Подкласс Synaptosauria
Группа ящерицеобразных животных, существовавших с начала перми до мела.
Обладают явно выраженным верхним (парапсидным) окном, при этом функциональный аналог нижнего окна, если он имеется, формируется за счет изгиба верхней скуловой дуги, вследствие чего сохраняется контакт между скуловой и квадратно-скуловой костями черепа. Водные группы никогда не приобретают рыбообразной формы тела, а основным приспособлением локомоции являются крупные ластоногие конечности.
Существуют предположения, что примитивные группы синаптозаурий выступают в качестве возможных предков диапсидных рептилий. Так, Claudiosaurus (поздн. Пермь, Мадагаскар) считается предком завроптеригий, входящих в состав диапсид в широком смысле, т.е. является переходной формой от ранних диапсид-эозухий, к типичным завроптеригиям.
Подкласс состоит из примитивных наземных Araeoscelomorpha и полуводных и водных Sauropterygiomorpha.


Petrolacosaurus kansensis Lane, 1845 (Araeoscelomorpha) Pistosaurus grandaevus Meyer,1839 (Sauropterygiomorpha)
Состав надотрядов выглядит следующим образом:
Надотряд Araeoscelomorpha
Отряд Araeoscelidia
Отряд Trilophosauria
Надотряд Sauropterygiomorpha
Отряд Sauropterygia
Подотряд Eosauropterygia
Подотряд Plesiosauria
Отряд Placodontia
Подотряд Placodontoidea
Подотряд Cyamodontoidea
Характерной особенностью отряда Araeoscelidia является удлиненные шея и череп.
Петролакозаурус и Араеосцелис из ранней перми и карбона, являются одними из наиболее ранних диапсид. Это сходные ящерицеобразные формы, питающиеся насекомыми и имеющие хорошо развитую систему зубов.

Petrolacosaurus kansensis Lane, 1945
Как приспособление к более мощному укусу (для раскола твердых панцырей насекомых) у Араеосцелиса имеются только задние ряды зубов, но они крупнее, а также зарастает нижнее окно, для укрепления челюстей.

Araeoscelis gracilis (Williston 1910)
Более развитыми являются мелкие планирующие виды из семейства Weigeltisauridae у которых по бокам тела имеются специализированные ребра, наиболее вероятно, служащие для поддержания летательной перепонки. Сходное приспособление имеется у современных Летучих драконов (Draco L., 1758).


Передние и задние конечности одинаковой длины, фаланги длинные с короткими мощными когтями. В черепе также, как и у Араеосцелиса, имеется одно височное окно.
Такие формы характерны для всех родов семейства:

Weigeltisaurus Kuhn, 1939 - поздняя пермь Центральной Европы;


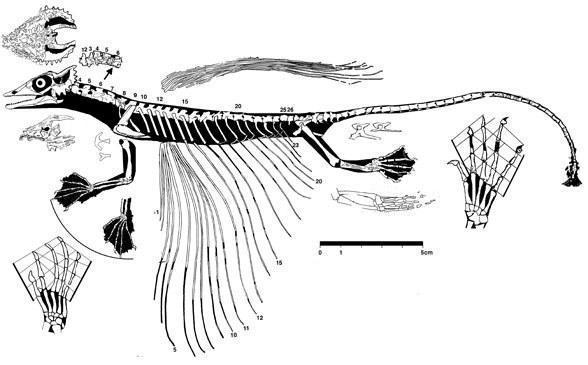
Coelurosauravus Piveteau, 1928 - поздняя пермь Мадагаскара;
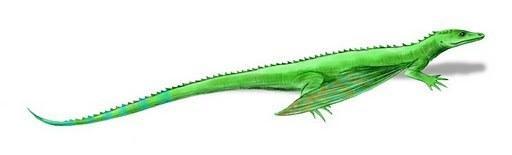
Rautiania Bulanov et Sennikov , 2006 - поздняя пермь Восточной Европы;
а также для Wapitisaurus Brinkmann, 1988 из раннего триаса Северной Америки.
Дополнительно к араеосцелидиям относят: нижнепермских представителей из рода Zarcazaurus (Brinkman et al., 1984), возможно вторично утративших нижнее височное окно; мелких длинношеих из родов Kadaliosaurus (Kuhn., 1969) и Aphelosaurus (Evans, 1988); одними из первых возвратившихся в воду рептилий - плавающих Spioequalis (deBraga & Reisz, 1995), о чем свидетельствуют анатомические преобразования: удлиненные остистые и гемальные отростки хвостовых позвонков.
Отряд Trilophosauria
Крупные (до 2,5 м в высоту) триасовые травоядные рептилии.
В состав отряда входит единственное семейство Trilophosauridae Gregory, 1945, для представителей которого характерно отсутсвие передних зубов и замещение их своеобразным роговым клювом. Имеется только один коракоид. Таз широкий, задние конечности длинее передних, стопа с пятью пальцами.
Выделяют 7 родов:
Trilophosaurus Case, 1928;
Teraterpeton Sues, 2003;
Anisodontosaurus Welles, 1947;
Variodens Robinson,1957;
Coelodontognathus Otchev, 1967;
Doniceps Otschev, 1968;
Vitalia Ivachnenko, 1973.
Представители из первых двух родов являются наиболее типичными:

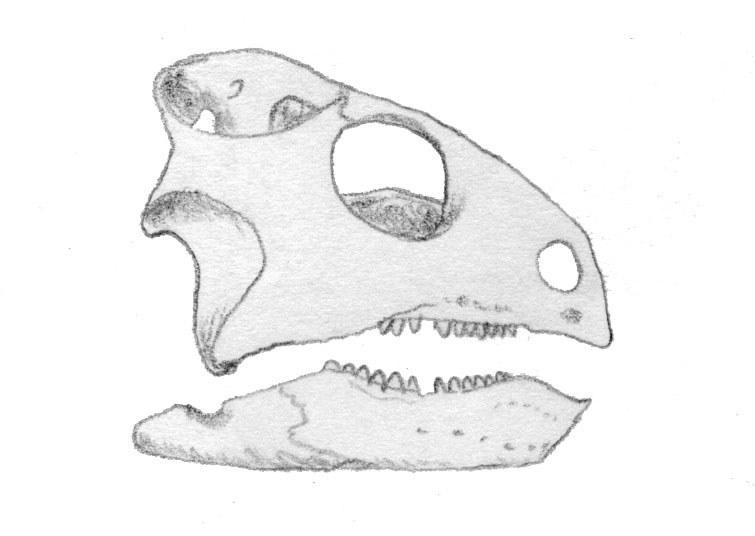
Trilophosaurus buettneri Case, 1928 - поздний триас Северной Америки
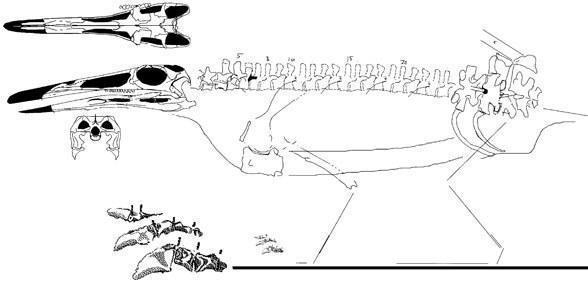
Teraterpeton hrynewichorum (Sues, 2003) - поздний триас Северной Америки
Надотряд Sauropterygiomorpha
.Отряд специализированных крупных водных и полуводных (морских) мезозойских рептилий, с массивным, коротким и уплощенным туловищем и удлиненной шеей. В черепе редуцированы надвисочные, табличатые и заднетеменные кости, закрытое небо. Позвонки платицельные или слабо амфицельные. Интерцентров нет. Развит один коракоид. Конечности преобразованы в ласты. Фаланги удлинены, характерна гиперфалангия.

Отряд Sauropterygia
Рыбоядные рептилии, обитающие в лагунах и прибрежной зоне морей, известные с нижнего триаса Западной Европы и Китая и просущестовавшие до терминального мела.
Среди наиболее уникальных апоморфий выделяют:
-Лопатки в плечевом поясе распологаются поверхностно по отношению к ключицам (у других тетрапод наоборот), нижние выросты которых образовывают полную дугу впереди коракоидов, которая способна соединяться с ними срединным выростом лопаток. Массивные коракоиды объединены протяженным симфизом (Carroll, Gaskill, 1991; Rieppel, 1987). Коплекс этих преобразований связан с утратой функции поддержания головы и передней части тела в пространстве при водном образе жизни.
-Между платицельными позвонками образуются сложные сочленения: зигосфены и зигантры, которые обеспечили завроптеригиям большую подвижность и гибкость тела.
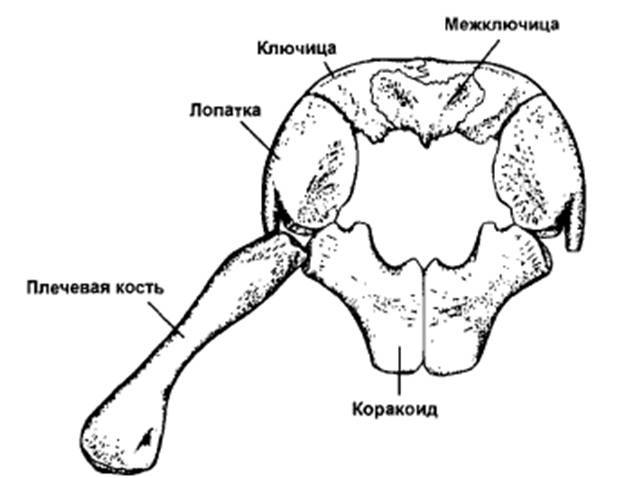
Плечевой пояс примитивной завроптеригии Pachypleurosaurus edwardsi (средн. триас Швейцарии, по Carrol, Gaskill, 1985).
Подотряды Sauropterygia - примитивные Eosauropterygia и более специализированные Plesiosauria хорошо различаются по степени их приспособленности к жизни в воде.
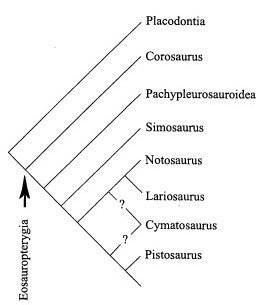
Филогения Sauropterygia по Rieppel, 1997
В составе Eosauropterygia рассматриваются слабо специализированных пистозавров, пахиплезиозавров с сильно выраженным пахистозом ребер и позвонков, а также высокоспециализированных нотозавров (Rieppel, 1994).
Пистозавры обладают удлиненным рылом, теменные кости очень узкие, лобные кости образуют задне-боковые отростки, выходящие в край височных окон. Также сохраняется межптеригоидная яма и хорошо развиты задневисочные окна. Зубы расположены редко, а передние нижнечелюстные преобразованы в клыки. Позвонки амфицельные, крестцовых позвонков три, крестцовые ребра на конце расширены.
Типичный представитель Pistosaurus longaevus (Huene, 1948)

Среди пахиплезиозавров наиболее известен мелкий среднетриасовый Keichousaurus hui Young, 1958, у которого отмечен значительный рост плечевой кости, по строению которой также выражен половой диморфизм. Такое преобразование плечевого пояса свидетельствует о более примитивном способе плавания - по типу гребли, а не "полета".


Keichousaurus hui Young, 1958
Нотозавры - более крупные ящеры, чем пахиплеврозавры. Обладают крупными зубами (могут развиваться клыковидные зубы). Позвонки платицельные, шея удлиненная. Заглазничный отдел черепа несколько удлинен, скуловая кость входит в переднее основание височной дуги, а теменные кости резко сужены, что приближает нотозавров к высшим завроптеригиям. Для надсемейства характерно наличие разных морфологических форм.
Самое обособленное положение имеет Simosaurus из среднего триаса Западной Европы, уникальность которому придают:
-Т-образная форма межключицы;
-морда сравнительно короткая и широкая, ее предносовое сужение не выражено;
-теменные кости сравнительно широкие, но срастаются с возрастом;
-зубы расположены тесно, клыковидных форм не наблюдается.

Simosaurus gaillardoti Meyer, 1842
Продвинутые Plesiosauria появляются в юре и достигают наибольшего разнообразия в юре и мелу Западной Европы и Северной Америки. Обычно обитатели морских водоемов, но встречались и в пресных водоемах. Длина более 3 м, в отдельных случаях 15-16 м. Укороченный хвост и благодаря развитию мощных брюшных ребер, занимающих все пространство между плечевым и тазовым поясами - малоподвижное туловище.
Такое необычное строение дало основание Д.Сили (Seeley, 1892) сравнивать этих морских ящеров "со змеей, продетой сквозь панцирь черепахи", как, например, это хорошо видно у Rhomaleosaurus cramptoni.

Rhomaleosaurus cramptoni (Tate and Blake, 1863)
Плезиозавры плавали при помощи обеих конечностей, не используя боковых изгибов тела, путем одновременных вертикальных взмахов, хотя задние конечности могли выступать и в качестве рулей, такой тип плавания называют "подводным полетом". Морфологически это ассоциировано с разрастанием и уплощением вентральных отделов поясов конечностей.
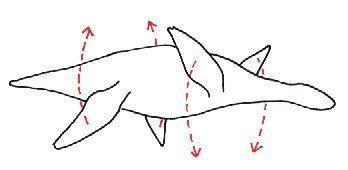
Тип плавания "подводный полет" у Plesiosauria.
В их состав входят надсемейства Pilosauroidea и Plesiosauroidea, обладающие противоположными морфологическими характеристиками, что свидетельствует о разных путях адаптации к одинаковым условиям.
| Pilosauroidea | Plesiosauroidea |
|
крупная удлиненная голова и короткая шея нижнечелюстной симфиз удлинен шейные позвонки укорочены шейные ребра двухголовчатые пердние конечности короче задних |
короткий узкий череп и удлиненная шея нижнечелюстной симфиз укорочен шейные позвонки удлиненные шейные ребра одноголовчатые передние конечности длиннее задних |
Для Pliosauroidea характерно наличие крупной головы (до 3 м) и сранительно короткой шеи (11-30 позвонков). Pilosaurus irgisensis (Novozhilov, 1948) дин из наиболее известных юрских представителей, обнаруженых на территории России.

Pilosaurus irgisensis (Novozhilov, 1948)
Наиболее длинношеими среди плезиозавров являются представители рода Elasmosaurus из верхнего кампана и нижнего маастрихта Северной Америки, встречавшиеся не только в морях, но и в персноводных озерах. По длине шея превышала голову в 14 раз, а число позвонков в ней достигало 76. В челюстях часть зубов превращена в клыки.
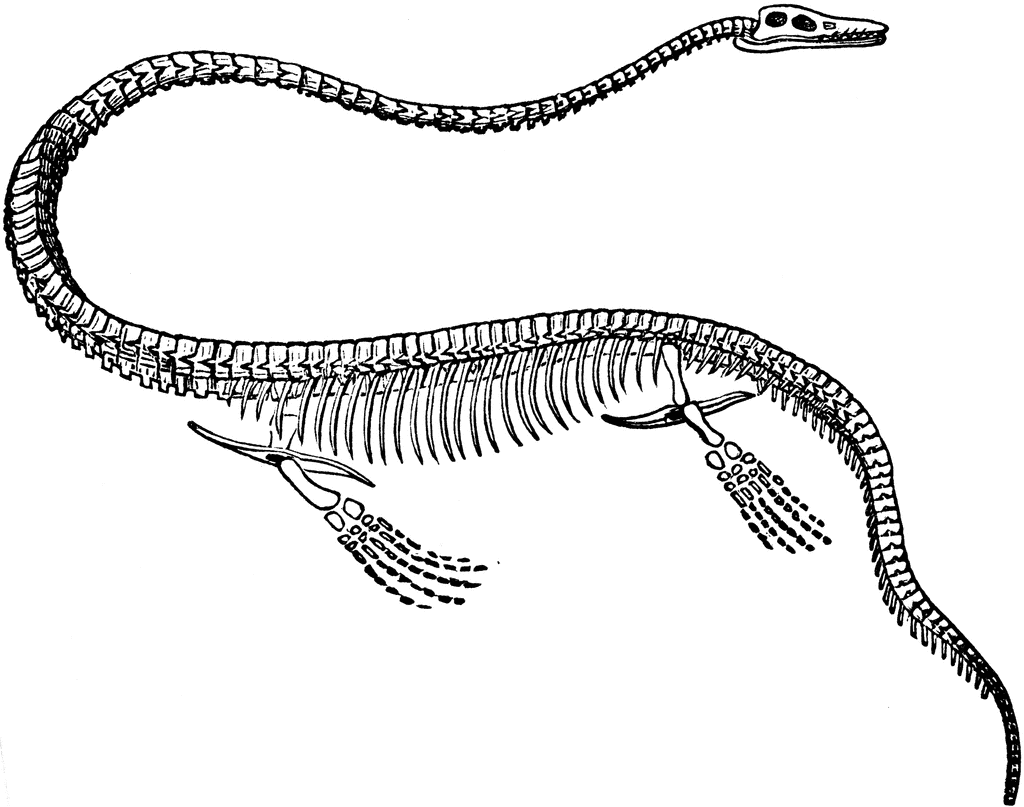
Elasmosaurus (Coup, 1868)
Считается, что именно плезиозавры стали прототипом для мифа о Лохнесском чудовище.

Отряд Placodontia
Обычно Placodontia относят к завроптеригиями в ранге отряда. От типичных завроптеригий они радикально отличаются в строении щеки - с необычно массивной квадратноскуловой костью, которая может входить в край височного окна.
Прибрежноморские триасовые ящеры, обитающие преимущественно в районе моря Тетис (предшественник Средиземного моря). Свое название получили по причине чрезвычайно уплощенных и крупных зубов, особенно больших на небе. Длина без хвоста до 1-1,5 м (иногда до 2,5 м). Череп массивный, акинетичный с упрочненной связью крыши черепа с небом. Как и завроптеригиям, плакодонтам свойственно одно височное окно, но область нижнего, где у завроптеригий обычно развита более или менее обширная вырезка, у плакодонтов, как и у примитивных ихтиозавров, закрыта крупными и массивными quadratojungale, перекрывающим всю щечную область. Морда короткая, но у некоторых панцирных вытянута в недлинный хоботок. Позвонки глубоко амфицельные, туловищные ребра одноголовчатые, массивная ключица, укороченная подвздошная кость, пальцы без признаков гиперфалангии, некоторые представители снабжены когтями, что указывает на сохраненную способность передвижения по суше.
Мало глубоких приспособлений к плаванию, а те, которые имеются выполняют эту функцию недостаточно эффективно.
Плакодонтов обычно разделяют на два подотряда: Placodontoidea (Дюгонеобразные) и Cyamodontoidea (Панцирные "черепахообразные").
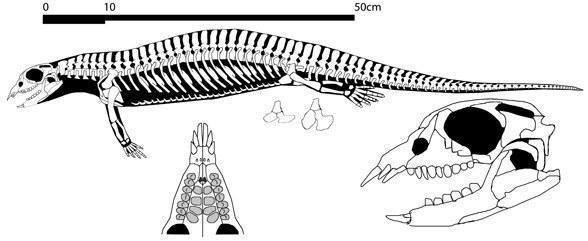
Беспанцирные Placodontoidea (более примитивные) обладают грузным телосложением с удлиненным хвостом (40-50 позвонков) и хорошо развитыми пятипалыми конечностями. Длинные, выступающие вперед массивыне резцы свидетельствуют о приспособлении к отрыванию от прикрепленных моллюсков и брахипод. Отряд представлен двумя семействами Paraplacodus (в максилле 7 зубов) и Placodus (до 5-ти).
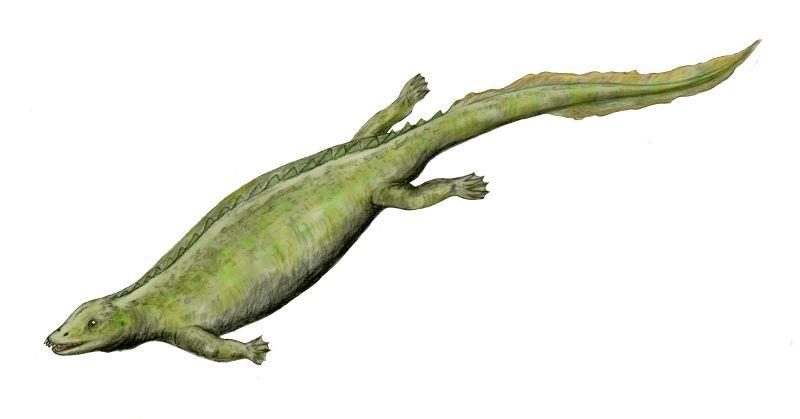
Paraplacodus broilli (Peyer, 1931)

Placodus gigas (Agasis, 1833)
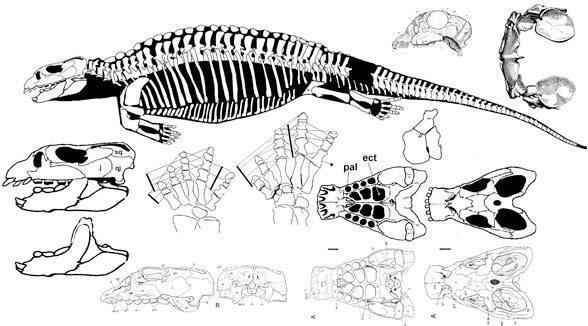
Панцирные Cyamodontoidea обладают большим разнообразием форм, чем плаконтоидеи. Тело резко уплощено и покрыто панцирем, образовавшимся из расширенных ребер и остеодермальных пластинок, который не прикрывает плечевой пояс, как у черепах. Тазовая часть спинного панциря обычно отделена от основной передней части, за исключением Placochelys с цельным панцирем. Панцирь покрыт роговой чешуей, которая образует продольные кили, по-видимому, увеличивающие гидродинамические качества этих плакодонтов. Рыло укорочено, резцы сохранены у отдельных представителей. Рыло образует подобие хоботка: наиболее удлинен у верхнетриасовых Plachochelys и Psephoderma, отсутствует у Henodus.


Psephoderma alpinum Meyer, 1858 Placochelys placodonta Jaekel, 1902

Henodus helyops (von Huene, 1936)
В составе подотряда выделяют около пяти семейств: Cyamodontidae, Macroplacidae, Protenodontosuridae, Placochelyidae, Henodontidae.
Большинство исследователей склоняются к тому, что наиболее примитивными являются Cyamodontidae c уменьшенным до двух максиллярных верхнечелюстным рядом зубов, у китайского Sinocyamodus спинной панцирь не прикрывает поясов конечностей. У всех остальных, исключая Protenodontosuridae, резцы исчезают. Macroplacidae и Protenodontosuridae считают близкими к Placochelyidae и удаленными от Cyamodontidae. У плахохелид очень узкие небные отростки, а у Macroplata расширенные задние небные зубы. У Henodus также нет зубов и не образуется хоботок, а сильно поперечно расшииена морда.
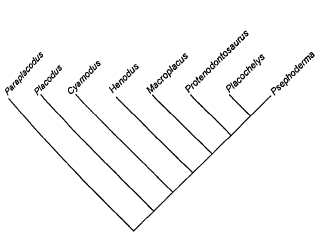
Кладограмма филогении плакодонтов (по [Rieppel, 2000b])
Плакодонты несмотря на относительно небольшое разнообразие, ярко иллюстрируют обилие параллелизмов в эволюции их черепа и зубов.