|
|
|||
|
БІОСИСТЕМИ. БІОСФЕРА
ЕКОСИСТЕМИ. ПОПУЛЯЦІЇ
ОРГАНІЗМИ У ДОВКІЛЛІ
ЛЮДСТВО ТА ЙОГО ДОЛЯ
|
|||
|
IV-15. Екологічні стратегії |
|||

IV-15. Екологічні стратегії
Як визначити цінність особи для популяції?
«Природний добір визнає лише один різновид «валюти» — успішне потомство» (Е. Піанка, 1981).
Ми говорили, що популяція — потенційно безсмертна сутність, що складається із смертних особин. Щоб підтримати існування популяції, особина має вижити сама й залишити нащадків, які теж зможуть вижити. Зверніть увагу на двоїстість цього завдання. Ймовірно, найбільші шанси на виживання матиме та особина, яка взагалі не витрачатиме ресурси та отриману з них енергію на виробництво потомства. Але мине трохи часу — і така особина безслідно зникне з популяції. На протилежному «полюсі» знаходиться гіпотетична особина, яка відразу після появи починає всю свою енергію спрямовувати на виробництво нащадків. Така істота загине сама і, якщо її нащадки успадкують такий самий неефективний спосіб розподілу ресурсів, призведе на світ нащадків, які не матимуть шансів на виживання.
Відтак, найбільшу цінність для популяції повинна мати особина, яка поєднує витрати на власне виживання та виробництво нащадків в оптимальному поєднанні. Можна оцінити, наскільки це поєднання оптимальне. Для цього треба вирахувати, за якого поєднання в цих умовах особина залишить найбільший можливий внесок у майбутнє покоління. Захід, який використовується для цього в математичній популяційній біології, називається репродуктивною цінністю. Репродуктивна цінність — узагальнена міра виживання та плодючості, що враховує відносний внесок організму в майбутні покоління.
«Легко описати гіпотетичний організм, який має всі ознаки, необхідні для досягнення високої репродуктивної цінності. Він розмножується майже відразу після народження, дає численне, крупне, захищене потомство, про яке піклується; розмножується багаторазово та часто протягом довгого життя; він перемагає у конкурентній боротьбі, уникає хижаків і легко здобуває їжу. Описати таку істоту легко, але уявити важко......» (М. Бігон та ін., 1989).
Ви розумієте, що така неможливість випливає із суперечливості завдань самопідтримання та розмноження (рис. 4.15.1). Одним із перших це усвідомив у 1870 р. англійський філософ Герберт Спенсер, який говорив про альтернативність підтримки організмом власного існування та продовження себе в нащадках. Сучасною мовою можна сказати, що ці параметри пов'язані негативними кореляціями, відношенням, у якому поліпшення системи за одним параметром має супроводжуватися її погіршенням за іншим.

Рис. 4.15.1. У коловратки Asplanchna шанси на виживання зменшуються зі зростанням плодючості (Піанка, 1981).
Різні види (і різні популяції) по-різному перерозподіляють енергію між самопідтримкою та розмноженням. Можна говорити про видову стратегію, що виражається в тому, як представники виду видобувають ресурси і як вони їх витрачають. Успішною може бути лише та стратегія, за якої особини отримують достатню кількість енергії, щоб вони могли зростати, розмножуватися та компенсувати всі втрати на активність хижаків та різноманітні нещастя.
Ознаки, що стосуються різних адаптивних стратегій, можуть бути пов'язані відношенням трейдоффа, тобто непереборними негативними кореляціями (відношенням або-або). Так, відношенням трейдоффа пов'язані кількість нащадків та їх виживання, швидкість зростання та стійкість до стресу тощо. Американські екологи Р. Мак-Артур та Е. Вілсон описали у 1967 році два типи видових стратегій, які є результатом двох різних типів добору та пов'язані відношенням трейдоффа. Ухвалені позначення цих стратегій (r- і K-) взято з логістичного рівняння.
Згідно з логістичною моделлю, у зростанні популяції можна виділити дві фази: з прискореним і з уповільненим зростанням (рис. 4.15.2). Поки N невелика, на приріст популяції здебільшого впливає сомножитель rN, і зростання популяції прискорюється. На цій фазі (r-фазі) зростання популяції прискорюється, і її чисельність тим вища, чим вища здатність особин до розмноження. Коли значення N стає досить високим, на чисельність популяції починає впливати в основному сомножитель (K-N)/K. На цій фазі (К-фазі) зростання популяції сповільнюється. Коли N=K, (K-N)/K=0, і зростання чисельності популяції припиняється. На K-фазі чисельність популяції тим вища, чим більший параметр K. Він тим вищий, чим вища конкурентоспроможність особини.
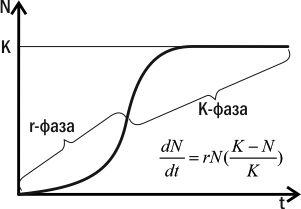
Рис. 4.15.2. r- та K-фази популяційного зростання відповідно до логістичної моделі
Можна припустити, що популяції деяких видів переважну частину часу перебувають у r-фазі. У таких видів максимальну репродуктивну цінність мають особини, здатні швидко розмножуватися і захоплювати незаселене середовище своїми нащадками. Іншими словами, на цій фазі добір сприятиме підвищенню параметра r — репродуктивного потенціалу. Такий добір називається r-добором, а види, що виникають в його результаті, — г-стратегами.
У видів, популяції яких переважну частину часу перебувають на K-фазі, ситуація зовсім інша. Максимальна репродуктивна цінність у цих популяціях буде властива особинам, які виявляться настільки конкурентоспроможними, що зможуть отримати свою частку ресурсу навіть за умов його нестачі; тільки в цьому випадку вони зможуть розмножитися і зробити свій внесок у наступне покоління. Популяція, яка складається з таких особин, матиме більш високе значення параметра K — ємності середовища, ніж та, що складається з особин, які не «вміють» боротися за ресурси. На цій стадії на популяцію діє К-добір, результатом якого є поява видів К-стратегів. K-добір спрямований на збільшення витрат на розвиток кожної особи та підвищення її конкурентоспроможності.
Між цими стратегіями можливі переходи, але вони мають проміжний характер, а не поєднують типові прояви двох форм.
«Не можна бути одночасно салатом і кактусом» (Е. Піанка).
Важливе значення для визначення, який саме добір (r- чи K-) діятиме на вид, має динаміка зміни кількості доступного ресурсу і гострота конкуренції за нього. При різкому невиборчому скороченні чисельності популяцій, викликаному браком ресурсу, зумовленому зовнішніми причинами, переваги набувають r-стратеги, а за конкурентної боротьби за недостатній ресурс — K-стратеги.
Вибір між r-стратегією (підвищенням плодючості) і K-стратегією (підвищенням конкурентоспроможності) — досить простий, проте він зачіпає безліч параметрів організмів та їх життєвих циклів. Порівняємо ці стратегії в їхній типовій формі (табл. 4.15.1).
Таблиця 4.15.1. Особливості r-і K-добору та стратегій
|
Характеристики |
r-добір та r-стратеги |
K-добір та K-стратеги |
|
Клімат |
Мінливий, не передбачуваний |
Постійний, передбачуваний |
|
Смертність |
Катастрофічна, незалежна від щільності популяції |
Викликана конкуренцією, яка залежить від щільності популяції |
|
Крива смертності |
Зазвичай типу III |
Зазвичай типу I или II |
|
Розмір популяцій |
Мінливий, нерівноважний |
Постійний, близький до граничної ємності середовища |
|
Вільні ресурси |
Поява вільних ресурсів, заповнення «екологічного вакууму» |
Вільних ресурсів майже не буває, вони зайняті конкурентами |
|
Внутрішньо- та міжвидова конкуренція |
Слабка |
Гостра |
|
Розмір тіла |
Відносно дрібний |
Відносно великий |
|
Розвиток |
Швидкий |
Медленное |
|
Статева зрілість |
Рання |
Пізня |
|
Швидкість розмноження |
Висока |
Низька |
|
Розмноження впродовж життя |
Часто одноразове |
Неодноразове |
|
Потомків у виводку |
Багато |
Мало, часто один |
|
Кількість ресурсу на одного потомка |
Низька |
Висока |
|
Тривалість життя |
Коротка |
Довга |
|
Пристосування |
Примітивні |
Довершені |
|
Оптимізується |
Продуктивність |
Ефективність |
Може здатися дивним, чому для r-стратегів характерне одноразове розмноження, а для K-стратегів неодноразове. Цю особливість легше пояснити прикладом. Уявіть собі мишей, що заселяють комору з зерном (ресурсу вдосталь, конкуренції ніякої). Розглянемо стратегії двох видів.
Вид № 1. Статевозрілість у 3 місяці, кількість нащадків у виводку — 10, самка живе рік і здатна розмножуватися кожні три місяці.
Вид № 2. Статевозрілість у 3 місяці, кількість нащадків у виводку — 15, вигодувавши їх, самка гине від виснаження.
У першому випадку через три місяці до розмноження приступить 10 нащадків та їхні батьки (всього 12 голів), а в другому — цілих 15 нащадків. Більш високу швидкість захоплення вільних ресурсів зможе забезпечити другий вид. Типова r-стратегія змушує особин викладатися в розмноженні якомога раніше і якнайсильніше, і тому r-стратеги часто обмежені одним-єдиним сезоном розмноження.
З іншого боку, легко зрозуміти чому типові K-стратеги розмножуються багато разів. У конкурентному середовищі виживе тільки той нащадок, на розвиток якого витрачено багато ресурсу. З іншого боку, щоб вижити та розмножуватися, доросла особина повинна витрачати значну кількість енергії на власну підтримку та розвиток. Тому в граничному випадку K-стратеги приносять по одному нащадку за один раз (як, наприклад, слони та кити, а також, у більшості випадків, і люди). Але хоч якими б досконалими були ці тварини, пара батьків згодом загине. Щоб популяція не припинилася, пара батьків повинна залишити пару нащадків, що вижили, а отже, народити, повинна більше двох. Відтак, необхідною умовою виживання K-стратегів виявляється багаторазовість розмноження складових їх особин.
1935 р. радянський ботанік Л.Г. Раменський виділив три групи рослин, які він назвав цінотипами (поняття про стратегії ще не було сформовано): віоленти, патієнти та експлеренти. У 1979 році їх ці групи (під іншими назвами) відкрив заново англійський еколог Дж. Грайм (рис. 4.15.3). Ці стратегії такі.
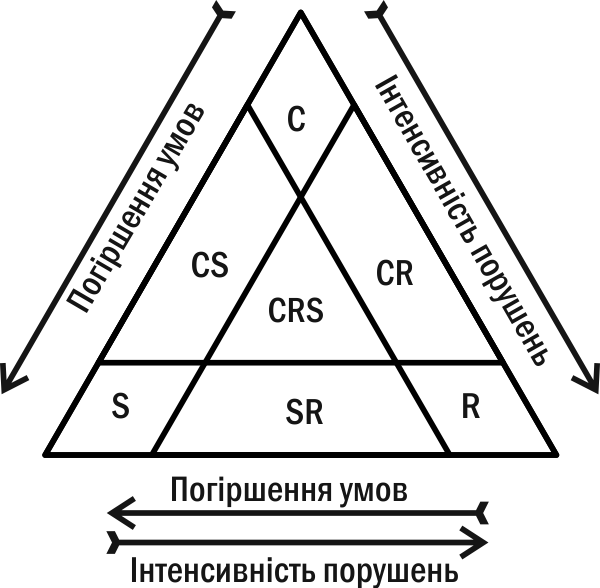
Рис. 4.15.3.«Трикутник Грайма» — класифікація видових стратегій
– Тип C (competitor, конкурент), віолент по Раменському; витрачає більшу частину енергії на підтримання життя дорослих організмів, домінує у стійких спільнотах. Серед рослин до цього типу найчастіше відносяться дерева, чагарники чи потужні трави (наприклад, дуб, очерет).
– Тип S (stress-tolerant, стрес-толерант); патієнт по Раменському; завдяки спеціальним адаптаціям виносить несприятливі умови; використовує ресурси там, де з ним за них майже ніхто не конкурує. Зазвичай це повільно зростаючі організми (наприклад, сфагнум, лишайники).
– Тип R (від лат. ruderis, рудерал), експлерент по Раменському; заміщає віолентів у зруйнованих спільнотах чи використовує тимчасово незатребувані іншими видами ресурси. Серед рослин це однорічники або дворічні, які виробляють безліч насіння. Таке насіння утворює банк насіння в грунті або здатне ефективно поширюватися на значну відстань (наприклад, кульбаба, іван-чай). Це дозволяє таким рослинам чекати на момент вивільнення ресурсів або вчасно захоплювати вільні ділянки.
Багато видів здатні поєднувати різні типи стратегій. Сосна належить до категорії CS, оскільки вона добре росте на бідних піщаних ґрунтах. Кропива — CR стратег, бо вона домінує на порушених місцеперебуваннях.
Стратегія виду може бути пластичною. Черешчастий дуб — віолент у зоні широколистяних лісів та патиент у південному степу. Японська технологія бонсай (вирощування карликових дерев у горщиках) може бути представлена як спосіб перетворення віолентів на патієнти.
Цікавим завданням виявляється порівняння стратегій за Мак-Артуром-Вілсоном і за Раменським-Граймом. Зрозуміло, що r-стратегам відповідають організми R-типу, експлеренти. А ось K-стратегам відповідають не лише організми C-типу, віоленти, а й ті, хто належать до S-типу, патієнтів. Віоленти максимізують свою конкурентоспроможність (і ємність середовища) в умовах гострої конкуренції за сприятливі для споживання ресурси, а патієнти — в умовах утрудненого споживання ресурсів. Іншими словами, у завданнях, які вирішує дуб, що конкурує за світло в умовах густого лісу, і папороть, що виживає при тьмяному освітленні в глибині печери, є багато спільного: необхідність оптимізувати споживання ресурсу, вдосконалювати індивідуальну пристосованість особини.