Українська мова (найновіша версія) / Русский язык (обновление прекращено)
|
5.27. (дополнение) Давление на глубине: претерпевание и преодоление |
5.26. (дополнение) Факторы, влияющие на развитие организма
Наследственность, окружение, случайность — вот три вещи, управляющие нашей судьбой.
Акутагава Рюноскэ
Положение человека в обществе определяет троица — дар, труд, случай.
Владимир Шкода
Тесная взаимосвязь организма и среды проявляется и во влиянии среды на развитие. В настоящее время широкое распространение сохраняют представления о том, что наследственые задатки организма (связанные прежде всего с ДНК) задают «норму реакции» — диапазон возможных результатов развития. Роль среды, в этих представлениях, состоит в том, чтобы определить, какой вариант из этой нормы реакции будет избран. Изменчивость, связанную с изменением нормы реакции, принято считать наследственной, генотипической (мутационной) изменчивостью и отождествлять с неопределенной изменчивостью по Ч. Дарвину. Разнообразие внутри нормы реакции принято рассматривать как проявление ненаследственной, фенотипической (модификационной) изменчивости. Ее принято считать тем, что Дарвин назвал определенной изменчивостью. Изложенное описание вполне согласуется с точкой зрения синтетической теории эволюции (СТЭ). По мнению авторов данной работы, эта теория плохо согласуется с современными данными и значительно уступает одной из своих альтернатив — эпигенетической теории эволюции (ЭТЭ).
Прежде всего, следует отметить, что наследственная изменчивость вовсе не является неопределенной, не зависящей от экологической потребности в ней. Классические опыты, показавшие, что изменения в генах не зависят от потребностей в них, были выполнены Дж. Ледебергом на кишечных палочках, которые в результате мутации потеряли способность расщеплять лактозу (молочный сахар). Таких бактерий выращивали на среде, где не было лактозы, а потом пересевали туда, где единственным источником пищи было это вещество. Большинство таких бактерий гибло, но некоторые (те, у кого произошла обратная мутация, давшая им возможность питаться лактозой) выживали. Экспериментаторы показали, что мутация, позволявшая расщеплять нужное вещество, происходила еще на исходной, безлактозной среде, то есть была случайной и ненаправленной.
Казалось бы, все ясно. Но в 1988 году Nature напечатал статью Дж. Кэйрнса (J. Cairns), который лишь слегка изменил условия классического эксперимента. У Кэйрнса неспособные питаться на новой среде бактерии не погибали. Они оставались живы, однако из-за недостаточного питания не могли делиться (М.Д. Голубовский, 2001). В этих условиях бактерии интенсивно перестраивали свой наследственный аппарат, и многие из них вскорости приобретали нужный признак. Поставленному перед выбором погибнуть или вернуться к норме мутанту удается «подобрать» обратную мутацию и стать нормальным!
Второе из упрощений, приведенных выше, заключается в том, что все приобретенные признаки носят ненаследственный характер. В свое время поиск наследственных изменений, вызванных воздействием среды велся весьма прямолинейно. Например, Август Вейсман доказывал ненаследственность приобретенных признаков, попросту отрезая мышам хвосты. Действительно, на протяжении значительного числе поколений длина образующихся хвостов у потомков таких мышей оставалась постоянной. И. И. Мечников отметил по этому поводу, что мучить мышей не было смысла: поколение за поколением женщины, перед тем как оставить потомков, лишаются девственной плевы, а та поколение за поколением образуется у их дочерей. Если результаты травм не наследуются, это не означает, что не наследуются никакие признаки вообще.
Т.Д. Лысенко на своем опытном хозяйстве в Горках Ленинских кормил коров шоколадом. Коровы давали очень жирное молоко (еще бы!), а Лысенко надеялся, что это свойство передастся их потомкам. Не передалось. Но в 1984 году журнал Science опубликовал статью Дж. Л. Маркса, в которой тот вспомнил о «призраке Лысенко». Когда растения льна выращивали на почве с избытком минерального питания, те вырастали высокими и мощными. Однако когда их потомков выращивали на обычной почве, те все равно во многих поколениях сохраняли часть родительской высокорослости. Как было показано в ряде статей (обзор — см. Грант, 1991), при избытке питания в хромосомы льна встраивались (и устойчиво передавались потомкам) дополнительные копии гена, ускоряющего рост.
В 2005 году обнаружено (статья Р. Пруитта в Nature), что у резуховидки Таля (Arabidopsis thaliana), классического объекта молекулярной генетики, при скрещивании двух особей, гомозиготных по дефектной версии гена hothead 10% потомков имеют номальное строение и восстанавливают дикое состояние гена. По всей видимости, в данном случае речь идет о коррекции ДНК по молекулам РНК, которые были синтезированы на нормальных генах родителей растений-мутантов и сохранялись в их генах.
В 2006 году тот же журнал опубликовал результаты работы М. Рассользадегана и других сотрудников института INSERM во Франции. Они работали с генетически измененными бурыми хомячками (Scotinomys), используя линию с искусственно вызванной мутацией гена Kit. Хомячки с двумя мутантными версиями гена Kit гибнут, носители мутантной и нормальной версий несут характерные белые отметины на шерсти, а носители двух нормальных генов имеют (должны иметь!) нормальный внешний вид. Однако выяснилось, что внешние признаки мутации сохранялись и у тех генетически нормальных хомячков, отцом, матерью или более отдаленным предком которых были хомячки с мутантными генами. Этот феномен называется парамутацией и заключается в том, что генетически нормальные, но внешне измененные особи передают проявления аномалии в потомстве в течение нескольких поколений (хотя и с постепенным ослабеванием). Видимо, синтезированная на мутантных генах РНК разбивается на фрагменты, но сохраняется в клетке, связываясь с какими-то переносчиками. Особенно много таких фрагментов оказывается в половых клетках. В клетках генетически нормальных потомков эти РНК вмешиваются в работу нормальных генов и воспроизводят аналогичные себе молекулы. Вероятно, в их присутствии синтезируемая по нормальному гену РНК оказывается аномальной.
Австралийские иммунологи Э. Стил, Р. Линдли и Р. Бланден в книге с характерным названием «Что, если Ламарк прав? Иммуногенетика и эволюция» (2002) выступили с хорошо обоснованной концепцией, согласно которой иммунные клетки передают в гаметы информацию о тех антигенах, с которыми сталкивались, обеспечивая наследование приобретенных признаков. Хотя в прямой форме предположения этих авторов не подтвердились, возвращение к идеям, которые на протяжении столетия считали «отжившими», весьма характерно.
С другой стороны, действительные мутации проявляются и наследуются очень неустойчиво. Их проявление у особей с одинаковой наследственностью весьма неустойчиво и колеблется от их полного проявления до отсутствия. Исключением из этого правила являются мутации, связанные с блокированием синтеза какого-то продукта генной активности, который присутствует у нормальных клеток. Например, блокирование синтеза пигмента приведет к альбинизму. С другой стороны, появление генной последовательности, которая может обеспечить синтез нового пигмента, вовсе не обязательно приведет к изменению окраски. Сторонники синтетической теории эволюции (СТЭ) иллюстрируют действие мутаций на примерах, когда у того или иного вида существуют два устойчивых возможных пути развития и наличествует хорошо отлаженный генетический переключатель между ними (растения гороха с желтыми/зелеными и гладкими/морщинистыми семенами в опытах Грегора Менделя). Впервые возникшие мутации не обладают такими свойствами.
Неустойчивость воплощения мутаций можно наблюдать даже на разных сторонах тела у двустороннесимметричных организмов, или на разных сегментах тела метамерных существ. Правая и левая стороны, разные сегменты оказываются отличающимися друг от друга по степени проявления мутации. С точки зрения эпигенетической теории эволюции причина этого состоит в том, что мутации не проходили отбора на устойчивость воплощения в онтогенезе.
Напротив, в случае, если даже нестабильные признаки проходят стабилизирующий отбор, их воплощение в онтогенезе становится все более устойчивым. Это было убедительно показано в опытах К. Х. Уоддингтона. В этих опытах отбор у дрозофил на способность формировать аберрации dumpy или bithorax в ответ на температурные или токсические воздействия приводил к тому, что они начинали развиваться и при нормальных условиях (Waddington, 1957). В экспериментах Г. Х. Шапошникова (1978) в результате отбора аберраций тлей, вызванных сменой кормового растения, возникали новые, морфологически и экологически своеобразные формы экспериментальных животных, репродуктивно изолированные от исходных (фактически был получен новый вид организмов).
Как было впервые показано Р. Гольдшмидтом, внешним воздействием на развивающийся организм можно вызвать эффект, эквивалентный мутации. Внешним воздействием на развитие мутантного организма можно обеспечить формирование нормального фенотипа. Это означает, что онтогенез может привести к фиксированному числу конечных состояний, выбор которых зависит как от наследственности, так и от влияний среды. Для их демонстрации удобна модель, предложенная К. Х. Уоддингтоном. Развитие организма сравнивается в этой модели со скатыванием шарика по поверхности сложной формы — эпигенетическому (epi — над, после) ландшафту. Эмергентное свойство системы управления развитием — наличие канализованных (от канала — направленного пути) вариантов развития — креодов. На эпигенетическом ландшафте креодам соответствуют углубления (канавки; рис. 5.26.1).
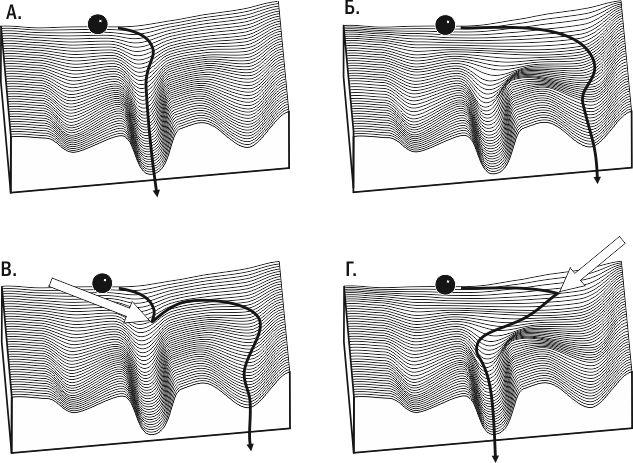
Рис. 5.26.1. Эпигенетический ландшафт по Уоддингтону. А. Типичный генотип с хорошо зарегулированным развитием; самый устойчивый креод ведет к нормальному фенотипу. Б. Изменение генотипа вызвало перестройку эпигенетического ландшафта и сделало вероятным развитие по одному из аберрантных креодов. В. Внешнее воздействие может быть причиной аберрантного развития даже при нормальном генотипе. Г. Внешнее воздействие может быть причиной нормального развития даже при измененном генотипе
Эпигенетический ландшафт моделирует работу эпигенетической системы — системы управления развитием. Эпигенетическая система может быть описана как распределение вероятности различных итогов развития при определенном значении действующих на это развитие факторов (наследственных, связанных с ДНК; наследственных, не связанных с ДНК; средовых).
Те или иные креоды практически никогда не являются результатом работы одного гена. Система с очень большим числом степеней свободы самоорганизуется, значительно уменьшая их количество на выходе. Еще в 1920-х годах С. С. Четвериков выяснил, что особи с нормальным фенотипом генетически весьма разнообразны. По подавляющему большинству признаков не существует «гена нормы», нормальность задана всем генотипом (рис. 5.26.2). Когда становление нормального фенотипа оказывается невозможным, появляются аберрации (отклонения). Особи с одинаковыми отклонениями развития оказываются генетически разнообразными (как и нормальные особи). То, что особи с разным генотипом развиваются нормально, является следствием длительного стабилизирующего отбора на нормальность развития. Дело в том, что норма — это фенотип, соответствующий наиболее часто встречающимся условиям. Вопреки мифологии СТЭ, жизненный успех или неудача особи зависит не от наличия или отсутствия у нее «хорошего» или «плохого» аллеля, а от ее фенотипа, как целого. Если из поколения в поколение преимущественно выживают носители нормального фенотипа, это приводит к тому, что все более разнообразные генотипы начинают развиваться по нормальному пути.
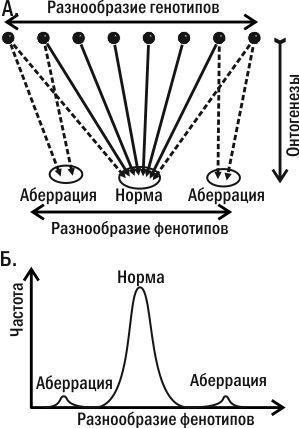
Рис. 5.26.2. Если бы онтогенез не был самоорганизующимся процессом, в постоянных условиях разнообразие фенотипов повторяло бы разнообразие генотипов (с учетом случайного влияния внешней среды). Эмергентное взаимодействие разных факторов в процессе развития приводит к тому, что распределение фенотипов оказывается сильно отличающимся от нормального. Канализованность развития приводит к тому, что фенотипической норме соответствуют весьма различные генотипы. В то же время результат развития каждой отдельной зиготы не может быть предсказан со 100% вероятностью
Как видно из приведенных примеров, «наследственные» и «ненаследственные» признаки — это лишь крайние точки единой шкалы, на который выстроены признаки, развитие которых в результате стабилизирующего отбора зарегулировано лучше (и осуществляется в широком диапазоне условий) или хуже (и происходит только в определенных случаях). Наконец, как указывает создатель современной версии эпигенетической теории эволюции М.А. Шишкин, понятия генотипической и фенотипической изменчивости вообще относятся к разным ситуациям. О генотипической изменчивости принято говорить, сравнивая развитие разных генотипов в одинаковых условиях, а о фенотипической — при сравнении развития одинаковых генотипов в разных условиях. Как показывают данные экспериментов, во всех случаях результат развития организма может быть предсказан лишь вероятностным образом, через описание набора возможных результатов развития и вероятности каждого из них.
Главной формой отбора с этой точки зрения оказывается стабилизирующий отбор (рис. 5.26.3). Пока условия среды соответствуют норме, особи, которые прошли в своем развитии по наиболее вероятному креоду, будут иметь максимальные шансы на выживание и оставление потомства. При этом неважно, какой вклад в нормальное развитие задан «наследственными» и «ненаследственными» признаками: оно определено всем эпигенетическим ландшафтом. «Движущий» отбор, как его представляют сторонники СТЭ, является выдумкой, никогда не регистрировавшейся в экспериментах. Отбор против нормы приводит к дестабилизации нормы и расширению диапазона изменчивости. В изменяющихся условиях этот эффект как раз позволяет «нащупать» те варианты развития, которые смогут стать новой нормой. При этом неважно, на какой основе будут развиваться эти новые варианты. Даже если их развитие соответствует классической схеме «модификаций» (всецело зависит от специфического воздействия среды), отбор на способность развивать такой фенотип приведет к стабилизации ведущего к нему онтогенетического пути.
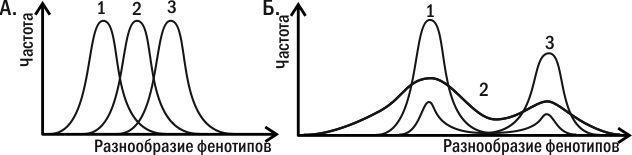
Рис. 5.26.3. Результаты отбора (на большее значение показанного по оси абсцисс признака) в соответствии с различными представлениями. А. Результат, ожидаемый с точки зрения синтетической теории эволюции (СТЭ). Отбор приводит к смещению нормы реакции. Б. Результат, ожидаемый с точки зрения эпигенетической теории эволюции (ЭТЭ). Отбор приводит к дестабилизации развития, а потом — к его стабилизации на новой норме. Регистрируемые в экспериментах результаты в большей степени соответствуют второму варианту
Дополнительные материалы:
Колонка: Вперед, к эпигенетическому будущему!
|
5.27. (дополнение) Давление на глубине: претерпевание и преодоление |