| ← | Дмитрий Шабанов | → |
| Королева проблем эволюционной биологии: кошмар Дженкина возвращается | Хватит ли восьми косвенных гипотез, описывающих преимущества полового размножения, чтобы объяснить возникновение пола? | Правда ли, что королева эволюционных проблем — это Красная (a.k.a. Чёрная) Королева? Не торопитесь с ответом — не забывайте о гермафродитах! |
| Колонка для Компьютерры #132 | Колонка для Компьютерры #133 | Колонка для Компьютерры #134 |
Парадоксальность полового размножения, о которой шла речь в моей предыдущей колонке, такова, что некоторые из читателей не смогли её осознать. Вдумайтесь: при половом размножении любой из генов особи передаётся потомку лишь с 50-процентной вероятностью, а при разных формах клонального воспроизводства — с вероятностью в 100%!
Давайте ещё раз рассмотрим тот сценарий, который обсуждает большинство авторов. Вид с клональным воспроизводством переходит к половому размножению (с самцами и самками). Как верно заметил Джон Мейнард Смит, такой переход может быть обусловлен только вдвое большей выживаемостью потомков от полового размножения, причём сразу же, в короткой перспективе. Это проще всего показать с помощью таблицы. Представьте себе популяцию, которая делится на две субпопуляции (части), A и B, различающиеся по характеру воспроизводства (классификация его типов дана в этой колонке).
|
«Соревно- |
Тип |
Произво- |
Потомков |
Выжива- |
Выживет |
Всего |
Результат |
|
Субпопу- |
Клональ- (тип I) |
Особь 1 |
100 |
25% |
25 |
50 |
Проигрыш |
|
Особь 2 |
100 |
25 |
|||||
|
Субпопу- |
Раздельно- (тип III) |
Самка |
100 |
51% |
51 |
51 |
Победа |
|
Самец |
— |
||||||
В показанном соревновании субпопуляция B вытеснит всех представителей субпопуляции A за относительно небольшое число поколений. Но обратите внимание: победа полового размножения в этом случае связана с невероятным, превышающим двукратный, ростом выживаемости потомков (вероятности для них дожить до какого-то сравнимого этапа их жизненного пути). Такой стремительный рост выживаемости должен быть связан с каким-то чрезвычайно весомым преимуществом! Неужели это — просто-напросто перетасовывание генов?
А если прирост выживаемости будет хотя бы немного меньшим? Давайте посмотрим.
|
«Соревно- |
Тип |
Произво- |
Потомков |
Выжива- |
Выживет |
Всего |
Результат |
|
Субпопу- |
Клональ- (тип I) |
Особь 1 |
100 |
25% |
25 |
50 |
Победа |
|
Особь 2 |
100 |
25 |
|||||
|
Субпопу- |
Раздельно- (тип III) |
Самка |
100 |
49% |
49 |
49 |
Проигрыш |
|
Самец |
— |
||||||
Если выигрыш в выживаемости потомства не дотягивает до двухкратного, клональные организмы в краткосрочной перспективе вытесняют раздельнополых. Долгосрочные преимущества полового размножения в такой ситуации просто не успеют проявиться!
Но почему же половое размножение распространено столь широко? Это требует особого объяснения, и в этой колонке я опишу его разные варианты. О «викарии из Брея», упомянутом в прошлый раз, вы, я надеюсь, помните.
Гипотеза «эгоистичного гена пола» Докинза — версия, которую читатели трёх последних колонок упоминали чаще всего.
"Трудности, с которыми сталкиваются теоретики, пытаясь объяснить эволюцию пола, по крайней мере отчасти связаны с тем, что по их представлениям индивидуум старается максимизировать число своих выживающих генов. В свете таких представлений половое размножение воспринимается как нечто парадоксальное. <...> Однако этот парадокс покажется менее парадоксальным, если в соответствии с приведёнными в моей книге доводами рассматривать индивидуум как машину выживания, создаваемую короткоживущей конфедерацией долгоживущих генов. <...> Eсли половое размножение как противоположное бесполому благоприятно для гена, определяющего половое размножение, то этого достаточно для объяснения существования последнего. Благоприятно ли оно для всех остальных генов данного индивидуума, не очень важно. С точки зрения эгоистичного гена половое размножение вовсе не такое уж странное явление". Ричард Докинз. Эгоистичный ген
Чтобы верно понять Докинза, нужно разобраться, что же он вкладывает в понятие «ген».
«...общепринятого определения гена не существует. <...> Я хочу воспользоваться определением, принадлежащим Дж. Уильямсу: ген — любая порция хромосомного материала, сохраняющаяся на протяжении достаточного числа поколений, чтобы служить единицей естественного отбора. <...> Ген — это репликатор с высокой точностью копирования». Ричард Докинз. Эгоистичный ген
Я б, конечно, не привязывал так однозначно понятие «естественный отбор» к уровню генов... С сохранением смысла подход Докинза можно выразить так: ген — фрагмент генетического текста, который передаётся из поколение в поколение как единое целое (что не исключает возможности его изменений). Посмотрим, существует ли «ген полового размножения» как нечто единое.
Нет! В позапрошлой колонке я не случайно потратил немало сил, объясняя, что половое размножение — феномен, связанный с гапло-диплоидным жизненным циклом с оплодотворением и мейозом. В этом эволюционном изобретении сошлись механизмы гомологической рекомбинации, достигшие совершенства в мейозе, а также системы, обеспечивающие сингамию и кариогамию (пояснение этих терминов — в той колонке). Объяснять возникновение полового размножения как возникновение какого-то «гена», который вдруг приобрёл такой эффект, — верить в сказку.
Что же, объяснение Докинза полностью ошибочно? Тоже нет. Вспомните (опять же из позапрошлой колонки) об инфекционной передаче «пола» у бактерий. Кишечная палочка, в цитоплазме которой находится небольшая кольцевая молекула ДНК, названная F-фактором, в силу этого приобретает способность формировать пили (структуры для инъекции ДНК) и передавать другим бактериям и F-фактор, и другие фрагменты ДНК. «Половая» особь заражает «полом» бесполую, воткнув в ту пиль. В данном случае объяснение Докинза оказывается как минимум вполне работоспособным. У организмов, обладающих гапло-диплоидным жизненным циклом с оплодотворением и мейозом, сама готовность особи к размножению — результат согласованной работы многих независимо возникших и эволюционировавших генов, каждый из которых сталкивается с 50-процентной вероятностью не попасть в следующее поколение. «Эгоистичного гена пола» у таких организмов просто не существует!
А каково же то преимущество, которое обеспечило триумф полового размножения? По этому поводу высказано немало гипотез, и несколько из них я хочу коротко описать. Подробнее эти гипотезы охарактеризованы в "Красной королеве" Мэтта Ридли, книге Вигена Геодакяна "Два пола. Зачем и почему?", а также в статьях К. Ю. Попадьина и И. А. Рухленко.
Храповик Мёллера. Герман Мёллер обратил внимание на то, что при клональной передаче геномов возникшие в них ошибки — мутации — могут быть исправлены только со смертью или отстранением от размножения их носителя. Если носители генотипов, свободных от ошибок, исчезнут из-за мутационного процесса или просто в силу случайности, самыми «чистыми» станут генотипы, несущие по одной мутации. Эта ситуация может сохрраняться какое-то время, но все равно рано или поздно закончится случайным исчезновением генотипов с одной мутацией. Тогда лучшими станут генотипы с двумя мутациями. «Безошибочный» генотип уже никогда не восстановится. Такое непрерывное ухудшение клональных геномов заставляет вспомнить о храповике — таком колёсике, из-за которого наручники могут только затягиваться, но не ослабляться.
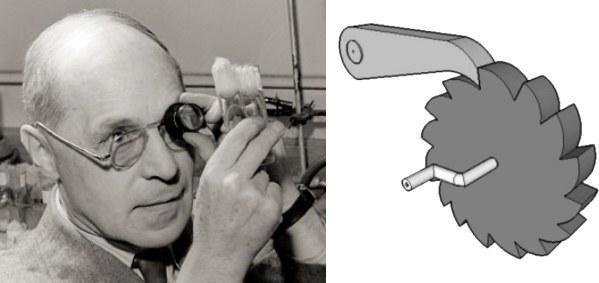
Слева — Герман Джозеф Мёллер (1890–1967). Справа — храповик: зубчатое колесо, которое может вращаться только в одну сторону (в показанном на рисунке случае — по часовой стрелке)
Храповик Мёллера, к примеру, отвечает за дегенерацию человеческой Y-хромосомы, и причина этого именно в том, что она передаётся из поколения в поколение клонально. А гипотеза, связанная с этим механизмом, заключается в том, что это он ограничивает возможности существования клональных видов, создавая преимущества для половых.
В результате действия храповика Мёллера подавляющее большинство видов, отказавшихся от рекомбинации, по большому счету обречено. Те, что имеют относительно небольшой геном, высокую численность и находятся под эффективным контролем отбора, вымирают относительно медленнее, но все равно их перспективы печальны. Прогрессивную эволюцию до происхождения полового размножения обеспечивали иные формы рекомбинации, а после возникновения пола развитие и просто сохранение в работоспособном состоянии генотипа оказалось связано с ним. Посудите сами: при скрещивании организмов, каждый из которых несёт по одной неблагоприятной мутации, четверть их потомков не унаследуют ни одной из них!
Гипотеза Наннея — это развитие идеи храповика Мёллера. Л. Нанней предположил, что все виды с половым размножением, в эволюции которых возникали бесполые клоны, были вытеснены этими клонами и вымерли, а позже (в результате работы храповика Мёллера) вымерли и эти клоны. Поэтому в результате группового отбора сохранились лишь такие виды с половым размножением, которые в силу каких-то причин утратили возможность перейти к клональному размножению.
А как же отказывающиеся от полового процесса дафнии, о которых я рассказывал в прошлой колонке? С точки зрения гипотезы Наннея они — на полпути к вымиранию...
"Топор Кондрашова" — мутационная детерминистическая гипотеза Кондрашова. Выдвинута российским (сейчас российско-американским) биологом Алексеем Симоновичем Кондрашовым. Эта гипотеза обращает внимание на то, как слабовредные по отдельности мутации распределяются в популяции организмов с половым размножением. Некоторые особи несут небольшое количество таких мутаций, некоторые — много. Предположим, что действие этих мутаций обладает накопительным эффектом: чем их больше, тем ниже жизнеспособность. Рассмотрим распределение особей по количеству несомых ими мутаций. Отбор будет отсекать, как топором, наиболее перегруженные классы особей, попросту удаляя «хвост» распределения. Каждая смерть особи из этих классов распределения уносит из популяционного генофонда множество мутаций; у бесполых организмов такая же очистка потребовала бы гибели многих особей.
Кроме того, Кондрашов обратил внимание на эволюционные последствия чередования гаплоидности и диплоидности, о которых мы говорили в прошлый раз. Летальная мутация у гаплоидного организма ведёт к его смерти. Диплоидный организм может в двух своих хромосомных наборах иметь несколько летальных мутаций и прекрасно себя чувствовать. С другой стороны, те гены, которые на гаплоидной фазе приводят к гибели (или отдельного гаплоидного поколения, или хотя бы гаплоидных половых клеток) будут эффективно устраняться из популяции, не допускаясь до диплоидной фазы.
В отличие от «викария из Брея» и храповика Мёллера, гипотеза Кондрашова касается краткосрочных преимуществ организмов, возникших в результате полового процесса.
Гипотеза полового отбора Мэннинга. Английский биолог Джон Мэннинг обращает внимание на то, что самки, выбирая самцов, могут проводить достаточно эффективную оценку приспособленности самцов. Это показано и для человека, и для многих других животных — впрочем, тех, кто характеризуется сложным поведением. Сомневаться в том, что этот механизм во многих случаях работает, не приходится. С другой стороны, этой гипотезой не объяснить переход к половому воспроизводству от клонального, когда тонкие механизмы полового отбора ещё не возникли.
«Эволюционную теорию пола» Геодакяна я бы назвал информационной гипотезой Геодакяна. Российский физик и генетик Виген Геодакян выдвинул ряд оригинальных идей, касающихся того, как информация о том, какие генотипические особенности особей оказываются востребованы средой, отражается в генофонде вида. Он убедительно показывает, что эволюционные новшества опробуются прежде всего на самцах (на представителях того пола, которого менее жалко). Конечно, о взглядах Геодакяна надо писать отдельно - и желательно подробно. Здесь скажу только то, что, на мой взгляд, у него есть целый ряд хороших идей, но и немало явно сомнительных суждений. И, конечно, я не могу согласиться с его интегральной оценкой результата его творчества.
"Широта теории, её объяснительный и предсказательный потенциал позволяют ей занять место многих частных и несовершенных теорий в области пола, в частности теории полового отбора Ч. Дарвина". В. Геодакян. Два пола. Зачем и почему. - С. 5.
В эту колонку поместятся ещё только лишь две экологические гипотезы, касающиеся краткосрочных экологических преимуществ организмов, возникших в ходе полового размножения.
Гипотеза лотереи Уильямса нравится мне особенно. По простой причине: я раньше дошёл до неё сам — и лишь потом прочитал о ней у Ридли и Геодакяна. Я думал о брекетинге экспозиции в фотографии. Когда фотограф не знает, подходит ли установленная им экспозиция к имеющимся условиям освещения и особенностям предмета съёмки, а также когда ему не нужно дорожить каждым кадром, он может использовать брекетинг. Один кадр автоматически делается с теми установками, которые выставлены в фотоаппарате, один — с увеличенной на ступень экспозицией, один — с уменьшенной. Экспозицию можно только увеличивать или уменьшать, а для изменения свойства потомков существует множество степеней свободы. Что надо менять? Те признаки, по которым родитель (прошедший горнило отбора) отличается от своих сородичей, также «проверенных» естественным отбором.
На первый взгляд, гипотеза Джорджа Уильямса хорошо согласуется с анализом жизненных циклов. Пока условия остаются постоянными, дафнии и тли размножаются клонально. Когда условия меняются, появляется половое поколение, повышающее разнообразие потомства. Когда земляничный куст или колония коралловых полипов осваивают своё ближайшее окружение, они делают это клонально: земляника выбрасывает усы, а кораллы почкуются. Зато отдалённые местообитания заселяются с помощью семян или личинок, образующихся при половом размножении. Организмы, которые ищут места для своих потомков лишь вдалеке (как вязы или устрицы), обходятся исключительно половым размножением.
Увы, считается что анализ распространения клональных и бесполых форм опровергает гипотезу Уильямса. Согласно ей, можно было бы предположить, что половое размножение будет эффективнее в суровой и изменчивой среде, а бесполое — в благоприятной и постоянной. В то же время известно, что к бесполому размножению чаще переходят виды, населяющие экстремальные местообитания.
Гипотеза заросшего берега Белла — результат развития гипотезы лотереи. Грем Белл исходил из того, что половое размножение выгодно там, где появляющиеся в избыточном количестве потомки конкурируют прежде всего друг с другом. Чем более похожи друг на друга особи, тем острее конкуренция между ними; емкость среды для идентичных организмов ниже, чем для разнообразных. Название этой гипотезы отсылает к прославившемуся финальному пассажу «Происхождения видов» Дарвина.
"Любопытно созерцать густо заросший берег, покрытый многочисленными, разнообразными растениями с поющими в кустах птицами, порхающими вокруг насекомыми, ползающими в сырой земле червями, и думать, что все эти прекрасно построенные формы, столь отличающиеся одна от другой и так сложно одна от другой зависящие, были созданы благодаря законам, ещё и теперь действующим вокруг нас". Чарльз Дарвин. Происхождение видов
Но гипотеза заросшего берега плохо согласуется с тем, что самые крупные и самые совершенные организмы, зачастую производящие немногочисленных потомков, размножаются почти исключительно половым путём.
Из гипотез, объясняющих преимущества полового размножения, о которых я хотел вам рассказать, не упомянута только одна — та, которую считают фавориткой многие авторитетные авторы. Это гипотеза Красной Королевы Ван Валена — Гамильтона. О ней надо рассказывать подробно, и в эту колонку её обсуждение не поместится никак. Ну что же — в следующий раз. Посмотрим, сможет ли она объяснить скачкообразное повышение выживаемости потомков вдвое...
| ← | Дмитрий Шабанов | → |
| Королева проблем эволюционной биологии: кошмар Дженкина возвращается | Хватит ли восьми косвенных гипотез, описывающих преимущества полового размножения, чтобы объяснить возникновение пола? | Правда ли, что королева эволюционных проблем — это Красная (a.k.a. Чёрная) Королева? Не торопитесь с ответом — не забывайте о гермафродитах! |
| Колонка для Компьютерры #132 | Колонка для Компьютерры #133 | Колонка для Компьютерры #134 |
Коментарі
_
Спасибо за колонки. Очень кратко и доходчиво.