|
|
А.П.Расницын. Теоретические основы эволюционной биологии // В.В.Жерихин, А.Г.Пономаренко, А.П.Расницын. Введение в палеоэнтомологию. М.: КМК. 2008. 371 с
1.1. ПРОЦЕСС ЭВОЛЮЦИИ
1.1.1. СИНТЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
1.1.2. ЭПИГЕНЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
1.1.2.1. Основные положения
1.1.2.2. Адаптивный компромисс
1.1.2.3. Проблемы
|
1. ТЕОРЕТИЧЕСКИЕ ОСНОВЫ ЭВОЛЮЦИОННОЙ БИОЛОГИИ
Эволюция и систематика насекомых в этой книге (и в последующих выпусках этой серии) реконструируется, исходя из предположений, иногда не слишком привычных для отечественных биологов, воспитанных на синтетической теории эволюции. Поэтому кажется полезным дать здесь краткое обоснование моих взглядов на эволюционный процесс и на способы изучения его результатов. Подробнее эта точка зрения изложена в других работах (Расницын, 1987, 2002, 2005; Rasnitsyn, 1996, 2006).
1.1. ПРОЦЕСС ЭВОЛЮЦИИ
Представления о полной и окончательной победе дарвинизма в современной биологии не вполне соответствуют действительности: альтернативные взгляды сохраняются и продолжают развиваться. Среди прочих свидетельств на это указывает, например, появление объемистой книги Ю.В. Чайковского (2003) с многообещающим названием «Эволюция». Поэтому прежде, чем разбирать конкурирующие парадигмы селектогенеза (того направления мысли, которое чаще обозначают неудачным термином дарвинизм, см. ниже), мне придется как-то обосновать выбор именно этого направления.
В качестве альтернативы селектогенезу предлагаются различные концепции. Чайковский (2003) называет четыре системы взглядов на эволюционный процесс: дарвинизм, номогенез, ламаркизм и жоффруизм (по имени одного из знаменитых ранних эволюционистов Э. Жоффруа Сент-Илера). Эта классификация неудачна и терминологически, и по существу. Именем основателя логично называть учения, суть которых состоит в толковании канонических текстов (христианство, буддизм, марксизм), но не системы научных взглядов, поскольку логика научного поиска почти неминуемо ведет к пересмотру большей или меньшей части утверждений прежних авторов. В результате порой возникают анекдотические ситуации. Тот же Ю.В. Чайковский при всем своем яростном антидарвинизме признает не только отбор (в сильно урезанном виде) и наследование приобретенных свойств (в сильно расширенном виде), но и пангенезис, и оказывается большим дарвинистом, чем те, кто считаются и считают себя последователями Дарвина. Поэтому вне этого раздела я буду по возможности избегать использования таких названий.
По существу предложенная классификация неудачна в том смысле, что ее подразделения перекрываются широко до неразличимости. Для номогенеза знаковым указано особое внимание к закономерному характеру эволюционного процесса, но как известно, эволюционные исследования всех направлений заняты поиском (небезуспешным!) таких закономерностей (см., например, Раутиан, 1988). Даже если сузить номогенез до признания широкой повторяемости эволюционных путей (параллелизмы, рефрены Мейена, и т.п.), то, как будет видно из дальнейшего, этот подход прекрасно совместим с селектогенезом, признающим важную роль отбора.
Жоффруизм определен как направление, придающее особое значение онтогенетическим преобразованиям как механизму эволюции: в равной мере это справедливо для эпигенетической версии селектогенеза (Уоддингтон - Шмальгаузен - Шишкин - Расницын - Тихомирова, см. ниже). Ограниченная эффективность отбора, признание которой Чайковский указывает как важную черту номогенеза, жоффруизма и ламаркизма, также относится к важнейшим положениям эпигенетического селектогенеза (см. ниже).
С самим ламаркизмом положение несколько более сложное. С этим термином связывают два существенно независимых утверждения — о прогрессивном характере эволюционного процесса (усложнение и совершенствование организмов, их строения и функционирования) и о наследовании приобретаемых свойств. Проблема прогресса естественно входит в проблематику селектогенеза (опять сошлюсь на свою работу: Расницын, 1971). Наследование приобретаемых свойств долгое время считалось несовместимым с дарвинизмом (хотя сам Дарвин считал иначе), но когда с открытием ретровирусов была доказана обратная связь от фенотипа к генотипу минуя отбор, оказалось, что ничего серьезного для селектогенеза не произошло. Действительно, если организму выгодно, скажем, закрепить в ДНК определенную модификацию и если возможно выработать механизм такой фиксации (например, с участием ретровирусов), почему отбор не может реализовать такой механизм? Другое дело, что ДНК на то и существует, чтобы надежно хранить информацию и, соответственно, по возможности исключать внешние воздействия на ее структуру, поэтому всякий механизм, направленно осуществляющий такое воздействие, опасен для осуществления основной функции ДНК. Именно с этим, на мой взгляд, связан тот факт, что наследование приобретаемых свойств не относится к числу рутинных механизмов эволюционного процесса.
Таким образом, ламаркизм, жоффруизм и номогенез в приведенной выше трактовке не составляют принципиальной альтернативы селектогенезу. В то же время всем понятно, что по крайней мере ламаркизм и номогенез — это нечто совершенно иное, чем современный дарвинизм (т.е. селектогенез), просто их основополагающее (сущностное) отличие пока не сформулировано. Что касается жоффруизма, он описан недостаточно для того, чтобы поместить его по какую-то сторону водораздела, о котором пойдет речь.
По моему убеждению, сущностное различие между селектогенезом с одной стороны, ламаркизмом и номогенезом с другой, состоит в отношении к источнику целесообразности. Целесообразной (приспособительной, адаптивной) называют структуру (в том числе систему действий), которая выглядит, по крайней мере внешне, как созданная специально для выполнения определенной, в некотором контексте полезной функции. Поэтому объяснение происхождения целесообразности предполагает учет этой будущей функции возникающей структуры (системы): в определенном смысле будущая функция формирует структуру.
Возникновение существенно новой целесообразной структуры (системы) мыслимо лишь путем прямого целеполагания, т.е. с участием разума (грубо говоря, человека или бога), либо путем запоминания и воспроизведения удачного выбора, т.е. путем отбора. Эффективность отбора как метода поиска целесообразного решения продемонстрировано У. Эшби (1964) на модели гомеостата. Для приложения к эволюционному процессу эта модель была модифицирована как параллельный гомеостат, отличающийся тем, что ступенчатые переменные используются не последовательно, как у Эшби, а одновременно: в их качестве выступают фенетически и генетически различающиеся особи (Расницын, 1971). Другим ярким примером параллельного гомеостата является, по современным представлениям, система иммунитета теплокровных, как она описана Чайковским (2003) на с. 189-190.
Бесспорным механизмом принятия целесообразных решений является разум. Также бесспорно, что разум человеческого типа возникает поздно и источником эволюционной целесообразности быть не может. Поскольку существенная целесообразность эволюции никем, по-видимому, не отрицается, речь может идти о разуме, либо имманентном жизни, либо предсуществующем ей, т.е. о внеестественной (грубо говоря, божественной) природе эволюционной целесообразности. Именно здесь, на мой взгляд, лежит водораздел между селекционизмом, отказывающимся рассматривать эволюцию как продукт разумной деятельности, и номогенезом и ламаркизмом, предполагающими внеестественный (альтернативный отбору) источник прогресса (ламаркизм) или целесообразности (номогенез). Поскольку привлечение внеестественных факторов эволюции, на мой взгляд, законно лишь в рамках веры, а с наукой несовместимо, выбор селектогенеза для меня неизбежен.
В современной теории селектогенеза конкурируют два главных подхода — синтетический и эпигенетический. Синтетическую теорию эволюции, лучше знакомую нашим биологам, можно охарактеризовать как редукционистскую, сводящую эффекты на уровне целого к явлениям на уровне его элементов — прежде всего элементарных генетических факторов и их частот в популяциях. Противостоящую ей эпигенетическую теорию можно лишь очень условно обозначить как антиредукционистскую: ее приверженцы пытаются понять свойства эволюционного процесса, исходя из некоторых свойств целого организма, прежде всего — его онтогенеза. По существу это дальнейшее развитие холистического подхода к эволюции (Smuts, 1926, цит. по Smuts, 1987). Взаимоотношения синтетической и эпигенетической гипотез специально рассмотрены М.А. Шишкиным (1987, 1988а, 2006) и Д.Л. Гродницким (2000, 2002). Опираясь на их результаты, попытаюсь изложить свое видение этой проблемы.
1.1.1. СИНТЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
Синтетическая, а более точно — популяционно-генетическая теория эволюции (ибо синтез там скорее провозглашен, чем достигнут) рассматривает наследственность, т.е. способность к устойчивому воспроизведению фенотипа и его признаков в последовательных поколениях, как свойство особых элементарных носителей- генов, ныне отождествляемых с участками (локусами) хромосом и далее с участками ДНК (цистроны и т.п.). Соответственно эволюционный процесс представляется как динамика частот аллелей в популяциях, контролируемая отбором (дифференциальным воспроизводством фенотипов). Признаки фенотипа возникают и изменяются в результате мутаций и рекомбинаций генов. Поскольку все эти изменения не обладают свойством изначальной целесообразности, именно отбор формирует состав популяций и свойства организмов, и делает он это с точностью, ограниченной лишь стохастическими (вероятностными) факторами. Кроме фактора случайности, единственным препятствием полному соответствию организации живых существ требованиям отбора является обмен генами, который унифицирует структуру популяций на уровне, отвечающем не локальным, а усредненным характеристикам отбора.
В соответствии с этими взглядами организация живых существ представляется мягкой глиной в руках отбора, набором признаков, свободно тасуемых отбором. Соответственно, эволюционный процесс с данной точки зрения оказывается равномерным — по крайней мере настолько, насколько равномерны вариации среды и отбора. Равномерность эволюции устойчиво нарушается лишь дивергенциями и вымираниями, каковые в рассмотренной системе взглядов и представляют единственную естественную основу для построения системы организмов.
Синтетическая теория, конечно, признает и плейотропность генов (влияние одного гена на несколько признаков), и полигенность признаков (множественную генетическую обусловленность отдельного признака), так что каждый признак в действительности зависит от всего генома. Однако эти признания в действительности декларативны, так как тотальная взаимозависимость элементов структуры и функций организма и, соответственно, отсутствие сколько-нибудь постоянного вклада гена в приспособленность делают невозможной сведение эволюционного процесса к элементарным генетическим, а тем более молекулярно-генетическим событиям.
Эти упрощения не безобидны: они ведут к несоответствию между предсказаниями теории и результатами наблюдений. Приведем некоторые из таких несоответствий. Прежде всего обратимся к темпам эволюционного процесса, оцениваемым по скорости возникновения и вымирания таксонов того или иного ранга. Таксон — единственное понятие, самым смыслом существования которого является попытка интегрированной оценки сходств и различий организмов. Однако на деле сопоставимость таксонов одного ранга в разных группах не обеспечена ничем, кроме интуиции систематиков (о чем речь ниже), поэтому такой метод оценки скорости эволюции несовершенен. Но других способов сравнения темпов эволюции организмов у нас просто нет. Сколь ни несовершенна (субъективна) оценка различий в таксономических рангах, она более информативна, чем более объективная оценка различий с помощью признаков. Хорошо известно, что внешне одинаковые различия могут достигаться настолько разными способами и оказываются проявлениями настолько разномасштабных преобразований организации, что оценка темпов эволюции с их помощью будет только вводить в заблуждение. Особенно ярко подтверждают этот вывод материалы по эволюции в островных условиях и на ранних этапах эволюции таксонов (феномен архаического многообразия). Здесь быстро и легко возникают гротескные формы с несбалансированной организацией. Они часто отличаются по таким признакам, которые в норме (в стабилизированных группах) характеризуют таксоны гораздо более высокого ранга (подробнее см. ниже). В то же время палеонтологическая оценка темпов таксономической эволюции дает результаты и обнаруживает тенденции, которые вряд ли можно списать на несопоставимость таксонов одного ранга в разных группах (Расницын, 1987).
Согласно синтетической теории эволюционный процесс управляется в основном теми же факторами и механизмами, которыми оперирует генетика популяций. Но тогда и скорость макроэволюции должна подчиняться тем же закономерностям, что скорость изменения частот генов в генетике популяций. Эта скорость в свою очередь определяется мощностью потока адаптивно компетентных генетических вариаций (изменений, значимых для отбора), которые протекают через эволюционирующую совокупность. Следовательно, скорость должна быть тем выше, чем выше частота мутаций и скорость смены поколений, чем больше размер популяций и мобилизационный резерв изменчивости.
Палеонтологический материал не позволяет прямо оценить большинство перечисленных параметров, используемых в генетике популяций. Некоторые из них все же могут быть косвенно оценены у ископаемых — в той мере, в какой они коррелируют с другими признаками, такими как размеры, таксономическая принадлежность и т.п. В частности, если генетический подход к эволюции справедлив, то ее скорость должна быть минимальной у млекопитающих, обладающих сравнительно медленной сменой поколений и небольшими популяциями (в связи с относительно крупными размерами особи), максимальной у одноклеточных и промежуточной у беспозвоночных. В действительности же, как показывают многочисленные палеонтологические данные, эти соотношения имеют обратный характер (Расницын, 1987). При этом различия весьма велики даже на уровне вида, а ведь эта таксономическая категория считается сравнимой в разных группах. Так, возраст полусовременной фауны (фауны, в которой половина видов современные, половина — вымершие) у крупных млекопитающих (хоботные и копытные) 0,2 миллиона лет , у мелких 0,5, у птиц и рыб 0,7, у насекомых 3-7, у моллюсков 3,5-5, у диатомовых водорослей — 15 миллионов лет. Время полувымирания (по аналогии с полураспадом, время вымирания половины исходного числа видов) наименьшее у слонов (0,18 млн. лет), в среднем у млекопитающих 0,54, у костистых рыб 3,5, у граптолитов 1,3, иглокожих 4,2, двустворчатых моллюсков 7, у планктонных фораминифер 5, бентосных 18-24, у диатомовых водорослей 5,5, у динофлагеллат 9 млн. лет. На старших таксономических уровнях различия подобные, но часто более резкие. Например, возраст полусовременной фауны для родов млекопитающих 4 млн. лет, птиц 10, рептилий 20, рыб 30-50, насекомых 40, моллюсков 60, фораминифер 230 млн. лет.
Приведенные цифры не ставят под сомнение генетические предсказания. Отбор абсолютно автоматичен: если в популяции появляются особи, различающиеся по степени приспособленности, они будут неуклонно отбираться в полном соответствии с положениями генетики популяций. В той мере, в какой эти различия наследуемы, разное участие особей в воспроизводстве популяции столь же автоматически приведет к изменению ее наследственной структуры. Значит, предсказываемое генетическими теориями распределение скоростей эволюции обязательно должно иметь место в природе — в той мере, в которой мощность общего потока генетических вариаций в долговременном аспекте коррелирована с мощностью потока отбираемыгх (повышенно адаптивных) наследуемых изменений. Подтверждением служат хорошо известные различия по типичным скоростям селекции между породами домашних животных, сортами растений и штаммами микроорганизмов. Не менее показательны и изредка наблюдающиеся максимальные темпы эволюции насекомых, когда на острове Гавайи возрастом 0,5 млн. лет. (Rotondo et а1., 1981) в результате 17 вселений возникло 46 видов жука усача рода Plagithmysus (Gressit, 1978), т.е. не менее одного акта видообразования происходило в среднем за 150— 200 тыс. лет. Г.Х. Шапошников почти вывел новый вид тли всего за один сезон (см. ниже).
В то же время очевидно, что в целом распределение скоростей эволюции на уровне вида и выше в первом приближении противоположно предсказанному популяционной генетикой. Это означает, что популяционно-генетические ограничения не являются таковыми для эволюционного процесса. Грубо говоря, поток общей генетической изменчивости эволюционно избыточен даже в популяциях крупных млекопитающих, не говоря уже о других, более многочисленных и быстрее плодящихся организмах. Скорость изменений на надпопуляционном уровне (скорость эволюции) должна регулироваться какими-то другими факторами. Вопрос о том, какие это могут быть факторы, рассмотрим позже. Здесь можно только заключить, что тот уровень, к которому синтетическая теория эволюции пытается свести эволюционный процесс и тем самым его объяснить, не отвечает возлагаемым на него надеждам и не дает ожидаемых объяснений — по крайней мере в отношении парадокса эволюционных скоростей.
Существуют и другие группы данных, не укладывающиеся в рамки синтетической теории эволюции. Если организация живых существ — мягкая глина в руках отбора или набор признаков, свободно тасуемых отбором, то ее результаты должны быть предсказуемыми в той мере, в какой известны условия, в которых протекала эволюция. Антиселекционисты показали на многочисленных примерах, что это не так (см., например, Любищев, 1982). Даже близкородственные формы в сходных, казалось бы, условиях, могут вести себя совершенно по-разному. Самый парадоксальный пример этого — крайне экстравагантная в репродуктивной сфере пятнистая гиена и вполне ординарная в этом отношении близкая к ней полосатая гиена.
Еще менее объяснима с синтетической точки зрения широко распространенная, если не универсальная дискретность биологического разнообразия. В этой системе взглядов естественна лишь одна форма дискретности — дискретность амфимиктического вида в условиях, когда обмен генами достаточно интенсивен для унификации состава популяций вопреки локальным вариациям отбора. Но тогда между степенью (полнотой и древностью) изоляции и степенью дивергенции должна существовать хорошая корреляция. Этого в действительности не наблюдается: тормозящее (нивелирующее различия) влияние обмена генами на скорость дивергенции подтвердить не удается. К такому выводу пришел, в частности, С.С. Шварц (1980), сравнивавший виды млекопитающих с разной склонностью образовывать изоляты. Напрашивающийся контрпример островного видообразования в действительности относится к иному контексту (см. ниже).
Совершенно необъяснима с точки зрения синтетической теории эволюции дискретность видов у партеногенетических и бесполых организмов, лишенных обмена генами. Сравнение близких обоеполых и партеногенетических видов у коловраток (Майр, 1974) и жуков-долгоносиков (Иванова, 1978; В.В. Жерихин, личное сообщение), обоеполых и бесполых видов у протистов (Ю. Полянский, 1957; Ромашку, 1977), низших водорослей (В. Полянский, 1956) и папоротников (Fаггег, 1990) показало, что однополые и бесполые виды столь же дискретны, как и обоеполые.
Не менее показателен длительный (миллионы, десятки и возможно даже сотни миллионов лет) эволюционный стазис на видовом и родовом уровне, когда об обмене наследственной информацией не может быть и речи. Так, популяции четырех или пяти из тридцати австралийских видов бессяжковых насекомых (Ргоtuга) на видовом уровне неотличимы от популяций с далеких материков и островов — Калимантан, Япония, Южная Африка, Европа (Тихеп, 1967). Бессяжковые не стойки к высыханию и не покидают почву, поэтому трансконтинентальные миграции для них практически исключены. Объясняя их распространение, трудно избежать ссылок на дрейф континентов, тем более что для бессяжковых известен и классический «дрейфовый» ареал (Delamarentulus tristani Silv. распространен на обоих побережьях Атлантического океана, на Коста-Рике и в Западной Африке; Тихеп, 1963). Но из этого автоматически следует возраст вида, оцениваемый десятками миллионов лет.
Известны и более прямые указания на большую древность некоторых видов. Так, в фауне эоценового балтийского янтаря (возраст не менее 40 млн. лет) сейчас известно несколько видов насекомых и около десятка видов клещей, на видовом уровне неотличимых от современных; еще более обычно персистирование в течение 10-20 млн. лет (Zherikhin, 1999). Находки разнообразных плиоценовых галлов, неотличимых от галлов, вызываемых современными видами насекомых на тех же растениях, гораздо моложе (3-5 млн. лет), но они не менее важны (Zherikhin, 2002а). Морфологическая специфика галла обусловлена биохимическим воздействием насекомого-галлообразователя на ткани растения-хозяина. Поэтому морфологическая стабильность галлов доказывает биохимический стазис насекомых в течение миллионов лет. Многие из этих находок происходят из Северной Европы, так что речь идет о видах, переживших ледниковый период (правда, в основном за счет миграций), но не изменившихся даже биохимически. Таким образом, стазис длиной в миллионы лет осуществляется не только в отсутствие обмена генами, но и вопреки несомненным глубоким, возможно даже катастрофическим, изменениям условий.
Рекордсменом, однако, является щитень Triops cancriformis (Schaffer), который практически не изменился с раннего триаса (около 230 млн. лет), и даже вопрос о видовой самостоятельности пермских популяций остается нерешенным (Tasch, 1969). Этот вывод подтверждают данные молекулярных часов для двух морфологически едва различимых японских популяций другого вида, T. longicaudatus (LeConte): они свидетельствуют о расхождении видов около 15 млн. лет назад. Для других видов этого рода, включая T. cancriformis, дивергенции датируются 25-45 млн. лет (Suno-Uchi et al., 1997).
Не более понятен в этой системе взглядов обычный феномен, когда старшие таксоны оказываются более дискретными и демонстрируют более ясные родственные отношения, чем виды. Действительно, виды расходились относительно недавно, имели меньше времени на дивергенцию, чем старшие таксоны и, следовательно, должны были сохранить больше следов того, как именно протекала дивергенция (подробнее см. ниже).
Все сказанное, на мой взгляд, достаточно убедительно показывает серьезное несоответствие между популяционно-генетическим подходом с его производными (синтетической теорией эволюции и биологической концепцией вида) и результатами наблюдений. Поэтому пора уже перейти к другой системе взглядов на эволюционный процесс.
1.1.2. ЭПИГЕНЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
1.1.2.1. Основные положения
В отличие от синтетической, эпигенетическая теория видит эволюционный процесс прежде всего как процесс эволюционного преобразования онтогенеза (Шишкин, 1987, 1988а, б, 2006; Раутиан, 1993). При этом особое внимание обращается на целостность онтогенеза в том смысле, что и сам процесс, и его результат (строение организма на последовательных этапах его развития) гораздо устойчивее, чем любые отдельные факторы и процессы развития. Нормальное развитие эквифинально и способно релаксировать (подавлять, поглощать, нивелировать) очень широкий круг воздействий и возмущений, как внешних, так и внутренних, включая результаты различных ошибок и нарушений нормального развития.
Таким образом, наследуется (устойчиво воспроизводится в последующих поколениях) нормальный онтогенез (норма реакции) в целом, а не отдельные признаки. Помимо нормы, существует масса разнообразных уклонений (аберраций) развития с неустойчивым воспроизведением, которые в нормальных условиях реализуются редко. В неблагоприятных условиях, когда механизмы защиты нормы удается нарушить или преодолеть — в результате ли сильного внешнего воздействия или по внутренним причинам (ошибки развития), — развитие идет по аберрантному (уклоняющемуся и неустойчивому) пути. Поскольку эндогенные аберрации развития легко возникают не только при половом, но и при партеногенетическом и бесполом размножении, их возникновение есть реализация скрытой гетерогенности популяции, а не возникновение de novo каких-то генетических изменений.
Эпигенетическая теория рассматривает аберрации как материал отбора, который способен создать на ее основе новую адаптивную норму. Такой процесс легче описать в рамках метафоры эпигенетического ландшафта. Нормальный онтогенез представляет собой устойчивую (стабилизированную) последовательность тесно связанных отдельных эпигенетических процессов. Более или менее обособленные отрезки этой последовательности именуют креодами. Если представить себе креод в виде сильно заглубленной долины в некотором ландшафте, по уклону которой течет онтогенетический процесс (рис. 1), то аберрациями будут пологие боковые долинки, приподнятые в бортах главной. Чтобы аберрация реализовалась, т.е. чтобы онтогенез мог выйти в боковую долину, нужно либо сильное внешнее воздействие на развивающийся организм, выталкивающее его на боковую долину (рис. 2а), либо изменение самого ландшафта, выполаживающее дно долины (дополнительное аберративное изменение, рис. 2б), либо то и другое вместе (рис. 2в).
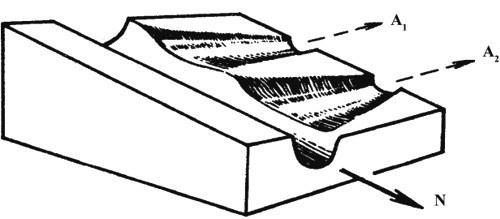
Рис. 1. Участок эпигенетического ландшафта, показывающий перепады высоты стенок (порогов устойчивости) креода в местах ответвления аберративных долин. А1, А2 - аберративные долины, N - главная долина (креод) (по Шишкину, 1987)
Аберрации мало устойчивы («не наследственны»; более устойчивые из них часто именуются модификациями), но они постоянно, хотя и непредсказуемо в деталях, воспроизводятся в популяции, поскольку эпигенетический ландшафт с соответствующими боковыми долинками — это адаптивная норма вида. Поэтому, несмотря на свою слабую наследуемость, именно аберрации служат материалом отбора. Если аберрация оказалась полезной, отбор будет избирательно сохранять те эпигенотипы, которые более устойчиво ее воспроизводят. Другими словами, преимущество получат те эпигенотипы, где соответствующая долинка углублена, и уровень ее дна на стыке с главной долиной более приближен ко дну последней. Если при этом еще и главная долина будет выполаживаться ниже развилка (т.е. если понизится устойчивость прежней нормы), прежняя аберрация будет иметь шанс стать новой нормой. Если же прежняя норма сохранит свое значение, то стабилизируются обе нормы и будет выработан механизм онтогенетического переключения между ними. В случае когда этот механизм использует для переключения средовой сигнал, мы получаем типичную модификационную изменчивость. Там, где используется генетический сигнал (например, рекомбинация), мы получим одну из форм менделирования. Таким путем менделирующие признаки возникают и в природе (например, обычные механизмы определения пола), и в лаборатории (например, в процессе стабилизации чистых линий).
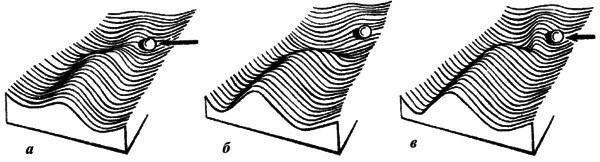
Рис. 2. Соотношения между строением эпигенетического ландшафта и характером повреждающего воздействия: а - уклонение развития на боковую долину за счет сильного внешнего воздействия (длинная стрелка); б- такое же уклонение под действием сильной мутации, вызывающей нарушение креода; в - промежуточное состояние (по Шишкину, 1987)
В целом для эпигенетической теории эволюции зафиксированный («запомненный») в адаптивной норме спектр возможных путей развития (креодов и их аберраций), т.е. эпигенетический ландшафт — это сущность живого организма, его онтогенетический потенциал, то, что определяет существование организма и с чем работает отбор. Чисто генетический уровень (гены и их аллели, динамика их частот в популяции, мутации, рекомбинация и т.п.) лежит гораздо глубже и не определяет специфики эволюционных процессов. Точно так же из процессов и закономерностей квантово-механического уровня нельзя вывести специфики того, что происходит на макроуровне. «Гены приходят и уходят, а креоды остаются»: хорошо известно, что признаки более устойчивы, чем гены и аллели, будто бы их определяющие.
Более подробно онтогенетические процессы в аспекте наследования рассмотрены М.А. Шишкиным (1987, 1988а, 2006). Для нас же более важна другая сторона проблемы эволюции онтогенеза.
1.1.2.2. Адаптивный компромисс
1.1.2.2.1. Формулировка и обоснование
Целостность организации живых существ оказывает существеннейшее влияние на характер их эволюции. На морфогенетическом уровне это влияние проявляется в том, что из-за глубокой взаимозависимости между изменениями разных креодов и, как следствие, между изменениями разных свойств и признаков организма крео- ды оказываются высоко стабилизированными. Они с большим трудом поддаются изменению за пределы нормальной внутривидовой изменчивости, которая сама уже стабилизирована отбором предыдущих поколений. Поэтому эволюция, т.е. успешныш выход за пределы нормального, стабилизированного эпигенотипа, происходит с трудом, нечасто и с мало предсказуемым результатом.
Другими словами, в руках отбора организация живых существ оказывается хрупким, капризным материалом, эволюционные преобразования — более или менее скачкообразными, а возникающее в результате биоразнообразие — более или менее дискретным. Эта дискретность не абсолютна, и даже там, где она существует, нам не всегда удается ее легко нащупать (из-за перекрывания пределов стабилизированной изменчивости, см. ниже). Но она существует и очень широко распространена, что биологи и особенно систематики не слишком склонны подчеркивать, сосредоточившись на исключениях и трудных случаях. Существование же исключений вполне естественно. Стабилизированный эпигенотип (адаптивная норма) включает в себя определенный, более или менее широкий спектр изменчивости, точно так же возникшей в результате отбора предшествующих поколений, как и модальные характеристики фенотипа. Спектры нормальной изменчивости близких видов вполне могут перекрываться, создавая проблемы для систематиков, даже если сами эпигенотипы при этом различаются дискретно.
Проблема в том, что мы пока не знаем, как увидеть, очертить и измерить сам по себе эпигенотип, а не только его внешние проявления. Эпигенотип недоступен прямому наблюдению, но он почти так же надежно выводится из имеющихся данных, как атом или нейтрино. А недоступен прямому наблюдению он из-за того, что эпигенотип — это очень сложная система взаимосвязей и взаимодействий. Но подобно атомному заряду, эпигенотип существенно определяет свойства и поведение его воплощений — особей, видов или иных таксонов. Поэтому его можно даже определить как материализованную сущность всех этих воплощений, что является в некотором смысле возвратом к эссенциализму — но к эссенциализму существенно иного толка, чем у Платона, поскольку сущность здесь признается материальной и познаваемой.
Однако вернемся к эволюции эпигенотипа. Высокая связность эпигенотипа и забуференность его эволюционных изменений удобно анализировать в рамках еще одной метафоры — метафоры адаптивного компромисса (Расницын, 1987). Действительно, устойчивость сбалансированного эпигенотипа можно рассматривать как одно из следствий теории систем, в частности, того ее утверждения, что ни одна система не может быть оптимизирована одновременно более чем по одному параметру. Не случайно, что стриж не умеет бегать и плавать, пингвин — бегать и летать, а страус не летает и едва ли умеет хорошо плавать.
Оптимизация реальных систем возможна лишь как нахождение компромисса между противоречивыми требованиями оптимизации различных параметров. Для живых существ, организацию которых пронизывают разнообразные корреляции и взаимозависимости, компромисс между различными адаптивными функциями должен быть особенно напряженным. Поэтому устойчивый эпигенотип должен быть организован по принципу глубоко проработанного компромисса между противоречивыми потребностями максимальной оптимизации всех адаптивных функций.
Отношение к живому существу как к адаптивному компромиссу не является чем-то новым. Э. Майр имел в виду, по-видимому, именно его, когда писал, что «эпигенотип вида, его система канализаций развития и обратных связей часто столь хорошо интегрирована, что с замечательным упорством противостоит изменениям» (1974, с. 353). Значительно раньше близкие идеи борьбы (конкуренции) частей в организме высказывали В. Ру и А. Вейсман (Вейсман, 1905), до них в виде принципа компенсации или уравновешивания — И.В. Гете и Ж. Сент Илер (см.: Дарвин, 1991, с. 128), а еще раньше как принцип экономии — Аристотель (1937).
Метафора адаптивного компромисса важна тем, что из нее можно вывести широкий круг сравнительно легко проверяемых следствий. Наиболее общее из них — трудность изменения хорошо сбалансированной организации. Конечно, эта трудность не абсолютна, и в той мере, в какой изменения осуществимы, утрата адаптивного значения какой-либо функции будет вести к редукции обеспечивающих ее систем. Утрату приспособления, потерявшего свое значение, естественно объяснить тем, что редукция или дезинтеграция ставших ненужными систем позволяет дополнительно оптимизировать другие системы, сохранившие адаптивность.
Для нас все же более существенна сама трудность перестройки однажды сформированного компромисса, существование барьеров на пути изменения сложившейся организации. По-видимому, эволюционный ландшафт не следует представлять так, как его обычно изображают: в виде адаптивных пиков, разделенных долинами пониженной приспособленности и соединенных хребтами, по которой эволюционирующая группа может постепенно двигаться по направлению от исчезнувшего пика к сохранившемуся. Более правдоподобной выглядит модель Ф.Р. Шрама (Shram, 1983) — система лунок, разделенных барьерами адаптивной неустойчивости.
Трудность преодоления устойчивости однажды достигнутого удачного компромисса предопределяет многие важные свойства эволюционного процесса: уже упоминавшиеся неравномерность и заторможенность (забуференность) эволюции, дискретность живых существ, малую эволюционную эффективность элиминации. Многие из вопросов, на которые синтетическая теория не может дать вразумительного ответа, такие как неравномерность эволюции, широкое распространение эволюционного стазиса, отсутствие видимого влияния популяционно-генетических факторов на скорость эволюционного процесса — все эти эффекты оказываются естественными и ожидаемыми при рассмотрении процесса с позиции адаптивного компромисса. Однако, как и следовало ожидать, их место занимают другие проблемы, не менее трудные для разрешения.
Эволюция идет, несмотря на все барьеры, но ценой вымирания множества групп, не сумевших эти барьеры успешно преодолеть. Попытаемся понять, в каких условиях и каким образом это происходит, как сбалансированный эпигенотип может быть преобразован в другой, также сбалансированный. Традиционно постулируемые механизмы эволюционного процесса, основанные на взаимодействии внутри- и межпопуляционного полиморфизма с изоляцией, не дают ответа на поставленный вопрос. Убедительно показано, что репродуктивная изоляция не только не необходима для эволюции, но даже не стимулирует ее (Гриценко и др., 1983, гл. 7). По всем законам генетики популяций, универсальность внутри- и межпопуляционного полиморфизма в сочетании с чудовищным прессом нормальной элиминации (см. ниже) должна вести к постоянной и быстрой эволюции с характерным распределением скоростей (высоких у низших организмов и низких у высших, см. выше). Этого также не наблюдается.
Из рассмотренного противоречия я могу сделать единственный вывод: в нормальных условиях внутривидовой полиморфизм не содержит эволюционно компетентных признаков. Проще говоря, ни один из вариантов фено- и генотипической организации, регулярно реализующихся в популяциях вида, в нормальных условиях не имеет эволюционно значимого преимущества перед другими вариантами. Все звери равны, и нет среди них более равных. Как же это может быть, если полиморфизм порой затрагивает, казалось бы, жизненно важные органы и структуры? Ответ, видимо, все тот же: все имеющиеся варианты потому и имеются, что согласуются с наличной эпигенетической системой, т.е. не разрушают ее стабильности, но они же и не способны создать новую стабильность (какую-то другую, не обязательно лучшую, но иную стабилизированную систему). Другими словами, внутривидовой полиморфизм вместе с модификационной изменчивостью на своем, надорганизменном уровне выполняет ту же функцию стабилизации системы в непредсказуемо изменчивой среде, что онтогенетическая регуляция, физиологические реакции и поведение выполняют на уровне организма.
Поскольку анализ отдельных факторов не дает ответа на вопрос о конкретном механизме, обеспечивающем эволюционный процесс, попытаемся подойти с другой стороны. Проанализируем те условия, в которых эволюция реально идет, причем идет относительно быстро.
Хотя репродуктивная изоляция, как уже говорилось, в нормальных условиях не является важным эволюционным фактором, хорошо известно, что в условиях изолированных островов, водоемов и т.д. эволюция идет особенно быстро и часто приводит к образованию сильно измененных, даже гротескных форм. С.С. Шварц (1980), специально анализировавший этот парадокс, пришел к выводу, что причина состоит в пониженной напряженности конкурентных отношений в бедных, незаполненных (в эволюционном масштабе времени) островных биоценозах. Конкретно эволюционная роль незаполненности ценозов состоит, по Шварцу, в том, что ослабленная конкуренция делает возможной быструю одностороннюю специализацию. Это привлекательная интерпретация для концепции адаптивного компромисса: предковый вид, попадая с материка в обедненный островной биоценоз, оказывается в условиях, смягченных по многим параметрам и допускающих дополнительную оптимизацию функций, оставшихся под жестким контролем среды, за счет других, не контролируемых отбором столь же строго. На самом деле, как мы увидим, дело сложнее и не сводится к ослаблению отбора, но пока воспользуемся этим нестрогим термином.
Изложенная модель эволюции на островах интересна еще и тем, что она соответствует концепции инадаптивной эволюции по В.О. Ковалевскому (Расницын, 1986). Более важно, однако, что она демонстрирует достаточно реальный механизм, способный преодолеть, сломать устойчивость хорошо сбалансированного адаптивного компромисса, тормозящую эволюцию. Естественно возникает вопрос, не может ли подобный механизм действовать не только при заселении островов, но и в каких-то других ситуациях.
Рис. 3. Корреляция между вымиранием и возникновением у животных: доля семейств, найденных в палеонтологической летописи: А - в последний раз, Б - в первый раз (по Newell, 1967, из Гранта, 1980)
Чтобы ответить на этот вопрос, рассмотрим некоторые другие случаи перехода группы в похожие условия — не будут ли и там наблюдаться ускорение эволюции и ее сходство с инадаптацией, т.е. увеличение размаха изменений в сочетании с их односторонностью, несбалансированностью. Переход в относительно свободное, незаполненное экологическое пространство характерен, например, для ранних этапов эволюции многих таксонов, и палеонтологическая летопись действительно подтверждает, что на ранних этапах эволюции таксона увеличивается как частота и масштаб эволюционных изменений, так и интенсивность отмирания вновь образующихся инадаптивных групп по мере заполнения ниши. Известно, что массовые вымирания и диверсификации часто происходят почти одновременно. Более того, периоды диверсификации следуют за периодами вымирании (рис. 3), что подтверждает тот факт, что освобождение экологического пространства вымершими группами стимулирует эволюцию сохранившихся. Ускорение эволюции на ранних этапах истории таксона более конкретно демонстрирует факт приблизительного совпадения возраста рода и многих его видов (Шварц, 1980); это показывает, что значительная часть дивергенции происходит на самых ранних этапах эволюции возникшего таксона.
Закон архаического многообразия (Мамкаев, 1968) описывает резко повышенную изменчивость организации на ранних этапах эволюции таксона, отчего виды и роды в это время могут различаться по признакам, позже характеризующим семейства и отряды. Не менее характерна для этапа архаического многообразия и малая эволюционная устойчивость ранних членов таксона, отражающаяся в обилии коротких базальных ветвей едва ли не на любой филогенетической схеме, построенной с использованием богатого палеонтологического материала (рис. 4).
Рис. 4. Филогенетические древо скорпионниц (отряд Panorpida). По Новокшонову (Novokshonov, 2002), с изменениями
Все перечисленное позволяет сделать вывод, что переход группы в условия обедненного биоценоза и незаполненной экологической ниши действительно провоцирует эволюцию, причем эволюцию существенно инадаптивную. В таком случае становление эвадаптивных групп, т.е. гармоничных, приспособленных к заполненным биоценозам и напряженным конкурентным отношениям, можно связать с последующим ужесточением условий и интенсивным вымиранием большинства вновь возникших групп (тех, что оказались неспособными достаточно быстро превратиться из инадаптивных в эвадаптивные).
Проанализируем знаменитые опыты Г.Х. Шапошникова (1961, 1965, 1966, 1978). В этих опытах тли Dysaphis anthrisci maicopica Shap. пересаживались с пригодного кормового растения Anthriscus nemorosa MB. сначала на малопригодное (Chaerophyllum bulbosum L.), а затем на ранее совсем непригодное (Ch. maculatum Wild.). В результате тли за считанное число поколений не только изменились морфологически, но и приобрели репродуктивную несовместимость со своим видом и неполную совместимость с D. chaerophyllina Shap., исконным потребителем Ch. maculatum. В опытах Шапошникова отбор явно был односторонним и несбалансированным (инадаптирующим). Действительно, в опыте способность питаться на новом растении-хозяине на какое- то время оказалась единственным жизненно важным комплексом адаптации, а все остальные отошли на задний план. Конечно, опыт не был завершен, но если бы он был продолжен и перенесен в природу, то мы, вероятно, смогли бы наблюдать и следующий этап эволюции. А именно испытание зарождающейся группы всесторонним, эвадаптирующим отбором, для которого важны все аспекты адаптации — питание, размножение, индивидуальная устойчивость разных стадий онтогенеза в условиях различной плотности популяции, конкурентоспособность и т.д. Немного шансов, что конкретная популяция выдержит это испытание, но они есть.
Вернемся к проблеме мягких условий и ослабленного отбора. Можно ли считать условия опытов Шапошникова жесткими? На разных этапах эксперимента смертность личинок колебалась от 15-22 до 53-75%. Казалось бы, высокая смертность, но при указанном значении плодовитости (36,6 личинок от одной партеногенетической самки; данные только для конечного этапа опыта) она означает выживание как минимум около 10 личинок в потомстве каждой самки и, следовательно, почти десятикратный прирост популяции в каждом поколении. Другими словами, смертность в этих опытах была была весьма умеренной — очевидно, из-за того, что заботой экспериментатора враги тлей и другие неблагоприятные воздействия были по возможности устранены. Не удивительно, что линии в эксперименте оказались весьма устойчивыми и смогли быть поддержаны в течение 50 поколений — срок, который кажется не очень вероятным для отдельных линий в природных условиях. Что же до самого факта пересадки на малопригодное и совсем непригодное растение, то он как раз не является чем-то необычным. В природе тли должны часто оказываться в подобном положении: то крылатую самку занесло ветром в неподходящий биотоп, то дождь или ветер сбросили бескрылую тлю на другое растение. Просто в природе это трудно заметить, потому что там, в отличие от лаборатории, насекомое не на своем месте обычно гибнет.
Мы редко задумываемся, говоря о жестких, катастрофических и тому подобных условиях. Равновесная плотность популяции соответствует тем ее значениям, при которых смертность уравновешивает репродуктивный потенциал популяции. Это означает, что из всего потомства каждой самки в среднем только одной из ее дочерей удается оставить свое потомство («закон Бекетова»: А.Н. Бекетов, 1860; годом раньше о том же писал, например, Ч. Дарвин, 1991: 69). Поэтому элиминация в популяциях в норме (за исключением кратких и более или менее уравновешивающих друг друга эпизодов колебаний численности) оказывается практически равной плодовитости, т.е. близкой к максимуму.
Но если практически полная элиминация потомства есть норма существования популяции, если шансы на оставление потомства для каждой особи всегда минимальны, если вымирание целых популяций — обычное явление даже в экологическом (доэволюционном) масштабе времени, что же мы имеем в виду, говоря об ухудшении условий существования?
Похоже, что ужесточения условий на популяционном уровне в природе вообще не бывает. Даже в приложении к видовому и надвидовому уровню о нем говорить трудно. Здесь ухудшение условий может означать лишь тривиальное (в эволюционном масштабе времени) вымирание видов, влияние которого на эволюцию будет ощущаться лишь как освобождение экологического пространства и, соответственно, смягчение условий. Но природа не терпит пустоты, а экологическое пространство особенно, и выжившие виды мгновенно заполнят пустоту. Они интенсифицируют процесс размножения сохранившихся популяций и тем самым сведут к нулю кратковременное смягчение условий их существования.
Таким образом, эволюционно значимое смягчение условий существования столь же сомнительно, как и их ужесточение. Смягчение условий само по себе означает лишь повышение выживаемости и, следовательно, рост популяции. При этом мальтусова геометрическая прогрессия размножения за считанное число поколений приведет к насыщению экологического пространства и восстановлению нормальных для особи и популяции условий, включая нормальную (равновесную с плодовитостью, т.е. почти стопроцентную) смертность. Изменение характеристик популяции в результате колебаний ее плотности («волн жизни»), как известно, ограничено лишь изменениями в рамках нормальной внутривидовой изменчивости. Тот факт, что колебание плотности популяции представляет собой самое заурядное явление, и тем не менее случаев порожденного им необратимого эволюционного изменения до сих пор не описано, доказывает, что для эволюции это явление не существенно.
Итак, ни смягчение, ни ужесточение условий существования не представляют реальных факторов эволюции. Что же в таком случае может преодолеть устойчивость сбалансированного эпигенотипа и успешно вывести организацию живого существа за пределы исторически обусловленной нормы?
Причина устойчивости сбалансированного эпигенотипа заключается в однажды достигнутом компромиссе между противоречивыми потребностями оптимизации разных адаптивных функций организма. Эти потребности противоречивы в том смысле, что из-за сложного переплетения морфогенетических и функциональных связей между всеми частями и признаками организма адаптивное усиление одной функции должно отзываться потерями эффективности других. Ослабление целостности онтогенеза, облегчающее независимые преобразования отдельных систем, конечно, не только возможно, но и имеет место в природе. Однако это направление (упрощение организации и общая дегенерация) достаточно специфично, требует особых условий и не является ни характерным, ни особенно широко распространенным. В конце концов, чтобы терять высоту организации, надо сначала ее приобрести. Ослабление целостности как механизм эволюции существует, но для объяснения эволюционного процесса его явно недостаточно.
Таким образом, нам необходимо реконструировать условия, в которых изменение организации происходит в условиях сохраняющейся или растущей целостности, либо, скорее, когда временное снижение целостности быстро восполняется. Для этого попытаемся уточнить метафору адаптивного компромисса. Главную проблему для эволюции представляет, как мы видели, противоречивый характер отношений между адаптивными функциями в процессе их оптимизации. Противоречие создается, с одной стороны, тесными онтогенетическими и функциональными связями между системами обеспечения этих функций, а с другой — жестким контролем за должным исполнением всех этих функций. Мы видели, что ослабление связей возможно, но недостаточно для объяснения эволюции. Общее ослабление селективного контроля надлежащего исполнения адаптивных функций, как мы видели, тоже невозможно: в эволюционном масштабе времени элиминация всегда приближена к максимально возможным значениям. Где же выход?
Подсказкой здесь служат обсуждавшиеся ситуации, провоцирующие быструю эволюцию. Общее для них — попадание популяции в новые, непривычные условия. Наиболее прозрачные для нас опыты с тлями показывают, в чем именно состоит необычность новых условий. Там экспериментатор предъявил тлям очень жесткие требованию по одному или немногим параметрам (в данном случае — способность питаться на неподходящем растении), но при этом освободил их от жесткого средового контроля по другим параметрам: защитил от непогоды, хищников, паразитов, болезней и т.д.
Мне не известны исследования, анализирующие подобный расклад интенсивности селективного контроля у первопоселенцев на островах или в новых экологических нишах. Однако общие соображения относительно низкой напряженности конкурентных отношений в островных условиях по сравнению с материковыми весьма правдоподобны. Они подтверждаются относительно высоким иммиграционным потенциалом материковых видов на островах и крайне низким в обратной ситуации. В таких условиях ценотический контроль исполнения некоторых функций (например, функции защиты от хищников и специализированных паразитов, которые в островных сообществах нередко находятся в дефиците) может быть понижен в пользу усиления контроля по другим направлениям. В конце концов, на то условия и новые, чтобы расклад селективных требований там был иным, чем прежде, и в частности, чтобы некоторые функции контролировались существенно жестче, чем в прежних условиях. Если популяция при этом выжила, значит, суммарная элиминация устойчиво не превышала ее репродуктивный потенциал и, следовательно, селективный контроль других функций был снижен.
Таким образом, эволюцию провоцирует, по-видимому, только одностороннее смягчение средового контроля организации, при котором лишь некоторые из адаптивных функций остаются под жестким наблюдением отбора. Очевидным следствием такого одностороннего (несбалансированного) контроля является разбалансировка, дестабилизация сложившейся организации, и она действительно проявляется в подобных случаях (Жерихин в: Деятельность..., 1967; Беляев, 1974; Шишкин, 1984, 1987; МсСипе, 1990, и др.).
Поскольку при дестабилизации адаптивность системы неизбежно снижается, перестройка едва ли может происходить в нормальных условиях. Из этого следует, между прочим, что нарождающаяся группа вряд ли может внедриться в уже оккупированную нишу и вытеснить ее владельца. Более правдоподобно, что она либо займет нишу, освободившуюся при вымирании ее прежнего обитателя, либо сумеет создать (открыть) новую нишу. Может быть, именно поэтому нам известно довольно много правдоподобных примеров недавнего возникновения новых таксонов в островных условиях, а также при антропогенной интродукции. Особенно поражают гельминтологические примеры (цитированные в дискуссии по докладу Rausch, 1982: 184) о двух последовательных актах видообразования за 300 лет (у паразита домовой мыши, завезенной в Сев. Америку) и о дивергенции двух видов человеческих лентецов не более 16 тыс. лет назад. А вокруг нас таких примеров очень немного, хотя в обычных условиях и биоразнообразие, и степень его изученности гораздо выше, и, соответственно, больше вероятность обнаружения случаев естественного видообразования.
Сказанное позволяет предположить, что эволюция на видовом и в особенности на более высоких таксономических уровнях возможна лишь в результате такого изменения условий, при котором действующий на популяцию отбор становится существенно односторонним. Только такой отбор представляется способным преодолеть устойчивость прежнего хорошо сбалансированного адаптивного компромисса. Однако вновь возникшая группа, прошедшая дестабилизацию и потерявшая былую сбалансированность, должна пройти жесткий всесторонний контроль (отбор по всем адаптивным функциям) и под его действием восстановить свою устойчивость. Другими словами, эволюционное изменение должно пройти через два этапа — дестабилизацию и затем стабилизацию вновь сбалансированного эпигенотипа.
Как уже упоминалось, результаты этих событий можно отождествить соответственно с инадаптивным и эвадаптивным изменением организации. Другими словами, в процессе эволюции организация живых существ проходит три последовательных этапа: инадаптацию, эвадаптацию и стазис. Если, конечно, не случится вымирания, что возможно на любом из этапов, хотя наиболее рискованным в этом плане, естественно, является процесс эвадаптации, а наиболее спокойным — стазис. Первые два этапа как самые уязвимые по необходимости должны быть кратковременными, а стазис может продолжаться неопределенно долго. Выход из ста- зиса едва ли может быть спонтанным: причиной должно стать изменение условий. Если такое изменение вызывает рост общей элиминации за пределы репродуктивного потенциала, неизбежно вымирание популяции (и вида, если вымерли все его популяции, и старшего таксона, если то же случилось со всеми его видами). Если же условия существования меняются таким образом, что контроль организации становится односторонним, не исключено, что популяция сможет войти в процесс инадаптивного преобразования ее сбалансированного эпигенотипа. А вот сумеет ли она успешно пройти его, и тем более — пройти крайне рискованный процесс эвадаптации, это уже дело случая, причем случая весьма редкого.
Здесь нужно отметить еще одну важную черту нашей модели — ее авторегуляторность. Сколь высоко ни поднялась бы устойчивость (забуференность) эпигенотипа даже заметной части обитателей нашей планеты, катаклизмы вряд ли будут серьезно угрожать разнообразию жизни на Земле, если, конечно, иметь в виду эволюционно значимые отрезки времени. Даже если заметная часть биоразнообразия «закоснеет» в совершенстве и, стало быть, жесткости своих эпигенотипов и окажется существенно «прореженной» очередным катастрофой (субглобальным оледенением, астероидной либо ядерной зимой или подобным апокалипсисом), ничего страшного для биосферы не произойдет. Выжившие виды попадут в ситуацию неполных сообществ с существенно ослабленными конкурентными отношениями. Это будет провоцировать быструю инадаптивную эволюцию, и тем более быструю, чем более обширной была «прополка». По мере заполнения сообществ многие из вытащивших выигрышный билет его потеряют, но чем больше была доля неудачников, тем больше шансов у тех, кто сумеет продержаться чуть дольше. Свято место пусто не будет, оно заполнится. Правда, новые сообщества могут оказаться мало похожими на прежние.
Как известно, даже в окрестностях границы перми и триаса вымирание было хотя и масштабным, но не подавляющим: обычно говорят, что с середины верхней перми было потеряно порядка половины семейств и трех четвертей родов в морях и существенно меньше на суше (рис. 5). Тем не менее, в дальнейшем мир, оправившись приблизительно к середине триаса, оказался очень существенно иным, причем не только по составу, но и по тенденциям. Суммарное разнообразие, достигнутое в морях во второй половине ордовика, далее до середины верхней перми (двести миллионов лет, т.е. лишь немного меньше, чем мезозой и кайнозой вместе!) не росло и лишь колебалось вокруг позднеордовикского уровня. После же пермотриасового кризиса рост возобновился и продолжается до сих пор, уже многократно перекрыв позднепалеозойский уровень стабилизации (рис. 5). Очевидно, сама структура морских сообществ радикально изменилась, дав место гораздо большему росту биоразнообразия.
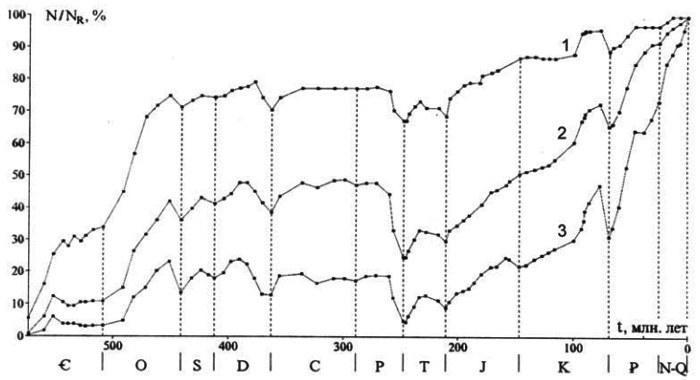
Рис. 5. Изменение разнообразия организмов моря [и суши] в фанерозое: отряды (1), семейства (2) и роды (3) морских животных (N/NR - отношение числа соответствующих таксонов на нижней границе века к числу современных таксонов, известных в рецентном состоянии)(по Алексееву и др., 2001)
Другая важная черта нашей модели эволюции — относительно малая предсказуемость результатов ее действия (весьма неполное соответствие между условиями протекания эволюции и ее результатами), что обусловлено большой сложностью эволюционирующей системы и, соответственно, невозможностью простых, предсказуемых решений. Что, конечно, является огорчительным свойством модели, но зато отражает реальность (в отличие от синтетической теории, из которой прогнозируемость эволюции следует, но не наблюдается).
Подход к адаптации как к компромиссу позволяет привести в соответствие с селекционистской парадигмой («эволюция есть процесс взаимодействия отбора с наличной организацией, т.е. с продуктом отбора предшествующих поколений») еще одну важную закономерность эволюции, не выводимую в общем виде из синтетической теории. Я имею в виду неравномерность эволюционного процесса, столь неопровержимо вытекающую из палеонтологических данных. Трехчленный цикл «инадаптация — эвадаптация — стазис» с очень быстро протекающими двумя первыми этапами и длительным периодом эволюционной стабильности легко находит параллели с моделями, предложенными палеонтологами (Симпсон, 1948; Eldredge, Gould, 1972; Stanley, 1975; Gould, Eldredge, 1977) и генетиками (Carson, 1975) с целью ввести неравномерность в синтетическую теорию. Это сходство несомненно, но его не следует переоценивать. Сходна лишь феноменология, а предполагаемые механизмы неравномерности в разных моделях различны, в том числе и в отношении их реалистичности.
Особняком стоит гипотеза ценотической регуляции эволюционного процесса (Красиков, 1969; Жерихин, Расницын, 1980; Жерихин, 1987; Е. Шварц, 2004). Эта гипотеза, также основанная на синтетической теории, на легкой податливости организации живых существ эволюционному давлению, вводит неравномерность в эволюцию, предполагая жесткость организации сообщества, которая и ограничивает свободу изменений организмов. В своем законченном виде, как она предстает у Е. Шварца (2004), логическая схема гипотезы ценотической регуляции эволюции может быть сформулирована следующим образом. Трофические цепи в нормальном сообществе напряжены (доступная энергия используется более или менее полностью) в результате эволюционного взаимного «притирания» организмов (коэволюции), так что «все места заняты» и эволюционировать некуда, кроме как медленно притираться дальше (когерентная эволюция по Красилову, 1969). Нарушения ценотической структуры провоцируют эволюцию, которая либо быстро затухает, либо, редко, переходит в лавинообразную некогерентную эволюцию, интерпретируемую как биоценотический кризис.
Описанная модель лучше соответствует наблюдаемым процессам, чем основанные только на дискретности биологического вида (см. выше), но она тоже далека от совершенства. Здесь не место разбирать гипотезу ценотической регуляции в деталях. Обращу внимание только на несоответствие в некоторых важных моментах. Если трофические сети повсеместно напряжены и «все места заняты», почему на суше (но не в воде) столь большая доля зеленой продукции устойчиво поступает прямо к редуцентам, минуя консументов? (знаменитый вопрос Ван Валена «Почему мир зелен?»; Van Valen, 1971). Если нарушения, тем более такие масштабные, как антропогенные, продолжаются тысячелетия, а скорость эволюции при снятых ценотических ограничениях должна приближаться к своим максимальным значениям (вид на считанное число поколений в опытах Шапошникова и за первые столетия на Гавайях, см. выше), то почему возникающий ценотический вакуум заполняется мигрантами из других стран или ценозов, а не вновь возникшими видами? И почему до сих пор нет грозных признаков предсказанной Жерихиным (Родендорф, Жерихин, 1974) катастрофы некогерентной эволюции? Все-таки эпигенетический подход предлагает более правдоподобную картину эволюционного процесса, чем синтетический — даже в биоценотической интерпретации.
Сказанное не означает, что сторонники синтетической теории никогда не обращались к идее устойчивости сбалансированного эпигенотипа. Именно об этом писали Э. Майр (1974; см. выше) и другие авторы (например, Van Valen, 1982; Laurin, Bruno, 1988). Проблема в том, что эта идея из совершенно другой, не синтетико-эволюционной парадигмы.
1.1.2.3. Проблемы
Таким образом, можно заключить, что многие наблюдаемые особенности эволюционного процесса находятся в неплохом соответствии с предсказаниями эпигенетической теории эволюции и, в частности, метафоры адаптивного компромисса. Однако остаются и проблемы. Загадка распределения эволюционных скоростей, столь очевидно противоположного предсказаниям синтетической теории, пока не поддается безупречному объяснению и с помощью эпигенетической теории, хотя и не находится в столь же неустранимом противоречии с ней.
Действительно, имеющиеся данные прямо противоречат концепции адаптивного компромисса, по крайней мере, на первый взгляд. Если главным фактором, тормозящим эволюцию, мы считаем компромиссную природу адаптации, в частности, трудность изменения хорошо сбалансированного эпигенотипа, то с усложнением эволюционирующей системы и, следовательно, с ужесточением компромисса следовало бы ожидать усиления этого торможения. На самом же деле, как мы видели, эволюция более сложных форм жизни протекает в целом быстрее.
Не все, однако, так просто. Прежде всего, повышенная трудность изменения высших организмов, предсказываемая концепцией адаптивного компромисса, означает меньшую частоту возникновения у них адаптивных изменений, но ничего не говорит о других факторах, определяющих скорость эволюции. Для высших организмов характерна направленность эволюции, своего рода ортогенетический фактор, повышающий кумулятивный эффект ряда последовательных изменений. Действительно, при ненаправленном, хаотическом характере внешних изменений, благоприятствующем броуновской эволюции, случайный возврат условий в состояние, близкое к ранее существовавшему, будет воспринят как таковой теми организмами, которые за прошедшее время изменились поверхностно, неглубоко. Более глубоко и целостно преобразованная система субъективно воспримет возврат к объективно прежней ситуации как новое изменение, требующее новых адаптаций. В результате эволюция высших форм жизни будет включать меньший броуновский компонент и, соответственно, больший объем накопленных изменений (Alberch, 1980; Rachootin, Thomson, 1981; Oster, Alberch, 1982).
Высшие организмы обладают также более ограниченными механизмами популяционной устойчивости. Конечно, индивидуальная устойчивость у них выше: в соответствии с «законом Бекетова» она тем выше, чем ниже характерный для группы темп воспроизводства (плодовитость и длительность генерации), а снижение этого темпа с повышением уровня организации вполне очевидно. Однако достигнутая на этом пути автономизация организма от среды, во-первых, ориентирована на эволюционно освоенную среду и не обязательно хороша в других условиях. Во-вторых, автономизация энергоемка, поскольку требует более тонкой и быстрой оценки состояния среды, без которой невозможны упреждающие реакции на неблагоприятные изменения. Как пишет С.А. Северцов, «горностай, которому для насыщения нужно в день добыть количество мяса, равное половине его веса, и гадюка, которая полгода проводит в оцепенении, а в летний период довольствуется двумя-тремя полевками в неделю, представляют хороший пример различия в энергии жизнедеятельности. Крокодилы в зоологическом саду довольствовались 36 г мяса в день. Близким по весу пантерам и леопардам дают 3,5-4 кг мяса в день» (1941: 288). Зависимость от среды здесь не столько снижается, сколько меняется, так что индивидуальная устойчивость совершенно не гарантирует устойчивости эволюционной.
Более важны в этом контексте механизмы популяционной устойчивости, прежде всего избыточная (резервная) плотность популяции, которая повышает смертность и (или) снижает плодовитость через факторы, зависимые от плотности популяции (Van Valen, 1976). Популяционная устойчивость универсальна, поскольку снижение избыточной плотности одинаково компенсирует снижение выживаемости независимо от причин этого снижения. Однако объем резервной плотности ограничен, так как ее нельзя устойчиво поднять за пределы емкости пространства ресурсов, а «срабатывание» избыточной плотности тоже имеет нижний предел, за которым выживание разреженной популяции становится проблематичным.
Существуют определенные основания полагать, что между объемом резервной плотности популяции и уровнем организации имеется отрицательная корреляция. Действительно, объем избыточной плотности в целом определяется положением порогов устойчивой плотности — нижнего, за которым популяция вымирает из-за слабости необходимого для выживания взаимодействия особей, и верхнего, превышение которого невозможно из-за ограниченности ресурсов. Верхний порог устойчивой плотности в конечном счете определяется величиной доступного популяции потока энергии, который она может использовать в обеспечение ее существования. При постоянстве этого потока верхний порог устойчивой плотности зависит от энергетических потребностей особи (я говорю об особях, а не о биомассе, поскольку обсуждаемые ниже факторы, влияющие на нижний порог устойчивой плотности, зависят именно от плотности особей). Энергетические же потребности особи быстро растут с повышением организации, во-первых, из-за в среднем более крупных размеров высших организмов, во-вторых, из-за дополнительных затрат на автоно- мизацию, о чем уже говорилось. Поэтому с повышением уровня организации идет быстрый рост энергозатрат особи и, соответственно, снижение верхнего порога устойчивой плотности популяции.
Нижний порог устойчивой плотности определяется различными факторами. Он возникает или повышается с появлением полового процесса, который для успешного размножения требует встречи двух особей, причем не любых двух. Сходный эффект дают адаптации кондиционирования среды (ее изменения деятельностью группы особей в направлении, благоприятном для группы). Первый фактор явственно, хотя и не жестко связан с высотой организации (партеногенез и бесполое размножение обычны и в умеренно продвинутых группах). Для второго механизма такая связь не очевидна, но допустима, поскольку высокая организация благоприятствует развитию более эффективных форм взаимодействия. Поэтому можно заключить, что с повышением организации верхний порог устойчивой плотности популяции сильно снижается, а нижний несколько повышается, снижая объем резервной плотности популяции и ее эволюционную устойчивость. Это не может не повышать скорость эволюции высших групп организмов.
Устойчивость популяции и вида в изменяющихся условиях в немалой степени зависит и от того, насколько организмы способны найти в новой среде мало изменившиеся островки и убежища и удержаться в них. Это, конечно, в большой мере определяется расселительными способностями организмов, которые вряд ли очень тесно связаны с уровнем организации. Но есть и другие, более перспективные факторы в данном контексте. Особую роль играют абсолютные размеры: там, где мелкие формы могут найти себе мало изменившиеся микробиотопы, крупные часто оказываются вынужденными измениться или вымереть. Так, возраст полусовременной фауны (фауны, составленной наполовину из вымерших таксонов, наполовину из доживших доныне) для крупных и мелких млекопитающих различается в 2,5 раза (200 тыс. лет для слонов и копытных против 500 тыс. лет для грызунов, насекомоядных и летучих мышей, Расницын, 1987, табл. 1), причем эти даты приходятся на крайне изменчивый по условиям плейстоцен. У насекомых полусовременная фауна по тем же данным датируется приблизительно 5 миллионами лет.
Как известно, большая часть крупных млекопитающих из своеобразной фауны вымершего тундростепного ландшафта, занимавшего в четвертичное время обширные пространства севера Евразии, вымерла в начале голоцена, а мелкие млекопитающие в основном выжили (Шер, 1997а,б; Жегалло и др., 2001). Насекомые же оказались устойчивыми в течение всего ледникового периода и послеледниковья: по данным В.И. Назарова (1984), из 2000 видов насекомых, известных для четвертичного периода, вымершими считаются не более 30. Из 731 вида, учтенного в базе данных для антропогена Сев.-Вост. Сибири (Sher еt а1., 2006), только 6-8 видов более или менее уверенно считаются вымершими. Более того, значительная часть характерных тундростепных видов насекомых позднего плейстоцена сохранилась не только в степях Южной Сибири и Монголии, но и в местах их прежнего обитания — на небольших реликтовых «степных» участках в Якутии и даже на Чукотке (Берман, 2001).
Размеры определенно, хотя и не строго, связаны с высотой организации: малые размеры ограничивают возможность усложнения, а крупные требуют какой-то минимальной сложности хотя бы для жизнеобеспечения и координации действий удаленных частей организма. Поэтому доступность стабильных островков в измененных условиях, пониженная для высших форм жизни, должна заметно снижать их эволюционную устойчивость и увеличивать скорость вымирания и эволюционных преобразований.
Перечисленные выше факторы вряд ли исчерпывают все разнообразие связей между высотой организации и устойчивостью, определяющих темпы эволюции, однако другие механизмы пока не столь очевидны. Так что в итоге нам удалось обнаружить один механизм, который, если концепция адаптивного компромисса верна, должен снижать темпы эволюции высших форм жизни. Это компромиссная природа адаптаций как таковая, т.е. трудность изменения более сложной сбалансированной системы. В противоположном направлении действуют более разнообразные силы. В их числе более выраженная направленность эволюции высших организмов, их пониженная эволюционная устойчивость из-за сближения порогов устойчивой плотности их популяций, и меньшая доступность для них «островков стабильности», сохраняющихся при общем изменении обстановки.
* * *
Завершая раздел по онтологии эволюционного процесса, можно сделать вывод, что эпигенетическая теория эволюции находится в хорошем согласии не только с результатами наблюдений и экспериментов в области осуществления и преобразования онтогенеза, но и с палеонтологическими данными по эволюции организмов.
Уверенной интерпретации оказался доступен довольно большой массив фактических данных в обеих областях, и это вселяет надежду, что и оставшиеся проблемы и вопросы со временем будут успешно разрешены.
|
|
А.П.Расницын. Теоретические основы эволюционной биологии // В.В.Жерихин, А.Г.Пономаренко, А.П.Расницын. Введение в палеоэнтомологию. М.: КМК. 2008. 371 с
1.1. ПРОЦЕСС ЭВОЛЮЦИИ
1.1.1. СИНТЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
1.1.2. ЭПИГЕНЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
1.1.2.1. Основные положения
1.1.2.2. Адаптивный компромисс
1.1.2.3. Проблемы
|