Парарептилии (Parareptilia) — большая группа наземных четвероногих [4], сборная, возможно искусственная группа [3], вероятно, независимо от остальных позвоночных, достигшая рептилийного уровня организации. Также, согласно некоторым представлениям, от парарептилий ведут свое начало черепахи. [4]
Систематика
Термин Парарептилии (Parareptilia) предложен Э. Олсоном (американский палеонтолог) в 1947 году. Различные классификации дают разный состав для этой группы. [3] Для начала можно сказать, что в настоящее время сеймуриаморф и диадектов обычно относят к амфибиям, а также большинство современных авторов сближает сеймуриаморф с пареязаврами. [1]
Кэролл (американский учёный, доктор наук в области палеонтологии позвоночных, специализирующийся на палеозойских и мезозойских земноводных и пресмыкающихся) склонен рассматривать парарептилий как сестринскую группу амниот, помещая их выше вестлотианы. Ватсон (английский палеонтолог, написал много трудов по эволюции ископаемых позвоночных) разделял примитивнейших рептилий без височных окон или котилозавров (Cotilosauria), на три подотряда: Seymouriamorpha, Diadectomorfa и Captorhinomorpha. В диадектоморф он включил диадектов и проколофонов, обладавших ушной вырезкой, и перейазавров, вторично утративших ушную вырезку. Диадекты помещались в основании диадектоморфных котилозавров, которые позднее составляли ядро парарептилий при их выделении. [1]
По Э. Олсону парарептилии первоначально объединяли сеймуриаморф, диадектов, проколофонов, пайеязавров и черепах. Сеймуриаморфы, поставленные в ранг подкласса, были соединены с диадектами, две остальные группы диадектоморфных котилозавров – проколофоны и пареязавры – выделялись отдельно. Олсон считал черепах более близкими к диадектам, а не к парейазаврам. Парарептилии противопоставлялись эврептилиям, включавших всех прочих рептилий. Сеймуриаморфы, по Олсону, считались предками диадектов. [1]
Позднее Олсон выделял сеймуриаморф в особый класс Batrachosauria и рассматривал в качестве парарептилий только проколонофоров и пайеязавров. Позднее он вообще отказался от концепции парарептилий, из которых последовательно, вслед за черепахами, были исключены сеймуриаморфы и диадекты. [1]
Ф. Хюне (немецкий палеонтолог, занимался изучением ископаемых пресмыкающихся) объединил всех парарептилий с антракозаврами в виде особой ветви эволюции тетрапод – Reptiliomorpha. В эту ветвь он включает и черепах, которых он, выводит от диадектоморф. В целом он выводил всех рептилий от рептилиоморф, но парарептилий он выделял в отдельную от рептилий ветвь эволюции. [1]
Ромер (американский палеонтолог и морфолог, автор учебников по палеонтологии, анатомии позвоночных) рассматривает сеймуриаморф вместе с диадектами в составе амфибий, а проколофонов и пайеязавров объединяет в подотряде Procolophonia отряда котилозавров. Туда же он относит и миллерозавров. [1]
М. Хитон воссоединил сеймуриаморф с диадектами в общем отряде Cotylosauria и перенёс его в класс амфибий. Он акцентировал внимание на том обстоятельстве, что отряд первоначально был выведен на основе рода Diadectes=Empedocles. Проколофонов, парейазавров и капториноморф. Хитон оставил в рептилиях, но исключил их из состава котилозавров. Котилозавров, выделяемых в подкласс Batrachosauria он разделяет на сеймуриаморф и диадектоморф. Хитон исключает из состава сеймуриаморф семейство Discosauriscidae, представители которого вели амфибийный образ жизни. Лантанозух в числе батрахозавров не рассматривается. В число батрахозавров Хитон включает Nycteroleteridae. [1]
Готье (палеонтолог Йельского университета), Клюге и Роу (профессор палеонтологии из Техасского университета) выделяют парарептилий, батрахозавры и диадекты перенесены обратно в амфибий. Авторы исключают из основного состава черепах. Первым ответвлением от общего ствола они считают зверообразных рептилий. [1] Solenodonosaurus помещается между сеймуриаморфами и диадектоморфами. [1]
М. Ф. Ивахненко (русский палеонтолог, доктор биологических наук) вернулся к названию и первоначальному объему группы, и переработал систематику парарептилий на основе изучения разнообразных их представителей из верхней перми и триаса. Им описаны новые богатые фауны позднепермских и раннетриасовых проколонофоров, показана принадлежность к лантанозухам Chalkosaurus. Ивахненко включил в состав парарептилий сеймуриаморф. Также парарептилии был объединены с другими рептилиями в классе Reptiliomorfa. Ивахненко устанавливает новый отряд Nycteroleteromorfa, в котором объединены никтеролеты и лантанозухи. Никтеролеты стоят в основании диадектозавров перед диадектоморфами. В 2001 году Ивахненко разбил парарептилий на 3 подотряда: Seymouriamorpha, Procolonomorpha и Cheloniamorpha. [1]
Так, согласно М. Ф. Ивахненко (2001), парарептилии — в классе рептилиоморф (Reptiliomorpha). В данной схеме в состав парарептилий входят сеймуриаморфы, проколофоны (надотряд Procolophonomorpha) и хелониаморфы (надотряд Cheloniamorpha). В последний надотряд входят никтеролетеры, токозавры, лантанозухи, рипейазавры, парейазавры и черепахи. В целом в этой классификации парарептилии выглядят естественной группой, возникшей от амфибийных (анамний) предков-сеймуриаморф, независимо достигшей амниотного уровня и приведшей в итоге к появлению современных черепах. [1]
Так как мы используем систематику составленную М. Ф. Ивахненко, систематика подкласса Parareptilia выглядит следующим образом:
Надотряд Procolophonomorpha
Отряд Procolohponida
Подотряд Nyctiphruretia
Семейство Nyctiphruretidae
Подотряд Procolophpnia
Семейство Spondylolestidae
Семейство Procolophonidae
Надотряд Cheloniamorpha
Отряд Pareiasaurida
Подотряд Nycteroleterina
Надсемейство Nycteroleteroidaea
Семейство Nycteroleteridae
Подсемейство Nycteroleterinae
Подсемейство Bashkyroleterinae
Семейство Tokosauridae
Семейство Rhipaeosauridae
Надсемейство Lanthanosuchoidea
Семейство Lanthaniscidae
Семейство Lanthanosuchidae
Надсемейство Elginioidea
Семейство Elginiidae
Подотряд Pareiasaurina [2]
Семейство Bradysauridae
Семейство Pareiasauridae
Время существования
Если учитывать только вымершие формы парарептилй то время их существования можно отнести к таким периодам как триас (252,17 - 201,3 млн лет назад) и пермь (298,9 - 252,2 млн лет назад) [1, 4]. Но если учитывать тот факт что современные черепахи относятся к этой группе, то их существование продолжается и по сей день.
Регионы наиболее распространенных находок
Наибольшее количество находок найдено в Европейской и Восточной части России(Оренбургская область, Башкирия, Архангельская обл., Кировская обл., Татария), Северной Америке (Колорадо, Катлер Юра, Оклахома), Западной Европе (Германия, Шотландия, Англия), Южной и Восточной Африке (Нигерия, Морадис), также имеются некоторые находки в Китае (провинция Шаньси), Бразилии (один род и один вид), Мадагаскаре и Антарктиде.
Морфологические особенности
Многие представители группы мозаично сочетают в своем строении признаки, свойственные как амфибиям, так и ящерам. Одной из архаичных особенностей многих парарептилий является отсутствие в строении черепа височных окон (полостей позади глазниц, в которых у многих рептилий располагаются мощные челюстные мышцы). [4]
Короткий параболический череп с крышей ангуститабулярного типа (табличатые кости соприкасаются с теменными) с уплощенным закрытым нёбом. Радиальная структура крыши черепа ямчато-гребенчатая. Вдоль всех швов крыши черепа имеются гладкие полосы, окружённые пористым валиком. Кости щечного комплекса соединяются с теменным щитом на всем протяжении контакта зубчатым швом без резкого перегиба между теменным щитом и щекой. На затылочном крае щеки по всей высоте располагается глубокая (ушная) вырезка. Хорошо выражены парафенестральные и базиптеригоидные крылья парасфеноида (длинная кость, лежащая в основании черепа). У наиболее примитивных форм не сформирован комплекс атлант-эпистофей. Вероятнее всего имелся покров из роговых чешуй. [1]
Образ жизни
В основной своей массе парарептилии - сравнительно небольшие (до 0,5 м в длину) хищники, питавшиеся беспозвоночными (никтеролетеры, некоторые проколофоны); в конце поздней перми среди них появляются крупные растительноядные формы - парейазавры. [4]
Яркие представители
Nyctiphruretus acudens найдены скелеты и черепа. Другие находки: р. Мезень в Архангельской области (Россия), Лешуконский район, Глядная Щьеля, Козьмогородское, Усть-Няфта, Пеза-I, Мезенский район. Челюстные зубы с высокими, чуть расширенными коронками, овальными в основании (рис. 1, 2). [2]

Рис. 1. Реконструкция
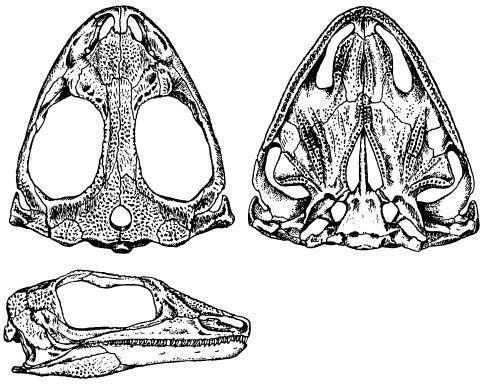
Рис. 2. Череп
Limnoscelis padulis – в черепе имелось небольшое задневисочное окно между верхнее-наружным концом парокципетального отростка и tabulare или ниже и внутри от затылочного фланга squamosum (рис. 3, 4). [1]
Рис. 3. Реконструкция
Tseajaia campi имел длину черепа 12,5 см. Череп умеренно высокий. Имеется крупное supraoccipetale; задневисочных окон нет. Челюстное сочленение расположено немного ниже затылочного. Задний край крыши черепа образован парными таблитчатыми и заднетеменными костями. Теменное отверстие очень большое, продольно удлиненное. Имеется длинная ушная вырезка. Хорошо развита межптеригоидная ямка неба. Поперечные фланги птеригоидов хорошо развиты, зубов не имеют. Небные зубы не развиты. Челюстные зубы конические, заостренные, примаксиллярно удлиненные. Посередине максилярного зубного ряда 1 зуб имеет подобие клыка. Подвздошная кость с длинным задним отростком (Рис. 3; 4). [1]

Рис. 3. Компюторная реконструкция
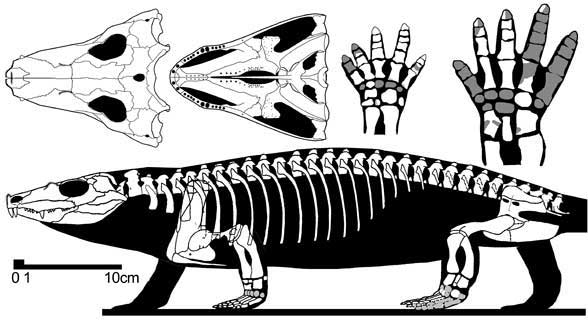
Рис. 4. Скелет
Tokosaurus perforatus - Очень маленькая форма, длина черепа не более 3 см. Отдельные округлые остеродермы (вторичные кожные окостенения, располагающиеся в мезодермальном слое кожи) только в щечной области черепа [2]. Имел зарождающееся височное окно (Рис. 5; 6) [1]. Челюстные зубы тонкие, остроконические, передние зубы верхнечелюстной кости не увеличены [2].
Рис. 5. Реконструкция
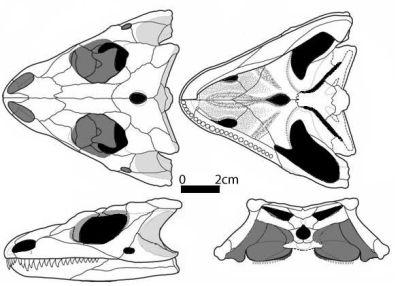
Рис. 6. Череп
Lanthaniscus efremovi – найден в Архангельской области [1] и описан по скелету [2] (Рис. 7; 8).

Рис. 7. Реконструкция
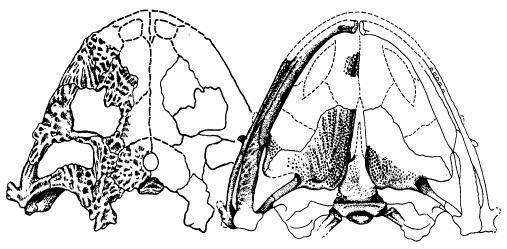
Рис. 8. Череп
Lanthaniscus watsoni обладает широким и резко уплощенным черепом, длиной около 19 см (рис.). Крыша черепа покрыта мелкоячеистой скульптурой, в щечной области усложненной продольными гребнями. Щечная область выступает назад от уровня затылочного и челюстного сочленений, задний край щеки орнаментирован выростами, расположенными главным образом на верхневисочных и квадратноскуловых костях; местами выросты имеются и на нижних краях черепа – в области челюстного сочленения, под глазницами и несколько позади уровня ноздрей. Немного позади глазниц имеется большое височное окно, ограниченное, как нижнее височное окно диапсид, заглазничной, скуловой, квадратноскуловой и чешуйчатой костями. Глазница по нижнему краю переходит в обширную вырезку, в которой могла размещаться железа, которая могла иметь функцию солевой, выделяющей избытки катионов. Заглазничные кости очень большие и широко соединяются с надвисочными костями. Ушная вырезка не развита, слуховая косточка не обнаружена (Рис. 9; 10). [1]
Рис. 9. Реконструкция
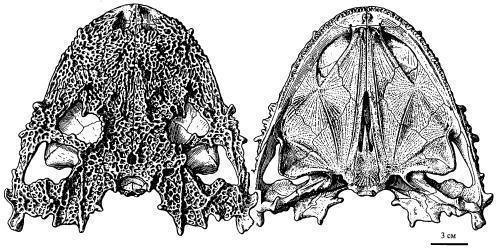
Рис. 10. Череп
Nanoparia luckhoffi имел длину черепа 15 см (Рис. 11; 12). [1]

Рис. 11. Реконструкция
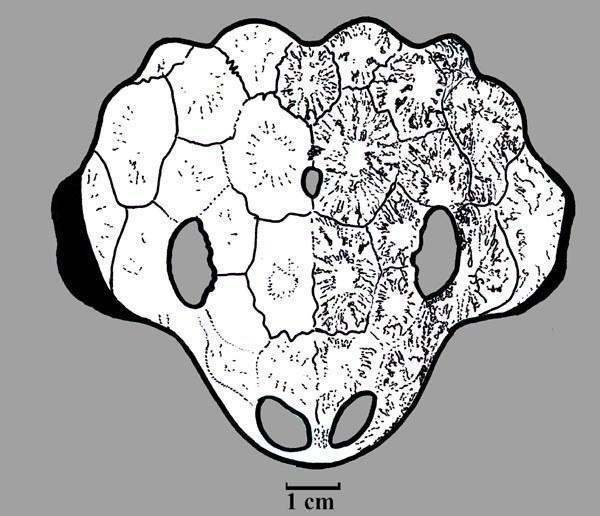
Рис. 12. Череп
Scutosaurus karpinskii имел очень тонкую подошву stapes. Сесамовидные кости прилегали к заднему краю supratemporale или squamosum. Ивахненко указывает, что ювенильные зубы имели плоскую коронку с округлым верхним краем, несущую до 12 зубцов. Зубы постепенно приобретали корень, врастающий в кость. Спинной панцирь отсутствовал (Рис. 13; 14). [1]
Рис. 13. Реконструкция
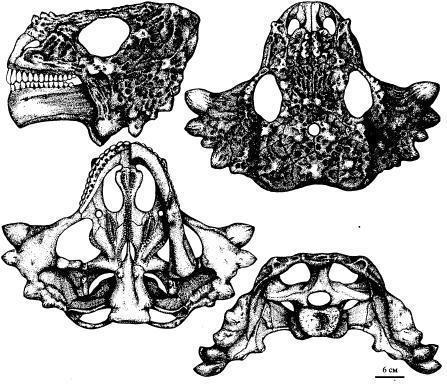
Рис. 14. Череп
Elginia mirabilis в длину имеет около 90 см. Более маленькие срединные выросты черепа элгинии расположены не на теменных костях, а на находящихся позади них дополнительных элементах, соответствующих, скорее всего, передним панцирным пластинкам, приросшим к черепу. Небо характеризуется относительно слаборазвитыми птеригоидными поперечными отростками, направленными, косо вперед и наружу. Зубов на птеригоидных флангах у элгинии нет. Хоаны позади характерно изогнуты внутрь. Зубной ряд укороченный, до 12 девятизубцовых зубов в верхней челюсти. Спинные ребра не расширенные, подвздошная кость с хорошо развитым передним отростком (Рис. 15; 16). [1]
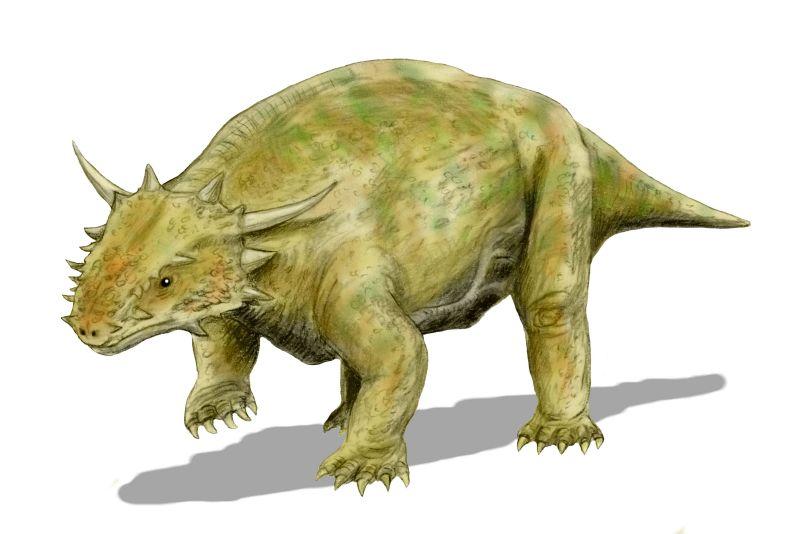
Рис. 15. Реконструкция
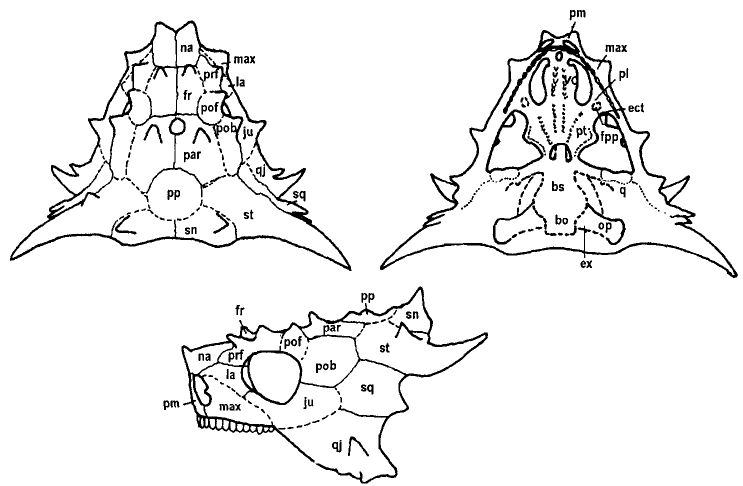
Рис. 16. Череп.
Owenetta kitchingorum заднетеменные образуют узенький и короткий задний выступ (Рис. 17; 18). [1]

Рис. 17. Реконструкция
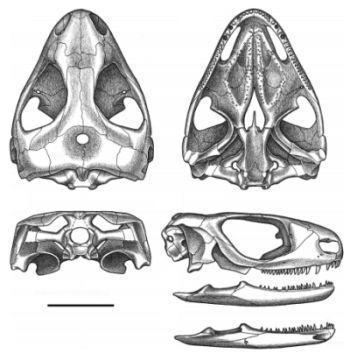
Рис. 18. Череп и нижная челюсть.
Список используемой литературы:
1. Л.П. Татаринов "Очерки по эволюции рептилий" - М. ГЕОС. - 2006 г. - 234 стр.
2. М.Ф. Ивахненко, Е.Н. Курочкин "Ископаемые позвоночные России и сопредельных стран" - М., ГЕОС - 2008 г. - 374 стр.
3. https://ru.wikipedia.org/wiki/Парарептилии
4. https://files.school-collection.edu.ru/dlrstore/4a71b57f-702d-448c-b206-...