|
|
|||
|
БІОСИСТЕМИ. БІОСФЕРА
ЕКОСИСТЕМИ. ПОПУЛЯЦІЇ
ОРГАНІЗМИ У ДОВКІЛЛІ
ЛЮДСТВО ТА ЙОГО ДОЛЯ
|
|||
|
← V-25. (доповнення) Обмін речовиною, енергією та інформацією |
V-26. (доповнення) Фактори, що впливають на розвиток організму |
V-27. (доповнення) Тиск на глибині: витримування та подолання → |
|

V-26. (доповнення) Фактори, що впливають на розвиток організму
Спадковість, оточення, випадковість — ось три речі, які керують нашою долею.
Акутагава Рюноске
Становище людини у суспільстві визначається трійцею — обдарованістю, працею, випадком.
Володимир Шкода
Тісний взаємозв'язок організму та середовища проявляється й у впливі середовища на розвиток. Нині стали вельми поширеними уявлення про те, що спадкові задатки організму (пов'язані передусім з ДНК) задають «норму реакції» — діапазон можливих результатів розвитку. Роль середовища в цих уявленнях полягає в тому, щоб визначити, який варіант цієї норми реакції буде обраний. Мінливість, пов'язану зі зміною норми реакції, прийнято вважати спадковою, генотиповою (мутаційною) мінливістю та ототожнювати з невизначеною мінливістю за Ч. Дарвіном. Різноманітність усередині норми реакції прийнято розглядати як прояв неуспадкованої, фенотипної (модифікаційної) мінливості. Її прийнято вважати тим, що Дарвін назвав певною мінливістю. Викладений опис цілком узгоджується з точкою зору синтетичної теорії еволюції (СТЕ). На думку авторів цієї роботи, ця теорія погано узгоджується з сучасними даними і значно поступається одній зі своїх альтернатив — епігенетичної теорії еволюції (ЕТЕ).
Насамперед, слід зазначити, що спадкова мінливість зовсім не є невизначеною, залежною від екологічної потреби у ній. Класичні досліди, які показали, що зміни в генах не залежать від потреб у них, були виконані Дж. Ледебергом на кишкових паличках, які в результаті мутації втратили здатність розщеплювати лактозу (молочний цукор). Таких бактерій вирощували на середовищі, де не було лактози, а потім пересівали туди, де єдиним джерелом їжі була ця речовина. Більшість таких бактерій гинули, але деякі (у яких сталася зворотна мутація, що дала їм можливість харчуватися лактозою) виживали. Експериментатори показали, що мутація, яка дозволяла розщеплювати потрібну речовину, відбувалася ще на вихідному, безлактозному середовищі, тобто була випадковою та ненаправленою.
Здавалося б, усе зрозуміло. Але у 1988 р. Nature надрукував статтю Дж. Кейрнса (J. Cairns), який лише злегка змінив умови класичного експерименту. У Кейрнса нездатні харчуватися на новому середовищі бактерії не гинули. Вони залишалися живими, проте через недостатнє харчування не могли ділитися (М.Д. Голубовський, 2001). У цих умовах бактерії інтенсивно перебудовували свій спадковий апарат, і деякі з них незабаром набували необхідних ознак. Мутанту, поставленому перед вибором загинути чи повернутися до норми, вдається «підібрати» зворотну мутацію та стати нормальним!
Друге зі спрощень, наведених вище, полягає в тому, що всі набуті ознаки носять неспадковий характер. Свого часу пошук спадкових змін, викликаних впливом середовища, вівся дуже прямолінійно. Наприклад, Август Вейсман доводив неспадковість набутих ознак, просто відрізаючи мишам хвости. Справді, протягом значної кількості поколінь довжина хвостів у нащадків таких мишей залишалася постійною. І. І. Мечников зазначив з цього приводу, що мучити мишей не було сенсу: покоління за поколінням жінки, перед тим як залишити нащадків, позбавляються цноти, але, покоління за поколінням, гімен (дівоча плева) знов утворюється у їхніх дочок. Якщо результати травм не успадковуються, це не означає, що жодні ознаки не успадковуються взагалі.
Т. Д. Лисенко на своєму дослідному господарстві годував корів шоколадом. Корови давали дуже жирне молоко (ще б пак!), а Лисенко сподівався, що ця властивість передасться їхнім нащадкам. Не передалась. Але у 1984 журнал Science опублікував статтю Дж. Л. Маркса, в якій той згадав про «привид Лисенка». Коли рослини льону вирощували на ґрунті з надлишком мінерального харчування, ті виростали високими та потужними. Однак коли їх нащадків вирощували на звичайному ґрунті, ті все одно у багатьох поколіннях зберігали частину батьківської високорослості. Як було показано в ряді статей (огляд — див. Грант, 1991), при надлишку харчування в хромосоми льону вбудовувалися (і стійко передавалися нащадкам) додаткові копії гена, що прискорює зростання.
У 2005 році виявлено (стаття Р. Пруітта в Nature), що у резуховидки Таля (Arabidopsis thaliana), класичного об'єкта молекулярної генетики, при схрещуванні двох особин, гомозиготних за дефектною версією гена hothead 10 % нащадків мають нормальний стан. Очевидно, у разі йдеться про корекції ДНК по молекулам РНК, які були синтезовані на нормальних генах батьків рослин-мутантів і зберігалися в їхніх генах.
2006 року той самий журнал опублікував результати роботи М. Рассользадегана та інших співробітників інституту INSERM у Франції. Вони працювали з генетично зміненими бурими хом'ячками (Scotinomys), використовуючи лінію зі штучно спричиненою мутацією гена Kit. Хом'ячки з двома мутантними версіями гена Kit гинуть, носії мутантної та нормальної версій несуть характерні білі мітки на шерсті, а носії двох нормальних генів мають (повинні мати!) нормальний зовнішній вигляд. Однак з'ясувалося, що зовнішні ознаки мутації зберігалися і в тих генетично нормальних хом'ячків, батьком, матір'ю або більш віддаленим предком яких були хом'яки з генами мутантів. Цей феномен називається парамутацією і полягає в тому, що генетично нормальні, але зовні змінені особини передають прояви аномалії у потомстві протягом кількох поколінь (хоч і з поступовим слабшанням). Очевидно, що синтезована на мутантних генах РНК розбивається на фрагменти, але зберігається в клітині, зв'язуючись з якимись переносниками. Особливо багато таких фрагментів виявляється у статевих клітинах. У клітинах генетично нормальних нащадків ці РНК втручаються у роботу нормальних генів та відтворюють аналогічні собі молекули. Ймовірно, у їх присутності синтезована за нормальним геном РНК виявляється аномальною.
Австралійські імунологи Е. Стіл, Р. Ліндлі та Р. Бланден у книзі з характерною назвою «Що, якщо Ламарк правий? Імуногенетика та еволюція» (2002) виступили з добре обґрунтованою концепцією, згідно з якою імунні клітини передають у гамети інформацію про антигени, з якими стикалися, забезпечуючи успадкування набутих ознак. Хоча у прямій формі припущення цих авторів не підтвердилися, характерне повернення до ідей, які впродовж століття вважали «віджилими».
З іншого боку, дійсні мутації проявляються та успадковуються дуже нестійко. Їхній прояв у особин з однаковою спадковістю дуже нестійкий і коливається від їхнього повного прояву до відсутності. Винятком із цього правила є мутації, пов'язані з блокуванням синтезу якогось продукту генної активності, який є у нормальних клітин. Наприклад, блокування синтезу пігменту призведе до альбінізму. З іншого боку, поява генної послідовності, яка може забезпечити синтез нового пігменту, не обов'язково призведе до зміни забарвлення. Прихильники синтетичної теорії еволюції (СТЕ) ілюструють дію мутацій на прикладах, коли у того чи іншого виду існують два стійкі можливі шляхи розвитку і є добре налагоджений генетичний перемикач між ними (рослини гороху з жовтим/зеленим і гладким/зморшкуватим насінням у дослідах Грегора Мена. Мутації, що виникли вперше, не мають таких властивостей.
Нестійкість втілення мутацій можна спостерігати навіть на різних сторонах тіла у двобічно симетричних організмів, або на різних сегментах тіла метамерних істот. Права та ліва сторони, різні сегменти відрізняються один від одного за ступенем прояву мутації. З погляду епігенетичної теорії еволюції причина цього полягає в тому, що мутації не проходили добору на стійкість втілення в онтогенезі.
Навпаки, якщо навіть нестабільні ознаки проходять стабілізуючий добір, їх втілення в онтогенезі стає дедалі стійкішим. Це було переконливо показано у дослідах К. Х. Уоддінгтона. У цих дослідах добір дрозофіла на здатність формувати аберації dumpy або bithorax у відповідь на температурні або токсичні впливи призводив до того, що вони починали розвиватися і за нормальних умов (Waddington, 1957). В експериментах Г. Х. Шапошникова (1978) в результаті добору аберацій попелиць, викликаних зміною кормової рослини, виникали нові, морфологічно та екологічно своєрідні форми експериментальних тварин, репродуктивно ізольовані від вихідних (фактично було отримано новий вид організмів).
Як було вперше показано Р. Гольдшмідтом, зовнішнім впливом на організм, що розвивається, можна викликати ефект, еквівалентний мутації. Зовнішнім впливом в розвитку мутантного організму можна забезпечити формування нормального фенотипа. Це означає, що онтогенез може призвести до фіксованого числа кінцевих станів, вибір яких залежить як від спадковості, так і впливів середовища. Для їхньої демонстрації зручна модель, запропонована К. Х. Уоддінгтоном. Розвиток організму порівнюється в цій моделі зі скочуванням кульки по поверхні складної форми — епігенетичного (epi — над, після) ландшафту. Емергентна властивість системи управління розвитком — наявність каналізованих (від каналу — спрямованого шляху) варіантів розвитку — креодів. На епігенетичному ландшафті креодам відповідають поглиблення (канавки; рис. 5.26.1).

Рис. 5.26.1. Епігенетичний ландшафт за Уоддінгтоном. А. — Типовий генотип із добре зарегульованим розвитком; найстійкіший креод веде до нормального фенотипу. Б. — Зміна генотипу викликала перебудову епігенетичного ландшафту і уможливило розвиток по одному з аберантних креодів. В. — Зовнішній вплив може бути причиною аберантного розвитку навіть за нормального генотипу. Г. — Зовнішній вплив може бути причиною нормального розвитку навіть за зміненого генотипу
Епігенетичний ландшафт моделює роботу епігенетичної системи — системи керування розвитком. Епігенетична система може бути описана як розподіл імовірності різних підсумків розвитку при певному значенні факторів, що діють на цей розвиток (спадкових, пов'язаних з ДНК; спадкових, не пов'язаних з ДНК; середовищних).
Ті чи інші креоди практично ніколи не є результатом роботи одного гена. Система з дуже великою кількістю ступенів свободи самоорганізується, значно зменшуючи їх кількість на виході. Ще у 1920-х роках С. С. Четверіков з'ясував, що особини з нормальним фенотипом генетично дуже різноманітні. У переважної більшості ознак немає «гена норми», нормальність задана всім генотипом (рис. 5.26.2). Коли становлення нормального фенотипу неможливе, з'являються аберації (відхилення). Особини з однаковими відхиленнями розвитку виявляються генетично різноманітними (як і нормальні особини). Те, що особини з різним генотипом розвиваються нормально, є наслідком тривалого стабілізуючого добору на нормальність розвитку. Справа в тому, що норма — це фенотип, який відповідає умовам, що найчастіше зустрічаються. Попри міфологію СТЕ, життєвий успіх чи невдача особини залежить не від наявності чи відсутності в неї «хорошого» чи «поганого» алелю, а від її фенотипу в цілому. Якщо з покоління в покоління переважно виживають носії нормального фенотипу, це призводить до того, що все більш різноманітні генотипи починають розвиватися нормальним шляхом.
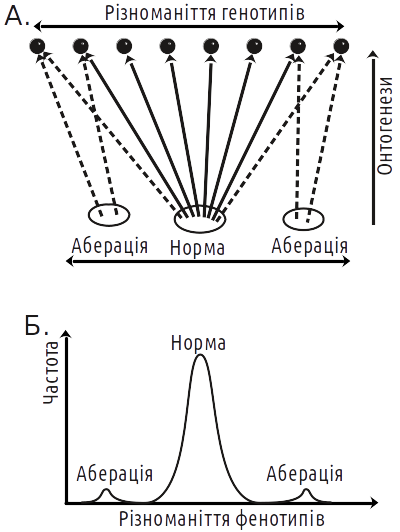
Рис. 5.26.2. Якби онтогенез не був процесом, що самоорганізується, у постійних умовах різноманітність фенотипів повторювала б різноманітність генотипів (з урахуванням випадкового впливу зовнішнього середовища). Емергентна взаємодія різних факторів у процесі розвитку призводить до того, що розподіл фенотипів сильно відрізняється від нормального. Каналізованість розвитку призводить до того, що фенотиповій нормі відповідають різні генотипи. Водночас, результат розвитку кожної окремої зиготи не можна передбачити зі 100% ймовірністю
Як видно з наведених прикладів, «спадкові» та «неспадкові» ознаки — це лише крайні точки єдиної шкали з вибудованими ознаками, розвиток яких у результаті стабілізуючого добору зарегульовано краще (і здійснюється в широкому діапазоні умов) чи гірше (і відбувається лише у певних випадках). Нарешті, як свідчить творець сучасної версії епігенетичної теорії еволюції М. О. Шишкін, поняття генотипної та фенотипної мінливості взагалі відносяться до різних ситуацій. Про генотипну мінливість заведено говорити, порівнюючи розвиток різних генотипів в однакових умовах, а про фенотипну — при порівнянні розвитку однакових генотипів у різних умовах. Як показують дані експериментів, у всіх випадках результат розвитку організму може бути передбачений лише імовірнісним чином, через опис набору можливих результатів розвитку та ймовірності кожного з них.
Головною формою добору з цього погляду є стабілізуючий добір (рис. 5.26.3). Поки умови середовища відповідають нормі, особи, які пройшли у своєму розвитку за найбільш ймовірним креодом, матимуть максимальні шанси на виживання та залишення потомства. При цьому не має значення, який внесок у нормальний розвиток заданий «спадковими» та «неспадковими» ознаками: він визначений усім епігенетичним ландшафтом. «Рухомий» добір, як його представляють прихильники СТЕ, є вигадкою, яка ніколи не реєструвалася в експериментах. Добір проти норми призводить до дестабілізації норми та розширення діапазону мінливості. В умовах, що змінюються, цей ефект якраз дозволяє «намацати» ті варіанти розвитку, які зможуть стати новою нормою. При цьому не має значення, на якій основі розвиватимуться ці нові варіанти. Навіть якщо їх розвиток відповідає класичній схемі «модифікацій» (цілком залежить від специфічного впливу середовища), добір на здатність розвивати такий фенотип призведе до стабілізації онтогенетичного шляху, що веде до нього.
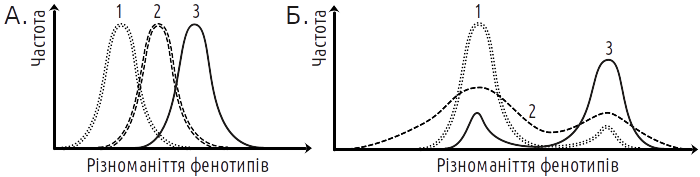
Рис. 5.26.3. Результати добору (на більше значення показаної по осі абсцис ознаки) відповідно до різних уявлень. А. — Результат, що очікується з погляду синтетичної теорії еволюції (СТЕ). Добір зумовлює зміщення норми реакції. Б. — Результат, очікуваний з погляду епігенетичної теорії еволюції (ЕТЕ). Добір призводить до дестабілізації розвитку і згодом — до його стабілізації на новій нормі. Реєстровані в експериментах результати більшою мірою відповідають другому варіанту
Додаткові матеріали: