Склад пуголовків зелених жаб (Pelophylax esculentus complex) в Іськовому ставі (НПП «Гомільшанські ліси»)
Макарян Р. М., Бірюк О. В., Коршунов О. В., Кравченко М. О., Мелешко О. В., Трохимчук Р. Р., Шабанов Д. А.
Харківський національний університет імені В.Н. Каразіна
d.a.shabanov@gmail.com
Іськів ставок, що розташований у Зміївському районі Харківської області в Національному природному парку «Гомільшанські ліси» (49º37´33´´ пн. ш.; 36º17´09´´ сх. д.) є водоймою з тривалою історією батрахологічних досліджень. Як зазначено в розміщеній у цьому ж збірнику роботі В.О. Стах зі співавторами, що присвячена Коряковому яру, дослідження зелених жаб на цьому ставку розпочато в 1995 р. Г.А. Ладою. Він описав тут «чисту» популяційну систему, що складалася майже виключно з диплоїдних Pelophylax esculentus. З 2000 р. Г.В. Шабанова та Д.А. Шабанов почали моніторинг складу популяції сірих ропух, що нереститься на цьому ставку. У 2000 р. Іськів ставок було спущено, а з 2001 р. — відновлено. Імовірно, саме це призвело до змін ГПС Pelophylax esculentus complex на цьому ставку; вона перетворилася на R-Em-HPS (склад Pelophylax esculentus complex і типологія його популяційних систем обговорюються у роботі В.О. Стах зі співавторами). За минулі з того часу роки ГПС зелених жаб Іськівого ставка зазнала кількох змін [6].
ГПС зелених жаб Іськівого ставка стала одною з перших ГПС у басейні Сіверського Дінця, що були досліджені сучасними методами. Унаслідок багаторічних досліджень було описано Сіверсько-Донецький центр різноманіття Pelophylax esculentus complex [3, 4]. Унікальність цього центру пов’язана з такими його особливостями:
— на частині його території поширені триплоїдні P. esculentus з обома можливими геномними композиціями (LLR і LRR);
— у ньому переважають R-E-HPS і R-E-Ep-HPS;
— у ньому немає статевозрілих P. lessonae; особини цього виду, які виникають у результаті гібридолізу (схрещування гібридів), гинуть на початкових етапах онтогенезу;
— у ньому реєструється значна кількість особин P. esculentus з гібридною амфіспермією (продукуванням статевих клітин обох батьківських видів);
— значна частина особин P. esculentus демонструє порушення фертильності.
Зміни ГПС Іськівого ставка та інших ГПС цього центру стали об’єктом імітаційного моделювання: спочатку засобами Microsoft Excel [1], а згодом — із застосуванням консольного додатку для Java, розробленого А.О. Леоновим [3, 8]. Моделювання проводили для ГПС, що складаються з диплоїдних жаб, які належать до форм, поширених у Сіверсько-Донецькому центрі різноманіття Pelophylax esculentus complex. Завдяки моделюванню встановлено, які ГПС зелених жаб є стійкими (зберігають у часі свою приналежність до певного типу). Такі ГПС можуть існувати лише в чотирьох стійких станах, у яких до їх складу входять представники P. esculentus. Два із цих станів належать до R-E-HPS, два — до E-HPS .
Для запису складу E-HPS і результатів схрещування, що в них відбувається, ми застосуємо таку систему. Геном P. lessonae позначається літерою L, P. ridibundus — R. Чоловічий геном позначається індексом Y, жіночий — X (у жаб така ж система спадкування статі, як у людей; гетерогаметною статтю є чоловіча). Клональність геному позначається взяттям його символу в дужки; у особин із гібридною амфіспермією обидва геноми клонально переходять у гамети і тому беруться у дужки. Можливі склади E-HPS є такими (символи особин батьківських видів, що виникають унаслідок схрещування гібридів та є нежиттєздатними, закреслені):
E-HPS type I:
♀(XL)(XR) × ♂(XL)(YR) → ♀♀(XL)(XL) : ♀♀(XL)(XR) : ♂♂(XL)(YR) : ♂♂(XR)(YR) → ♀♀(XL)(XR) : ♂♂(XL)(YR);
E-HPS type II:
♀(XL)(XR) × ♂(YL)(XR) → ♂♂(XL)(YL) : ♀♀(XL)(XR) : ♂♂(YL)(XR) :♀♀(XR)(XR) → ♀♀(XL)(XR) : ♂♂(YL)(XR).
В обох випадках сталий склад E-HPS характеризується кількома незвичайними особливостями:
— нерестове стадо в таких ГПС складається з представників двох клонів (чоловічого і жіночого), кожен із яких характеризується гібридною амфіспермією;
— у цих ГПС передається по три клональних геноми: геноми одного з батьківських видів обох статей і жіночий геном іншого батьківського виду;
— половина потомства в цих ГПС виникає внаслідок гібридолізу та гине до статевої зрілості;
— якщо гібридолізні представники хоча б одного батьківського виду доживають до віку, в якому можна визначити їх вид і стать, за нею можна розрізнити ці два типи ГПС.
Можна припустити, що більш імовірним є E-HPS type II. Гібриди, що клонально передають чоловічий геном P. lessonae, виникають частіше, ніж такі, що передають чоловічий геном P. ridibundus. Річ у тім, що самці P. lessonae під час нересту поводяться агресивніше й частіше утворюють пари з більш крупними самицями P. ridibundus.
Спостереження за ГПС Іськівого ставка [2, 6] доводять, що в ньому зменшується частка триплоїдів і знижується ефективність відтворення. Це може свідчити про повернення до одного з двох можливих типів E HPS.
Щоб визначити, до якого типу E-HPS наближається ГПС Іськівого ставка зараз (і, ймовірно, до якого типу вона належала у 1995 р., коли її досліджував Г. А. Лада), ми у серпні 2013 р. зібрали випадкову вибірку пуголовків і метаморфів із цього ставка. Ці пуголовки та жаби були дорощені до стану, коли їхню таксономічну приналежність можна було впевнено визначити за зовнішніми ознаками. Таке визначення дає надійний результат, коли стосується диплоїдів, і є недостатнім для точного визначення триплоїдів. Тому в усіх цих особин була визначена плоїдність із застосуванням непрямого (вимір розмірів еритроцитів) та прямого (каріоаналіз клітин кишечника) методів. Усі особини виявилися диплоїдними, що дає підстави вважати надійними результати визначення їх приналежності до P. ridibundus або P. esculentus. Стать молодих жаб визначали при розтині за морфологією гонад.
У тому разі, якщо б серед потомства було зареєстровано переважання самиць P. lessonae і самців P. ridibundus, можна було би зробити висновок, що ГПС Іськівого ставка наближається до E-HPS type I. E-HPS type II відповідає переважання ♂♂ P. lessonae і ♀♀ P. ridibundus. Зареєстрований варіант відповідає E-HPS type II, за винятком, що P. lessonae серед потомства не зареєстровано. Склад дослідженої вибірки був таким: 21 ♀♀ P. ridibundus (68%), 2 ♀♀ P. еsculentus (6%), 8 ♂♂ P. еsculentus (26%). Ми вважаємо, що відсутність P. lessonae серед потомства є наслідком того, що представники цього батьківського виду, які виникли внаслідок гібридолізу, гинуть на ранніх стадіях онтогенезу й не доживають до метаморфозу. Відсів P. ridibundus відбувається пізніше, і представників цього виду можна зареєструвати серед метаморфів і незрілих жаб. Така різниця у термінах загибелі представників батьківських видів, що утворюються внаслідок гібридолізу, може бути пов’язана з різним рівнем змін геномів батьківських видів, що викликані їх клональним спадкуванням. Є два механізми, здатні забезпечувати нежиттєздатність особин батьківських видів, які виникають внаслідок гібридолізу. По-перше, це — накопичення в них мутацій, очищення від яких негативним добором при клональному спадкуванні виявляється неефективним унаслідок популяційно-генетичного феномена, що має назву храповика Мюлера [7]. По-друге, — може бути наслідком позитивного добору клональних геномів на їх здатність забезпечувати елімінацію іншого, рекомбінантного геному [3]. Звісно, ці два механізми можуть працювати одночасно. У будь-якому разі геноми P. lessonae, статевозрілих представників яких немає в Сіверсько-Донецькому центрі різноманіття Pelophylax esculentus complex, мають бути змінені клональним спадкуванням більш глибоко, ніж геноми P. ridibundus. Таким чином, ми вважаємо, що в Іськівому ставку переважно реалізується схрещування, схему якого показано на рисунку.
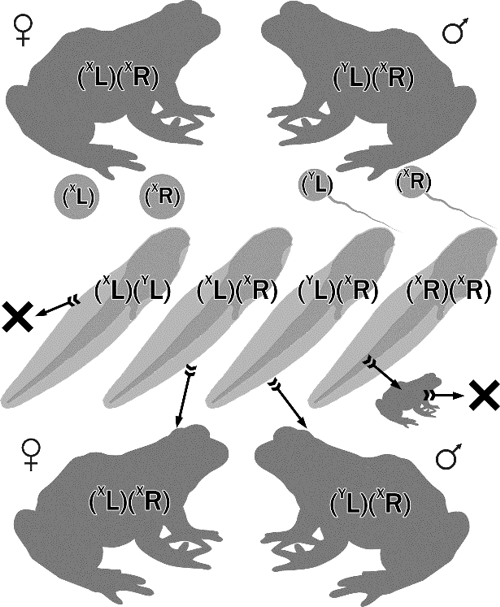
Схема схрещування двох амфіспермічних клонів, що характерне для E-HPS type II і переважає, за нашими даними, на Іськівому ставку
Як міг відбутися перехід від R-Em-HPS, що спостерігалася на ставку після його спуску та поновлення, до E-HPS type II? Для такого переходу достатньо було попадання у ставок однієї особини з гібридною амфіспермією. Ймовірний сценарій показано нижче. На етапі R-Em-HPS переважна кількість схрещувань була такою: ♀ XRXR × ♂ XR(YL) → ♂♂ XR(YL); ця ГПС могла існувати тільки завдяки потраплянню у ставок самиць P. ridibundus з інших місцеперебувань.
У разі попадання до ставка самиці з гібридною амфіспермією відбувалося таке схрещування: ♀(XL)(XR)×♂XR(YL)→♂♂(XL)(YL):♂♂(XR)(YL)→♂♂(XR)(YL). Схрещування ♂ (XR)(YL) з такого виводку зі самицями ♀ (XL)(XR) і є саме таким, що характерне для E-HPS type II. У разі виникнення такої ГПС вона виявляється певною мірою стійкою до попадання до неї інших геномів. Так, у разі, якщо деякі самиці P. ridibundus доживуть до статевої зрілості або потраплять у ставок з інших місцеперебувань, відбудуться такі схрещування: ♀ XRXR × ♂ (YL)(XR) → ♂♂ XR(YL): ♀♀ XRXR. Життєздатність самиць з такого потомства також, імовірно, має бути зниженою. Наявність певної частки ♂♂ XR(YL) може бути причиною переважання чисельності самців над самицями, що реєструється у ході спостережень за ставком.
Втім, якщо інші типи схрещування в такій ГПС будуть відбуватися систематично, інші форми жаб можуть витіснити два клони, що характерні для E-HPS type II. Річ у тім, що відтворення в обох типах E-HPS відбувається із нижчою ефективністю внаслідок того, що половина нащадків гине внаслідок гібридолізу.
Викладене дає підстави вважати, що подальші спостереження за ГПС зелених жаб Іськівого ставка становлять винятковий інтерес. Ми очікуємо, що ставок може вийти у стійкий E-HPS type II. На жаль, якщо відтворення жаб виявиться недостатньо ефективним, ця ГПС може загинути або, ймовірніше, зазнати витіснення формами жаб іншого складу.
1. Кравченко М.А., Шабанов Д.А. Моделирование трансформаций гемиклональных популяционных систем зеленых лягушек (Pelophylax esculentus complex; Amphibia, Ranidae) с помощью рекуррентных разностных уравнений // Вісник Харків. нац. ун-ту ім. В. Н. Каразіна. Сер.: Біологія. 2010. Вип.12 (№920). С. 70–82.
2. Черепашук И. В., Кочнева Е. П., Лаврикова С.С. и др. Исследование структуры популяционной системы зеленых лягушек (Pelophylax esculentus complex) Иськового пруда Змиевского района Харьковской области // Біологія: від молекули до біосфери: матеріали X Міжнар. конф. молодих учених (2–4 грудня 2015 р., м. Харків, Україна). Х.: ФОП Шаповалова Т. М., 2015. С. 185-186.
3. Шабанов Д.А. Еволюційна екологія популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) Лівобережного лісостепу України: автореф. дис. … д-ра біол. наук за спец. 03.00.16 — екологія // Дніпропетровськ, 2015. 36 с.
4. Шабанов Д.А. Коршунов О.В., Кравченко М.О. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми // Біологія та валеологія. 2009. Вип. 11. С. 116–125.
5. Biriuk O., Shabanov D., Korshunov O. et al. Gamete production patterns and mating systems in water frogs (hybridogenetic Pelophylax esculentus complex) in North-Western Ukraine // Journal of Zoological Systematics and Evolutionary Research, 2015. V. 54. I. 3. P. 215–225.
6. Meleshko O.V., Korshunov O.V., Shabanov D.A. The study of three hemiclonal population systems Pelophylax esculentus complex from the Seversko-Donetskiy center of green frogs diversity // Вісник Харків. нац. ун-ту ім. В. Н. Каразіна. Сер. Біологія. 2014. Вип.20, №1100. С. 153-158.
7. Plötner J. Die westpaläarktichen Wasserfrösche / Bielefeld: Laurenti-Verlag, 2005. 161 S.
8. Shabanov D., Vladymyrova M., Leonov A. et al. Simulation as a Tool to Identify Dynamical Typology of Water Frog Hemiсlonal Population Systems // Accepted to Acta Biotheoretica, 2016 (in press).