Українська мова (найновіша версія) / Русский язык (обновление прекращено)
4.11. Хищничество
Важнейшей характеристикой хищников является то, насколько они специализированы на питании определенными видами жертв. Например, обыкновенная лисица — чрезвычайно пластичный хищник, жертвами которого могут быть самые разные мелкие и даже средние по размеру млекопитающие, птицы, пресмыкающиеся, амфибии, самые разнообразные беспозвоночные и немало различных растений. В зависимости от того, какие виды пищи есть в ее местообитании, лисица может питаться и черепахами, и виноградом.
На противоположном полюсе высокоспециализированных хищников находятся, например, морские змеи из рода Микроцефалюс (буквально «мелкоголовые»). Голова и передняя часть туловища этой средней по размерам змеи очень узкие, «откалиброванные» по размерам норки донных морских угрей, которыми питаются эти змеи. Другим примером высокой специализации могут быть коалы, древесные представители австралийских сумчатых. Они питаются только молодыми листьями эвкалиптов.
Итак, по степени специализированности хищников на питании определенными категориями жертв их можно разделить на генералистов, как лисица, и специалистов, как микроцефалюсы. К этой классификации близко разделение хищников на монофагов (приспособленных к питанию одним видом жертв), олигофагов (ориентированных на несколько близких категорий жертв) и полифагов (способных питаться весьма разнообразными жертвами). Приведенные классификации не вполне идентичны, хотя, конечно, специализированные хищники обычно являются олиго- и монофагами, а генералисты — полифагами. Впрочем, бывает так, что какое-то достаточно узкое приспособление помогает освоить питание группой похожих по своих особенностям жертв. Например, гигантский муравьед высокоспециализирован на питании муравьями и термитами: его когти позволяют проламывать стенки термитников или добираться до муравьев, а длинный липкий язык, высовывающийся из вытянутой в трубку беззубой морды, дает возможность подхватывать и проглатывать одновременно сотни насекомых. Ничем другим, кроме муравьев и термитов, муравьед питаться не способен, но зато практически любой из достаточно многочисленных видов этих социальных насекомых может стать жертвой этого хищника-олигофага.
Специализированный хищник может быть более эффективным, чем неспециализированный, и это позволяет частично избегать конкуренции. С другой стороны, полифаги могут иметь более сбалансированную диету, легче находить корм, иметь более устойчивую популяционную численность.
Хищничество — один из важнейших механизмов передачи энергии от одних популяций к другим и является поэтому важным механизмом, делающим экосистему единым целым. Приведем некоторые примеры воздействия хищников на видовое разнообразие их жертв.
На литорали (прибрежных участках морского дна) в Новой Англии (США) чередуются участки, покрытые жесткими красными водорослями, относящимися к роду хондрус (Chondrus), а также участки, где преобладает мягкая зеленая водоросль энтероморфа (Enteromorpha). Исследователи решили выяснить, какие факторы определяют характер водорослей на том или ином участке берега — будет он поросшим хондрусом или энтероморфой. Выяснилось, что главным потребителем водорослей является брюхоногий моллюск литторина (Littorina). Литторина предпочитает энтероморфу и избегает хондрусов. Если взрослых литторин пересадить на участки, поросшие энтероморфой, они выедают зеленые водоросли и освободившееся пространство заселяется красными водорослями. Если на участке с красными водорослями убрать литторин, то поверхность хондрусов обрастает энтероморфами, которые со временем захватывают такие территории. Итак, хондрус растет там, где есть литторины, а энтероморфа — там, где их нет. Но почему моллюски есть в одних местах и отсутствуют в других?
Личинка литторин ведет планктонный образ жизни. Плавающие личинка оседают на берегу повсюду — и в зарослях хондруса, и в зарослях энтероморфы. В зарослях хондруса такие личинки нормально развиваются, а на участках с энтероморфой их быстро выедают крабы карцинусы (Carcinus).
Итак, красные водоросли растут там, где нет карцинусов, а зеленые — там, где они есть. Что же является причиной этого распределения? Почему крабы не обитают всюду?
Распространение крабов ограничивается выедающими их чайками. Чайки летают над всем побережьем, а крабы скрываются от них под густым покровом энтероморфы. Итак, там, где растет энтероморфа, крабы могут спрятаться от чаек и уничтожать моллюсков, которые привели бы к гибели энтероморфы… Видовое разнообразие прибрежных вод поддерживается на высоком уровне межвидовыми связями, основанными на хищничестве!
Вообще ситуация, при которой хищник препятствует снижению видового разнообразия своих жертв, достаточно типична. В степных заповедниках на территории бывшего СССР были вынуждены использовать умеренный выпас скота. Когда-то на степных территориях паслись стада копытных — диких лошадей тарпанов, диких ослов и южнее — даже джейранов и сайгаков. Поедая быстрорастущие растения, эти копытные не давали им вытеснить те виды, которые росли медленно. После того, как люди истребили стада диких копытных, они вывели в степи стада, табуны и отары домашних. А после того, как эти степи становились заповедными, вслед за прекращением выпаса скота в них начинали исчезать редкие виды трав. Исправить ситуацию помогает умеренный выпас домашнего скота — например, лошадей.
После того, как в 1964 г. на Британские острова был завезен миксоматоз кроликов (вызываемое простейшими заболевание), численность этих растительноядных животных резко снизилась. Результатом исчезновения кроликов стало снижение видового разнообразия луговой растительности. Кролики неспециализированы почти как сенокосилки, и выедают все луговые растения подряд. Тем самым они препятствует вытеснению более конкурентоспособными растениями более слабых и редких.
Обычно хищники-полифаги наиболее интенсивно используют самых многочисленных из своих потенциальных жертв, выравнивая при этом их относительную численность.
Вам приходилось когда-нибудь долго собирать грибы или ягоды или ловить рыбу на удочку? Вспомните, что вы видели после этого, закрыв глаза. Листву и ягоды, приподнятые грибами хвоинки или качающийся на волнах поплавок — в зависимости от того, на что было нацелено ваше внимание. В зависимости от той деятельности, которой вы занимались, в вашей психике активизировалась определенная поисковая модель. В результате ее активизации вы быстрее и лучше замечали неприметную ягоду, гриб или поклевку рыбы. С другой стороны, если вы были сосредоточены на поиске ягод, вы могли «пропустить» гриб, к обнаружению которого вы не готовились. В данном случае вы имели дело с механизмом переключения, которое происходит как путем формирования ключевого образа, выделяющегося из фона в первую очередь, так и изменением поискового поведения.
Как ни отличается психика человека от психики других животных, переключение на тот или иной вид жертв играет важную роль в активности всех хищников-полифагов. Например, в экспериментах гуппи (аквариумных рыб) кормили дрозофилами (мухами, плавающими на поверхности) и трубочником (малощетинковыми червями, зарывающимися на дне). Рыбы поедали непропорционально большее количество преобладающего вида жертв, так как искали свою добычу или у дна, или у поверхности — там, где было больше шансов найти корм. Мы не можем заключить, менялся ли образ искомой добычи в психике гуппи, но отчетливо регистрируем изменение ее пищедобывательного поведения.
Важная характеристика хищника-полифага, способного переключаться с одной жертвы на другую, — его реакция на изменение относительной частоты жертв (рис. 4.11.1). Хищник с реакцией I типа избирательно потребляет определенный вид жертв, даже если они редки, и повышает их потребление с ростом их распространенности. Хищник с реакцией II типа потребляет жертв определенного вида пропорционально их численности, пока они редки, а после какого-то уровня стабилизирует их эксплуатацию (насыщается). Самой распространенной, однако, является реакция III типа (которую и демонстрировали в эксперименте гуппи): редкая жертва потребляется непропорционально мало, и хищник обращает на нее «внимание» только тогда, когда ее численность достигает определенного предела. Распространенность третьего типа реакций хищников-полифагов приводит к тому, что хищники выравнивают численность своих жертв, сильнее эксплуатируя более распространенные виды и давая «передышку» редким.
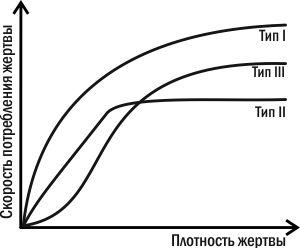
Рис. 4.11.1. Типы реакции хищника-полифага на изменение количества одного из видов его жертв
Многие особенности строения и образа жизни организмов объясняются их приспособлениями к защите от хищников или добыванию жертв. «Гонка вооружений» между хищниками и жертвами является одной из важных причин эволюции.
Например, растения, которые подверглись объеданию животными, производят новые листья с большим количеством защитных веществ: фенолов, восков, танинов. При разжевывании гусеницей дубовых листьев содержащиеся в отдельных компартментах клеток танины (дубильные вещества) высвобождаются, связывают белки и уменьшают их пищевую ценность. С другой стороны, если деревья и кустарники защитить от объедания растительноядными организмами, часть листьев может оказаться в условиях избыточного затенения другими листьями. В таких условиях дыхание в листьях преобладает над фотосинтезом: эти листья уменьшают, а не увеличивают продуктивность растения в целом. Удаление части листовой массы хищниками (растительноядными животными) лишь увеличивает продуктивность растения!
С другой стороны, если продуктивность растений будет оставаться постоянной, растительноядные животные могут достигнуть такой численности, что они смогут потребить весь урожай. Для многих растений (например, кедровой сосны) зарегистрированы периодические колебания урожайности. В неурожайный год семян немного, и такой год ограничивает численность млекопитающих и птиц, питающихся семенами. Зато в урожайный год семян будет столь много, что потребители просто не успеют съесть всю продукцию. За такой выигрыш растениям приходится серьезно «платить»: прирост колец у деревьев в урожайный год падает так же, как и при полном объедании листьев гусеницами. Синхронизация урожайных и неурожайных лет между разными деревьями обеспечивается как климатически, так и за счет химического взаимодействия растений.
Приспособления растений и растительноядных животных друг к другу могут быть самыми тесными. Вот один из многих примеров. На острове Маврикий встречается дерево кальвария (Calvaria major). Популяция этого растения находилась на грани уничтожения, так как в ней совсем не было молодых деревьев. Заинтересовавшиеся этим явлением биологи поняли, что возобновление кальварий прекратилось около 300 лет назад — тогда, когда на Маврикии были уничтожены дронты (Raphus cucullatus), — крупные нелетающие голубеобразные птицы. Было высказано предположение, что для нормального развития семян кальварии они должны были проходить через кишечный тракт дронтов. Для проверки этого предположения несколько семян кальварии скормили индейкам. Хотя несколько семян оказались раздавленными в желудках индеек, остальные прошли через кишечный тракт этих птиц неповрежденными и обрели всхожесть! Счастливая догадка помогла наладить искусственное восстановление популяции кальварий.
Долговременные реакции популяции жертвы на воздействия хищников не всегда отрицательны. Хищники могут уничтожать наименее ценных особей в популяции (больных, не имеющих индивидуального участка и т.д.), или же избежавшие гибели особи могут демонстрировать компенсаторные реакции. Если хищник эксплуатирует популяцию, страдающую от избыточной плотненности, его воздействие может повышать чистое пополнение или чистую продукцию.
В типичном случае популяции хищника стабилизируют численность популяций своих жертв. Когда жертв становится мало, хищники-полифаги переключаются на другие виды добычи, а хищники-олигофаги снижают свою численность вслед за жертвами в результате недокорма. Поскольку в большинстве случаев хищники размножаются медленнее своих жертв, восстановление их численности происходит медленнее, чем восстановление жертв. Таким образом, в типичном случае хищники просто не могут уничтожить своих жертв без остатка!
А в каком случае хищники могут истребить всех своих жертв? Человеку это удавалось неоднократно (как например, с уже упоминавшимися в этом пункте дронтами). До сих пор среди специалистов не существует единого мнения о том, какую роль человек сыграл в вымирании мамонтов (Mammuthus primigenius). Вероятно, исчезновение этого замечательного северного вида хоботных было связано с двумя группами причин: и потеплением климата, и охотой древних людей.
Потепление климата привело к тому, что тот биом, который населяли мамонты, — так называемая тундростепь или мамонтова степь, — исчез, на севере преобразовавшись в тундру, а на юге — в настоящую степь. Между тундрой и степью вклинились тайга и широколиственные леса. Однако не исключено, что масштабность перестроек экосистем была связана не только с изменением климата, но и с уничтожением со стороны людей мамонтов и других крупных растительноядных животных, питание которых поддерживало существование мамонтовой степи. А потенциальная возможность уничтожить мамонтов у человека была.
Полностью уничтожить определенный вид своих жертв может тот хищник, численность которого мало зависит от численности этого вида жертв. Значит, этот хищник должен быть полифагом и не быть специализированным на данном виде жертв. Человек с его гибким пищевым поведением мог выживать и в условиях недостатка или отсутствия мамонтов. Опасности подвергаются те жертвы, которые даже в условиях низкой численности не теряют привлекательности для хищников и будут приоритетно и эффективно потребляться в пищу. Это свойство было характерно для пары видов «человек-мамонт»: даже когда мамонтов было мало, они могли оставаться для древних людей «главной» добычей, которая эффективно отслеживалась и добывалась в первую очередь. Сложное поведение давало людям возможность успешно охотиться на мамонтов даже в условиях их низкой численности. Наконец, скорость роста популяции людей превышала скорость роста популяции мамонтов.
Итак, не зная точно, сыграл человек в вымирании мамонтов роковую роль или нет, мы можем предположить, что он потенциально мог ее сыграть и наверняка усугубил кризис численности этих животных, наступивший в результате потепления климата.
«Кто непосредственно угрожает существованию вида — это не «пожиратель», а конкурент; именно он и только он. Когда в давние времена в Австралии появились динго — поначалу домашние собаки, завезённые туда людьми и одичавшие там, — они не истребили ни одного вида из тех, что служили добычей, зато под корень извели крупных сумчатых хищников, которые охотились на тех же животных, что и они. Местные хищники, сумчатый волк и сумчатый дьявол, были значительно сильнее динго, но в охотничьем искусстве эти древние, сравнительно глупые и медлительные звери уступали «современным» млекопитающим.
Динго настолько уменьшили поголовье добычи, что охотничьи методы их конкурентов больше «не окупались», так что теперь они обитают лишь на Тасмании, куда динго не добрались» (К. Лоренц, 2001).
Фермеры, охотившиеся на тасманийских волков, смогли истребить этот удивительный вид хищных сумчатых. Истребление охотниками их жертв можно сравнить именно с отношением хищничества (хотя мотивация охотников на тасманийских сумчатых волков соответствовала скорее логике интерференционной конкуренции). До сих по Тасмании ходят легенды о том, что в каких-то труднодоступных участках острова какому-то счастливчику удалось встретить этого зверя, но, увы, скорее всего эти легенды отражают желания любителей дикой природы, а не суровую действительность.