефалов, которые появились до дивергенции между между амфибиями и рептилиоморфами, дивергенции, которая в конечном счете, привела к появлению лиссамфибий и амниот (пресмыкающихся, птиц и млекопитающих) соответственно. Стволовые тетраподы включают всех известных девонских стегоцефалов и, возможно, также много карбоновых и пермских из них. Существует значительная неопределенность, касающаяся отношений многих из этих таксонов, потому что филогения, которая долго принималась большинством палеонтологов была оспорена в конце 1990-х. Тем не менее, ясно, что основное эволюционное разделение стегоцефалов состоялось в конце девона и начале карбона, и что все основные линии (в том числе амфибии и рептилиоморфы) существовали в нижнем карбоне. Тем не менее, рептилиоморфы не были представлены в палеонтологической летописи до верхнего карбона.
Разнобразие последевонских стегоцефалов
Бафетиды (ранее известные как локсоматиды) включают в себя пять родов предположительно водных стегоцефалов из миссисипия и пенсильвания (340-305 млн лет назад) Европы и Северной Америки. Эти окаменелости известны в основном по черепам, которые обычно являются широкими и плоскими со странной формой орбит, напоминающих замочную скважину. Бафетиды могут быть родственниками стволовых тетрапод.
Три рода крокодилообразных колостеид, которые приблизительно были современниками бафетид, важны, потому что они когда-то считались тесно связаными с темноспондилами. Тем не менее, черепа этих ископаемых в Австралии и Северной Америке лишены чешуйчатой щели, особенности, которая характеризует темноспондилов, и его отсутствие колостеид и наличие органов боковой линии свидетельствуют о водном образе жизни.
Темноспондилы - это большая группа из более чем 150 описанных родов, они просуществовали от миссисипия до нижнего мела, на протяжении примерно 200 миллионов лет. Большинство ранних темноспондилов и все мезозойской представители были водными, но другие были амфибиотическими, а некоторые из них, как полагают, были наземными. Среди последних диссорофиды, которые заслуживают внимания, потому что о них утверждают,что они тесно связаны с современными амфибиями. Размеры темноспондилов изменялись от менее чем 12 дюймов (30 см) до более чем 9,8 футов (3 м) в длину. Многие, если не все, диссорофиды имели водных личинок с внешними жабрами. Некоторые личинки и/или педоморфичные (т. е. сохраняющие личиночные или ювенильные признаки в половозрелом состоянии) диссорофиды имели двувершинные (и, возможно, педицеллярные) зубы, как у лиссамфибий, следовательно, диссорофиды, как полагают некоторые ученые, тесно связаны с лиссамфибиями. Другие палеонтологи считают темноспондилов стволовыми тетраподами. Темноспондилы имели большое открытое небо (интерптеригоидные ямы), которые могут быть вовлечены в механизм щечного насоса, похожего на тот, который используют все лиссамфибии для вентиляции легких легкие. Стремечко (кость среднего уха) большинства темноспондилов более тонкое и ориентированное более латерально, чем у ранних стегоцефалов. Из-за этого, некоторые исследователи считают, что эти животные обладали тимпанумом (барабанной перепонкой) в ушной вырезке (или вырезке на чешуйчатой кости) черепа. Если эта гипотеза верна, то отсутствие тимпанума у червяг и хвостатых должно быть вторичным (т.е. следствием эволюционного потери). Позвонки ранних темноспондилов сохранили рахитомное строение, примитивное для стегоцефалов. Рахитомные позвонки имели крупный, вентральный, серповидный интерцентр и небольшие парные спинные плевроцентры, поддерживающие невральные дуги; такие позвонки плохо подходят для наземного образа жизни. У мезозойских темноспондилов позвоночник был консолидирован по стереоспондильному типу, характеризующимся большим интерцентром и небольшими плевроцентрами, которые иногда были хрящевыми. Функциональная причина этого укрепить позвоночник остается неясным, потому что большинство, если не все, мезозойские темноспондилы были исключительно водными.
Эмболомеры имели длину от 12 дюймов (30 см) до 9,8 футов (3 м) и были в основном водными и земноводными хищниками. Окаменелости эмболомеров встречаются от верхнего карбона до триаса, они известны из Европы, Северной Америки и России. Их позвонки состоят из цилиндрических интерцентра и плевроцентра. Потому что они имеют массивное стремя, считается, что эмболомеры были лишены барабанной полости, но стремя, возможно, проводило низкочастотные колебания земли и распространяющиеся в воде звуки. Эмболомеры важны, потому что раньше они думали, что они связаны с амниотами. Однако теперь многие ученые рассматривают их как еще одну группу стволовых четвероногих.
Сеймуриаморфы являются еще одной группой стволовых четвероногих, о которой раньше думали, что они тесно связаны с амниотами. Эта относительно небольшая (12 родов), но широко распространенная группа известна из перми Северной Америки, Европе и Азии, а также представлена личинками, несшими внешние жабры, и взрослыми (около 3 футов [90 см] в длину). Отсутствие органов боковой линии и жабры у взрослых показывают, что они были наземными. Взрослые сеймуриаморфы имели длинные, тонкие стремена, что позволяет предположить, что они, возможно, имели барабанную полость, и строение ребер, которые предполагают способность к реберной вентиляции легких.
Диадектоморфы представляют восемь родов верхнего карбона и нижней перми. Раньше они считались амниотами, теперь их считают тесно связанными с ними. Эти животные достигли длины 6,5 футов (2 м). Вдобавок к некоторым плотоядным или рыбоядным формам, диадектоморфы включает некоторых из самых ранних растительноядных стегоцефалов, диадектид.
Совокупность малых амфибий (большинство менее 12 дюймов [30 см] в длину) включает пять групп (аистоподы, нектридии, «микрозавры», аделогириниды и лисорофиды) эволюционного уровня, неофициально известного как «лепоспондилы». Эти животные известны из нижнего карбона до верхней перми. Некоторые, кажется, были обитали строго в водной среде (например, многие нектридии, аделогириниды), тогда как другие (например, «микрозавры» Pantylus и Tuditanus), видимо, были амфибиотичными или наземными. Большинство лепоспондильных амфибий не имело или имело только небольшую ушную вырезку или впадину чешуйчатой кости (например, аделогириниды), таким образом, они не должны были иметь барабанную перепонку. Если эти земноводные включают ближайших известных родственников лиссамфибий, то предок червяги и саламандр, вероятно, не имел барабанной перепонки, и тимпанум бесхвостых, возможно, появился только в триасе. Название группы происходит от структуры их позвонков, в которых доминируют большие, цилиндрические плевроцентры, сливающиеся с невральными дугами на ранних стадиях развития, как это происходит у лиссамфибий. У некоторых лепоспондильных амфибий сохраняется небольшой серповидный интерцентр, но у других он теряется, как и у большинства лиссамфибий.
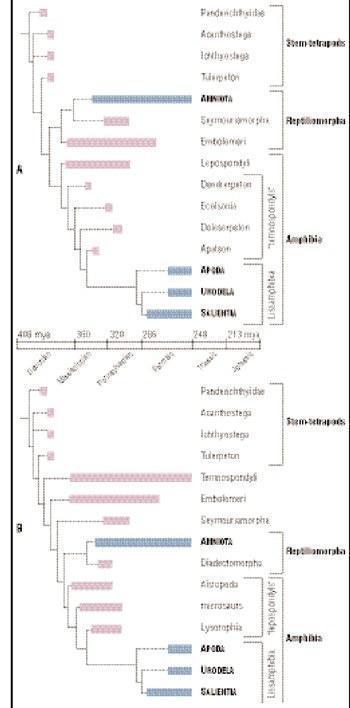
A и B: Возможные филогенетические деревья амфибий
Лиссамфибии
Самые старые из известных лиссамфибий, группы, включающей червяг, саламандр, бесхвостых и их ископаемых родственников, датируются триасом, около 250 млн лет назад. Окаменелости этой группы крайне скудны. Действительно, в триасе, который длился около 37 миллионов лет, известно только два вида лиссамфибий и оба они тесно связаны с бесхвостыми. Не все палеонтологи согласны, что лиссамфибии является естественной группой, но большинство неонтологов считают ее монофилетической. Независимо от выбираемых филогенетических отношений, существует разрыв в несколько десятков миллионов лет между сестринской группой (т. е. диссорофидами или лисорофидами [? лепоспондилами? – Прим. Д.Ш.]) и первыми известными лиссамфибиями. Начиная с юрского периода (206 млн лет назад) ископаемые лиссамфибии стали более распространенным, однако лучше всего представлены группы, которые жили водной среде.
Salientia, прыгающие (бесхвостые и Triadobatrachus)
Triadobatrachus, старейшая ископаемая лиссамфибия, происходит из нижнего триаса Мадагаскара. Как известно из одного образца, практически полного и сочлененного скелета (4.2 дюйма [10,6 см] в длину), он было тесно связан с бесхвостыми амфибиями. Он имеет много общих особенностей черепа с лягушками и жабами, но отличается от них, имея более длинное туловища, менее специализированный тазовый пояс, короткие конечности и короткий хвост. Другое, немного более молодое ископаемое, Czatkobatrachus из нижнего триаса Польши основано на нескольких изолированных костей, и, таким образом, о его сходстве или филогенетическом положении мало что можно сказать. Начиная с нижней юры, известны ископаемые, относящиеся к каждому из современных отрядов. Тем не менее, находки червяг и общее качество ископаемых хвостатых являются чрезвычайно скудными, в отличие от бесхвостых.
Ископаемый Triadobatrachus и художественная реконструкция живой особи
Triadobatrachus считается близкородственным бесхвостым (лягушкам и жабам). Вместе эти животные образуют таксономическую группу, известную как Salientia. Самые ранние известные бесхвостые, Prosalirus и Vieraella из нижней юры Аризоны (Соединенные Штаты)и Аргентины, соответственно, обитали приблизительно в то же время, что Eocaecilia, стволовая червяга. Каждое из этих некрупных по размеру (2 дюйма [50 мм] and 1.25 дюйма [30 мм] в длину от рыла до ануса, соответственно) бесхвостых отличается от более крупного триадобатрахуса наличием короткого стержня, уростиля, отсутствием хвоста,и наличием длинных задних конечностей в которых предплюсна ьыла изменена и сформировала дополнительных отдел конечности. В отличие от триадобатрахуса, эти бесхвостые, конечно, были способны к прыжкам, типичному способу локомоции бесхвостых.
Один род, Eodiscoglossus, представлен среднеюрскими остатками из Великобритании. Это весьма крупное бесхвостое (3,25 дюйма [80 мм] от рыла до ануса) замечательно напоминает современных дискоглоссид, особенно виды рода Discoglossus.

Художественная реконструкция живого представителя Prosalirus bitis
В поздней юре ископаемые остатки бесхвостых становятся более разнообразными таксономически и географически. Eobatrachus и Comobatrachus известны из Вайоминга (Соединенные Штаты), а Enneabatrachus из Вайоминга и Юры (Соединенные Штаты); все несколько похожи. Notobatrachus, из нескольких мест из Патагонии в Аргентине является одной из наиболее важных среднеюрских находок, потому что представлен полными сочлененными скелетами многих особей, включая молодых и полувзрослых. Это бесхвостое было крупным, достигая длины от рыла до ануса 5.7 дюймов (14.5 см). Оно имело несколько примитивных особенностей, включая свободные ребра у нескольких позвонков, слаборазвитый крестец, относительно короткий тазовый пояс и толстые, относительно короткие задние конечности. Это бесхвостое считается предком всех современных бесхвостых. Два других таксона, Callobatrachus и Mesophryne были описаны из юрских/меловых ископаемых отложений Китая; раньше их связывали с дискоглоссидами, но их родственные связи неизвестны. Старейшее ископаемое пиповое - это Rhadinosteus из поздней юры Юты.
В мелу произошло распростанение бесхвостых. Среди наиболее важных нижнемеловых находок дискоглоссида Eodiscoglossus из Испании и различные пиповые — Thoraciliacus, Cordicephalus и ShomroneIIa (личинки) из Израиля. Средне/верхнемеловые бесхвостые включают гобиатид из Монголии и Узбекистана, пиповых из Аргентины (Saltenia и Avitabatrachus) и Нигера (Pachybatrachus) и лептодактилид (Baurubatrachus и Estesius) из Бразилии и Боливии, соответственно.
Третичные остатки бесхвостых слишком обширны, чтобы рассказывать о них здесь. Достаточно сказать, что большинство семейств представлено в ископаемых отложениях Европы, Африки, Азии и Северной Америки

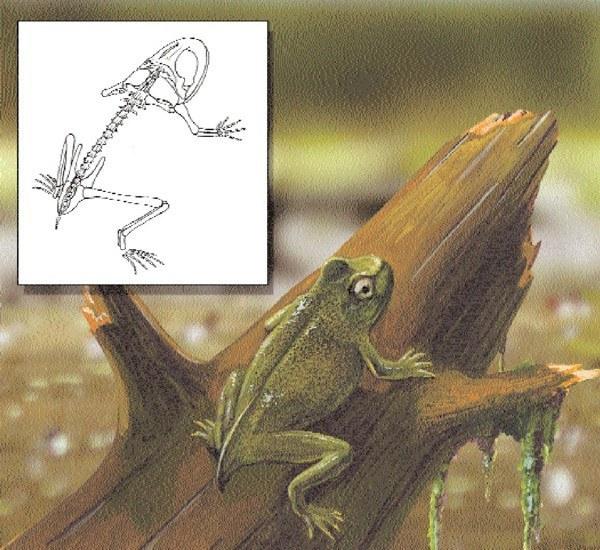
Ископаемый скелет лягушки в камне. Лягушки и жабы впервые появились 190-160 миллионов лет назад, в раннем юрском периоде
Гимнофионы (червяги и Eocaecilia)
Дошедшие до нас безногие или червяги составляют своеобразную группу лишенных конечностей, змееобразных амфибий, которые являются наземными или водными и специализированными для закапывания; они имеют щупальце с каждой стороны головы под редуцированными глазами. Большинство ученых считает червяг наиболее близкими к основанию лиссамфибий (последние находки заставляют изменить это мнение и относить червяг к лепоспондильным амфибиям – Прим. Д.Ш.). Нижнеюрское (около 204 миллионов лет назад) ископаемое Eocaecilia из Аризоны считается стволовой червягой. Она отличается от современных членов этой группы наличием маленьких, хорошо окостеневших конечностей и поясов конечностей, отсутствующих у современных червяг. Хотя глаза были крупнее, а череп содержал больше костей, чем у живущих видов, нижняя челюсть и механизм закрывания челюстей, кажется, напоминают современных представителей. Все другие ископаемые остатки, связываемые с гимнофионами, это позвонки из верхнего мела Судана, раннего палеоцена Боливии и позднего палеоцена Бразилии. Эти ископаемые дают мало полезной информации кроме того, что современные семейства червяг начали дифференцироваться в позднем мезозое.

Ископаемая Eocaecilia и художественная реконструкция живой особи
Хвостатые (саламандры и подобные ископаемые)
Ископаемые хвостатые известны из средней юры – нижнего мела (около 180—127 миллионов лет назад) из озерных и лагунных отложений Европы, верхнемеловых - эоценовых (около 90-3 5 миллионов лет назад) пойменных отложений Северной Америки и эоценовых-миоценовых (около 35-5 миллионов лет назад) бурых углей Европы. Кроме того, есть остатки из мела Боливии, Судана, Нигера и Израиля. Laccotriton, Sinerpeton, Jehohtriton и Liaoxitriton - это хвостатые из верхней юры/нижнего мела Китая. С тремя исключениями, указанными ниже, эти остатки отчетливо являются хвостатыми, то есть, относящимися к Caudata. Родственные связи Albanerpetontidae из среднеюрских – миоценовых обложений северных континентов неоднозначны. Эта группа может представлять особое, ранее ответвление линии хвостатых или отдельную группу лиссамфибий, происхождение которой предшествовало филогенетической дивергенции между хвостатыми и бесхвостыми. Филогенетический статус Ramonellus из нижнего мела Израиля неизвестен. Третий таксон, Triassurus, был описан из триаса Узбекистана. В последнем случае, если плохо сохранившиеся остатки оказываются хвостатыми, то эта группа возникла в триасе или раньше.
Самые ранние остатки, которые могут быть однозначно идентифицированы как хвостатые – это юрские находки из Китая, карауридные хвостатые Karaurus и Kokartus из Казахстана и Киргизстана и Marmorerpeton из Англии. Существующие группы саламандр представляют удивительное количество окаменелостей. (1) Sirenidae: верхний мел и палеоцен Северной Америки; меловые остатки нескольких таксонов с южных континентов могут также относиться к сиренидам; (2) Hynobiidae: верхний плиоцен Казахстана; (3) Cryptobranchidae: палеоцен монголии и России, верхний олигоцен-плиоцен Европы, верхний палеоцен-верхний миоцен Северной Америки с плиоцен-плейстоценовыми месторождениями криптобранхид в Северной Америке; (5) саламандроидные (Ambystomatidae, Amphiumidae, Dicamptodontidae, Plethodontidae, Proteidae, Rhyacontritonidae and Salamandridae): стволовые саламандроидные из нижнего мела Европы; амфиумиды из палеоцена-плейстоцена Северной Америки; дикамптодонтиды из палеоцена Канады; саламандриды из верхнего палеоцена и позже в Европе и верхнего миоцена Восточной Азии и неогена Северной Америки. Кроме того, известно три группы ископаемых хвостатых с неопределенными родственными связями: батрахозавроиды из среднего мела – нижнего плиоцена Северной Америки и Европы; скаферпетониды из верхнего мела, палеоцена и эоцена Северной Америки и, возможно, мела Азии и дикамптодонтидоподобные хвостатые из палеоцена-миоцена Европы.