Маро А. Н., Шабанов Д. А. Механизмы расселения серой жабы (Bufo bufo (L. 1758); Amphibia, Anura) и особенности ее популяций в недавно заселенных местообитаниях // Матеріали Першої конференції Українського Герпетологічного Товариства. – К.: Зоомузей ННПМ НАНУ, 2005. – С. 107–110.
Механизмы расселения серой жабы (Bufo bufo (L., 1758); Amphibia, Anura) и особенности ее популяций в недавно заселенных местообитаниях
А. Н. Маро, Д. А. Шабанов
Харьковский национальный университет им. В. Н. Каразина, пл. Свободы, 4, Харьков 61077
This work is devoted to the studies of common toad Bufo bufo (Linnaeus, 1758) artificial moving to new habitats consequences that was performed in Kharkov region in 1994. The natural dispersion of toads has been found to occur in two ways. The first way includes a transfer of tadpoles with water. The second way includes incoming of toads into the reservoirs located between a former place of spawning and a ground habitat of toads settled by toads upon their metamorphosis. The new-founded groups of breeding (local populations) differ from their progenitor groups with the substantially smaller size of males and bigger size of females. It seems that on settling of new habitats, common toad males tend to accelerate their maturing, while females tend to accelerate their growth.
Для жаб рода Bufo характерно образование хорошо изолированных групп размножения (локальных популяций), связанных с нерестовыми водоемами и поддерживаемых благодаря филопатрии. Каждая группа размножения обладает уникальностью, обусловленной ее историей, особенностями населяемых местообитаний и взаимодействием с соседними группами. Изучение формирования новых групп размножения при расселении жаб может быть очень полезно для понимания их особенностей. В 1994 г. в Харьковской обл. при участии одного из авторов данной работы было выполнено искусственное расселение серых жаб (Bufo bufo (Linnaeus, 1758)). В ходе изучения его последствий нами изучены выборки серых жаб из 8 перечисленных ниже местообитаний (всего 244 половозрелых особи, пойманы на нересте, из которых 84 самки и 160 самцов) и проведены полевые наблюдения за ходом их нереста.
1. Старица: берег р. Северский Донец в окр. биостанции ХНУ им. В. Н. Каразина в с. Гайдары Змиевского р-на; обмелевшее старое русло реки во влажной дубраве с зарослями черемши; 9 самок, 9 самцов.
2. Коряков яр: окр. биостанции ХНУ; заиленный пруд в дубраве; 22 самки, 27 самцов.
3. Иськов пруд: окр. биостанции ХНУ; балочный пруд между дубравой и лугом; 4 самки, 13 самцов.
4. Нижний пруд, Пятихатки: окр. пос. Лесное Харьковского р-на; нижний из трех прудов в Очешетянской балке в дубраве; 15 самок, 19 самцов.
5. Верхний пруд, Пятихатки: там же, верхний из трех прудов; 8 самок, 16 самцов.
6. Верхний пруд, Ольховая балка: окр. пос. Русская Лозовая Харьковского р-на, верхний из 7 прудов в широкой балке, проходящей через нагорную дубраву; 9 самок, 26 самцов.
7. Нижний пруд, Ольховая балка: 6-й пруд у выхода балки в пойму р. Харьков; 7 самок, 22 самца.
8. Добрая балка; пруд в балке, выходящей в пойму рядом с Ольховой балкой; 10 самок, 28 самцов.
Экологически местообитания 2—8 очень сходны и типичны для серой жабы в Харьковской обл.; местообитание 1 существенно отличается от них. Географически изученные точки образуют три группы, расстояние внутри которых не превышает 5 км: 1—3; 4 и 5; 6—8. Группы размножения в точках 1—3 являются старыми, а 4—8 — молодыми; моложе всего локальные популяции в точках 5 и 8. Генеалогическая связь изученных популяций: 3->4->5; 3—>6->7->8. Расселения 3->-4 и 3->6 произведены искусственно в 1994 г. в результате перемещения не отметавших икру жаб, пойманных на нересте в Иськовом пруду. В Пятихатки перенесено около 200 особей обоих полов, в Ольховую балку — около 100 особей; при этом была изъята лишь малая часть процветающей группы размножения Иськова пруда. Для вселения жаб были выбраны водоемы, экологически похожие на Иськов пруд, но расположенные в хорошо изученном харьковскими герпетологами лесном массиве, где отсутствовали серые жабы. Дальнейшее распространение жаб проходило естественно, причем в случае 4->5 против тока воды в системе прудов, в случае 6->7 — по току воды, а в случае 7->8 — через водораздел. В 1995 и 1996 гг. во время нереста серых жаб найти их в нижнем пруду в Пятихатках не удалось. Вероятно, вселенные жабы отнерестились в новом для них водоеме, но вторично в него не вернулись. Развитие группы размножения оказалось связано с возвращением потомства особей-основателей в водоем, где происходило их развитие.
В Ольховой балке, куда жабы были перенесены в верхний пруд, они быстро распространились по всей системе, связанной общим водотоком. Очевидно, их расселение было связано с переносом головастиков водой. В Пятихатках средний (расположенный между нижним и верхним) пруд заселялся существенно позже нижнего, а позже всего оказался заселен верхний пруд (5-е местообитание). В 2004 г. на верхнем пруду в Пятихатках зарегистрировано лишь несколько половозрелых жаб, пришедших на нерест, а в 2005 г. их количество увеличилось более чем на порядок. Очевидно, что в данном случае был использован иной механизм расселения, направленный против водотока. Для его понимания существенны наблюдения, сделанные в 2005 г. на среднем пруду.
Мы наблюдали, что большинство находящихся в амплексусе пар жаб на среднем пруду двигалось в направлении нижнего пруда. Самки подходили к пруду, неся на себе самцов, или встречали их в воде. Затем пары двигались в направлении удерживавшей пруд плотины, выходили на берег и скатывались в нижний пруд. Лишь некоторые пары в среднем пруду оставались в нем и далее не перемещались. Вероятно, при расселении сеголеток из нижнего пруда некоторые из них мигрировали за средний пруд. По достижении половозрелости, двигаясь к месту своего развития, они спускались в балку и попадали в расположенный на их пути средний пруд. Большинство жаб преодолевало его и доходило до конечной точки своего маршрута, но некоторые не успевали туда дойти и нерестились в среднем пруду. Их потомство возвращалось уже в средний пруд и со временем колонизировало верхний.
Итак, можно предположить, что освоение новых мест нереста жабами происходит двумя способами: при переносе головастиков током воды и при попадании в водоемы, расположенные между ранее освоенным местом нереста и наземным местообитанием расселившихся особей.
Сравнение выборок проведено по 24 морфометрическим, 21 качественному и 8 дискретным признакам жаб, а также по 29 пропорциям (соотношениям морфометрических признаков). Компонентный анализ проводился по нормализированным средним значениям в выборках и выполнялся с помощью пакета Statistica for Windows (рис. 1). Видно, что самки оказались менее изменчивыми, чем самцы, и взаимное расположение их выборок на плоскости главных компонент более соответствует генеалогическим отношениям между группами размножения. Отличия выборок из окр. биостанции ХНУ (1-3) превышают отличия выборок из молодых групп размножения, которые в большинстве случаев оказываются ближе всего к выборке из родительского местообитания (3). Самцы из самых молодых групп размножения (5 и 8) сильнее всего отличаются от остальных по своим абсолютным размерам, и очень близки к представителям родительской группы размножения по качественным, дискретным признакам и пропорциям тела.
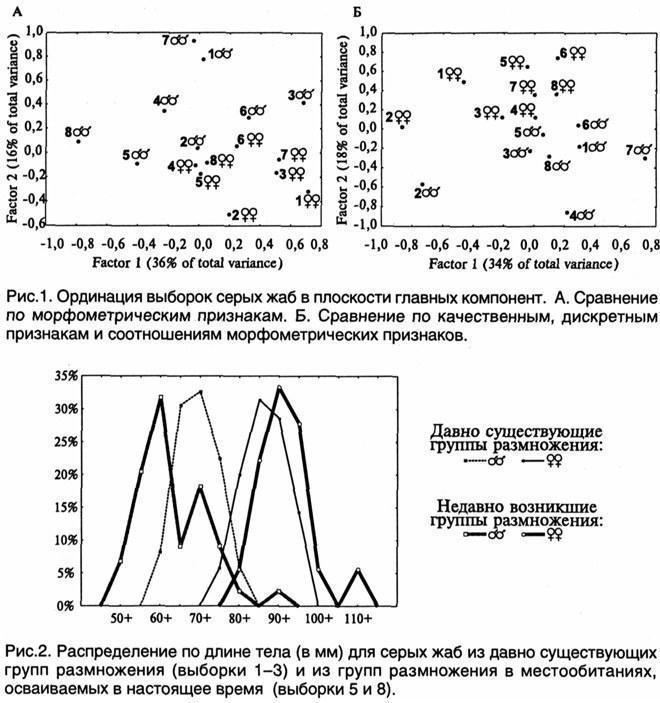
Молодые группы размножения высокодостоверно отличаются по размерам тела составляющих их особей (рис. 2). В заселяемых местообитаниях в нересте участвуют (и входят в состав образующихся пар) приходят весьма мелкие самцы (размер тела 45—60 мм). Возможно, это молодые особи, не участвующие в нересте стабильных популяций. Второй пик встречаемости самцов в «молодых» группах размножения (65—70 мм) соответствует их характерному размеру в давно заселенных местообитаниях. Парадоксальным образом участие в нересте самцов-«недоростков» сочетается с более крупными размерами самок. Поскольку в заселяющемся верхнем пруду в Пятихатках (выборка 5) на нерест одновременно пришли и мелкие самцы, и крупные самки, они сходны по своему возрасту. Вероятно, эти самки характеризовались особо быстрым ростом. Таким образом, можно заключить, что при заселении новых местообитаний у самцов серых жаб происходит ускорение созревания, а у самок — роста. Обнаруженный феномен нуждается в дальнейшем изучении (в частности, определении возраста и скорости роста жаб).
Авторы выражают благодарность А. В. Коршунову, М. А. Кравченко и Т. С. Фоменко за помощь в сборе материала и полевых наблюдениях.