Поскольку обсуждение первой модели Касума Ахмедли стало очень большим, а его новая модель дает новую отправную точку для обсуждения, выношу ее на отдельную страницу. Вот его письмо.
Дорогой Дмитрий!
Итак, я составил новую (химико-кинетическую) модель, основываясь на следующих предположениях. Гермафродиты имеют пару гомологичных хромосом HH, поддерживающих одновременно как оогенез, так и сперматогенез (не препятствующих им). В какой-то момент происходит мутация, добавляющая в хромосому H участок, блокирующий оогенез, и превращающая ее, таким образом, в хромосому Y. Мутантные особи HY по этой причине становятся самцами. В другой момент у гермафродитов HH возникает другая мутация, добавляющая в хромосому H другой участок, с превращением ее в хромосому X, которая, будучи в гомологичной паре XX, блокирует сперматогенез. Поскольку особи HX – по-прежнему гермафродиты, они могут произвести на свет самок XX. Таким образом, сочетания HH и HX определяют гермафродитов, HY и XY – это самцы, а XX – это самки. Сочетание YY оказывается невозможным, ввиду того, что нет ни гермафродитов, ни самок с хромосомой Y, которые могли бы произвести яйцеклетки с ней.
Эта модель во многом отличается от прежней. В частности, в ней нет димерного субстрата, а только мономерный S, из каждой единицы которого может образоваться 1 хромосомная единица (1 на гамету, 2 на зиготу) или, соответственно 1 единица которого образуется при разрушении 1 хромосомной единицы. Кроме того, я заменил непрерывную мутацию ступенчатой и делаю это вручную, добавляя в систему, близкую к равновесной, предполагаемого мутанта и наблюдая за дальнейшими изменениями в ней (к сожалению, используемая программа расчета не позволяет производить ступенчатые изменения автоматически). Также я убрал процессы митоза, как не имеющие прямого отношения к размножению. Но взамен этого яйцеклетки теперь образуются путем включения в их состав запаса субстрата, необходимого для первого деления после оплодотворения. Вот как выглядит схема протекающих процессов:
Таблица 1. Схема процессов
|
N |
Процесс |
k-> |
Примечания |
|---|---|---|---|
|
|
Мейоз |
|
|
|
1 |
HH + 4*S→ 2*SSHe |
Kho |
Оогенез |
|
2 |
HX + 4*S→ SSHe + SSXe |
Kho |
|
|
3 |
XX + 4*S→ 2*SSXe |
Kxo |
|
|
4 |
HH + 4*S→ 6*H |
Khs |
Сперматогенез |
|
5 |
HX + 4*S→ 3*H + 3*X |
Khs |
|
|
6 |
HY + 4*S→ 3*H + 3*Y |
Kys |
|
|
7 |
XY + 4*S→ 3*X + 3*Y |
Kys |
|
|
|
Оплодотворение |
|
|
|
8 |
SSHe + H→ 2*HH |
Kf |
|
|
9 |
SSHe + X→ 2*HX |
Kf |
|
|
10 |
SSHe + Y→ 2*HY |
Kf |
|
|
11 |
SSXe + H→ 2*HX |
Kf |
|
|
12 |
SSXe + X→ 2*XX |
Kf |
|
|
13 |
SSXe + Y→ 2*XY |
Kf |
|
|
|
Деградация |
|
|
|
14 |
HH→ 2*S |
Kd |
|
|
15 |
HX→ 2*S |
Kd |
|
|
16 |
HY→ 2*S |
Kd |
|
|
17 |
XX→ 2*S |
Kd |
|
|
18 |
XY→ 2*S |
Kd |
|
|
19 |
SSHe→ 3*S |
Kdh |
|
|
20 |
SSXe→ 3*S |
Kdx |
|
|
21 |
H→ S |
Kds |
|
|
22 |
X→ S |
Kds |
|
|
23 |
Y→ S |
Kds |
|
Сперматогенез, как видите, тоже идет с потреблением того же количества субстрата, что и оогенез, но этот субстрат не включается в сперматозоиды в качестве запаса, а расходуется на производство большего числа сперматозоидов, чем яйцеклеток. Остальные процессы, мне кажется, не требуют разъяснений.
Как и раньше, за единицу количества материала (1 P) принято начальное количество клеток с набором HH, а за единицу времени (1 L) - средняя продолжительность жизни клетки HH (не подвергнувшейся мейозу).
В табл. 2 приведены выбранные значения констант скоростей процессов.
Таблица 2. Константы скоростей
|
Процессы |
Константы скорости |
|
|
Оогенез гермафродитов |
Kho |
1.00E-04 |
|
Оогенез у самок |
Kxo |
1.70E-04 |
|
Сперматогенез у гермафродитов |
Khs |
7.00E-05 |
|
Сперматогенез у самцов |
Kys |
1.70E-04 |
|
Оплодотворение |
Kf |
1.00E+04 |
|
Деградация зигот |
Kd |
1 |
|
Деградация ооцитов гермафродитов |
Kdh |
100 |
|
Деградация ооцитов самок |
Kdx |
50 |
|
Деградация сперматозоидов |
Kds |
100 |
Таблица 3. Начальные условия
|
|
10 |
|
|
1 |
Прокомментирую выбранные значения констант скоростей. Значение Kd=1 следует из соглашения о единице времени. Это значит, что 1 диплоидная клетка разрушается в среднем за время 1 L. Гаплоидные клетки разрушаются в 100 раз быстрее (за 0.01 L), что обеспечивает соответственно более низкую концентрацию гамет. Для ооцитов самок выбрано вдвое большее время жизни, чем для ооцитов гермафродитов, что, предположительно, достигается ими за счет экономии сил, не затрачиваемых на производство сперматозоидов, и выглядит логично. Константы скоростей мейоза и оплодотворения выбраны так, чтобы при выбранных начальных условиях скорости мейоза и оплодотворения были того же порядка, что и скорость деградации зигот. Скорость сперматогенеза гермафродитов выбрана на 30% ниже скорости оогенеза у них же. А константа скорости сперматогенеза у самцов равна сумме констант обоих видов мейоза у гермафродитов, что означает, что и гермафродиты, и самцы тратят на мейоз одинаковые силы, но дает самцам численное преимущество сперматозоидов. Самки производят ооциты с такой же скоростью, что и гермафродиты, а то, что они вкладывают при этом столько же сил, проявляется в большей продолжительности жизни (большей жизнеспособности) их ооцитов.
Перейдем теперь к результатам моделирования.
Раунд 1
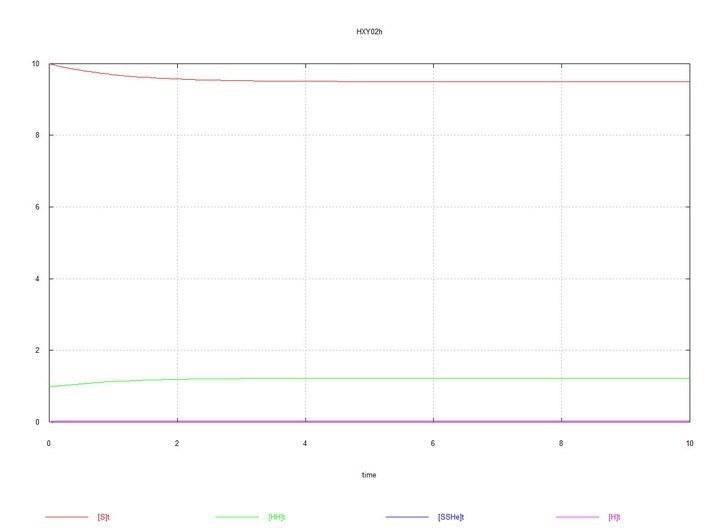
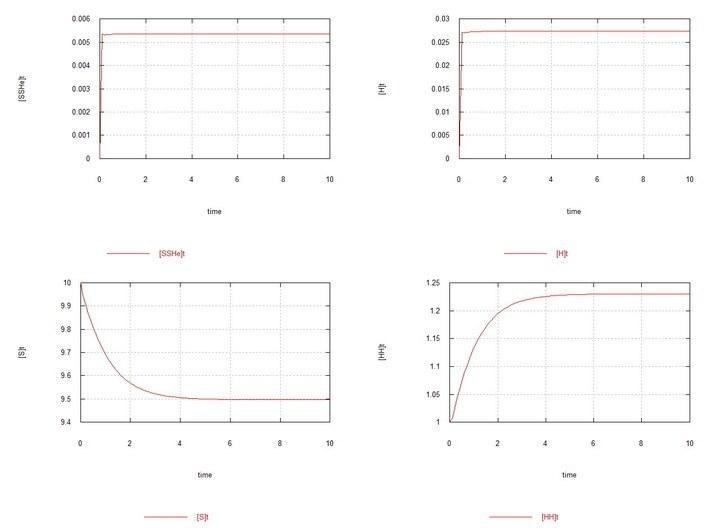
Рис. 1 (Fig01h) и Рис. 2 (Fig02h) – Равновесие в системе гермафродитов HH.
Графики системы, состоящей из одних гермафродитов HH привожу только с целью начального контроля – чтобы видеть устанавливающиеся в системе равновесные концентрации зигот и гамет.
Раунд 2
В систему гермафродитов, где 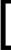 S
S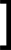 0 = 10 и HH0 = 1, добавляется HY0 = 0.01. Система движется к равновесию с перевесом гермафродитов HH и равновесной концентрацией самцов HY около 15%.
0 = 10 и HH0 = 1, добавляется HY0 = 0.01. Система движется к равновесию с перевесом гермафродитов HH и равновесной концентрацией самцов HY около 15%.
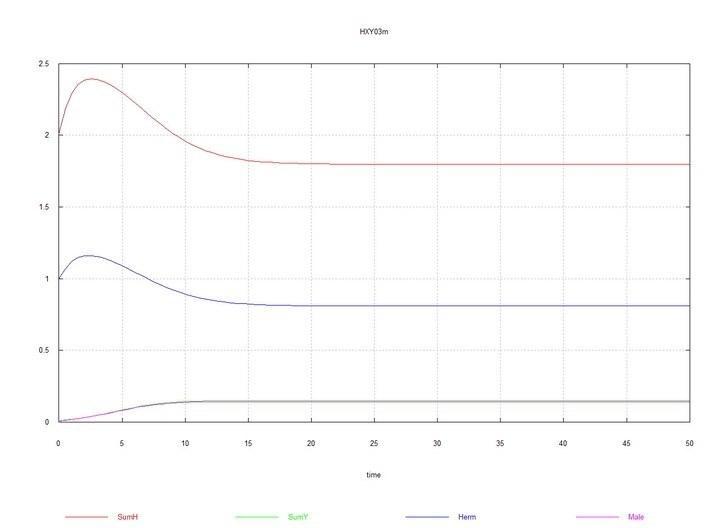

Рис. 3 и 4. (Fig03m и Fig04m) – Кинетические кривые при добавлении в среду гермафродитов 1% мутантов-самцов HY. SumH = 2*HH + HY + SSHe + H; SumY = HY + Y; Herm = HH; Male = HY.
Здесь, как было сказано выше и показано в табл. 2, константа скорости сперматогенеза самцов Kys принята равной 1.7e-4. Если ее оставить равной 1e-4, т.е. такой же, как у гермафродитов, то самцы и хромосома Y вскоре полностью вытесняются гермафродитами HH.
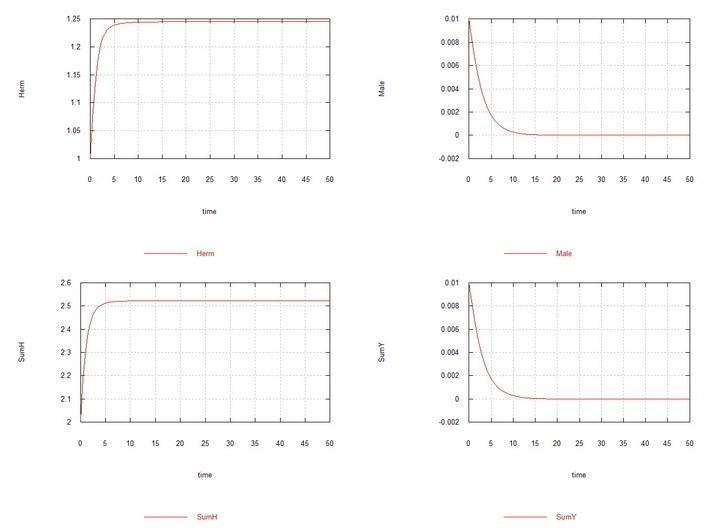
Рис. 5. (Fig09m) – Условия аналогично Fig04m с тем отличием, что Kys выбрано равным 7e-5, т. е. константы скорости сперматогенеза у гермафродитов HH и самцов HY одинаковы. Демонстрируется вытеснение самцов при этих условиях.
Раунд 3
В систему гермафродитов, где S0 = 10 и HH0 = 1, добавляется HX0 = 0.01. Система движется к равновесию с перевесом гермафродитов и равновесной концентрацией самок FF около 20%.

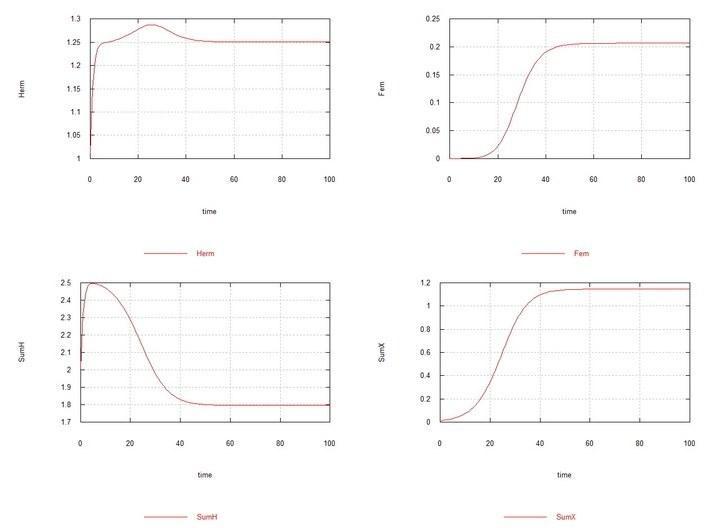
Рис. 6 и 7. (Fig05f и Fig06f) – Кинетические кривые при добавлении в среду гермафродитов 1% мутантов-гермафродитов HX. SumH = 2*HH + HX + SSHe + H; SumX = HX + 2*XX + SSXe + X; Herm = HH + HX; Fem = XX.
Здесь константа Kdx скорости деградации ооцитов самок SSXe выбрана равной 50 (табл. 2). Если же выбрать ее равной 100, как для ооцитов гермафродитов SSHe, то и в этом случае хромосома X вскоре полностью вытесняется хромосомой H.
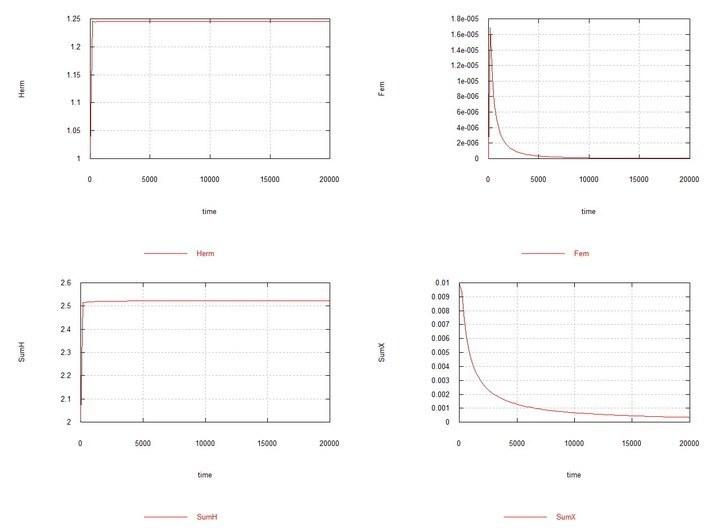
Рис. 8. Fig10f – Условия аналогично Fig06f с тем отличием, что Kdx выбрано равным 100, т. е. константы скорости деградации ооцитов у гермафродитов и самок одинаковы. Демонстрируется вытеснение самок при этих условиях.
Раунд 4
В около-равновесную систему гермафродиты- самцы с соотношением HH0 = 0.8, HY = 0.2 добавляется HX0 = 0.01. Самцы XY и самки XX постепенно полностью вытесняют гермафродитов обоих видов и самцов HY.
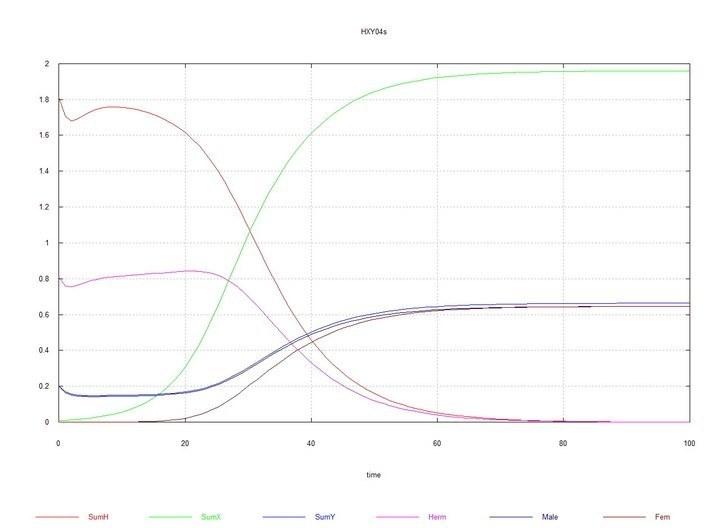
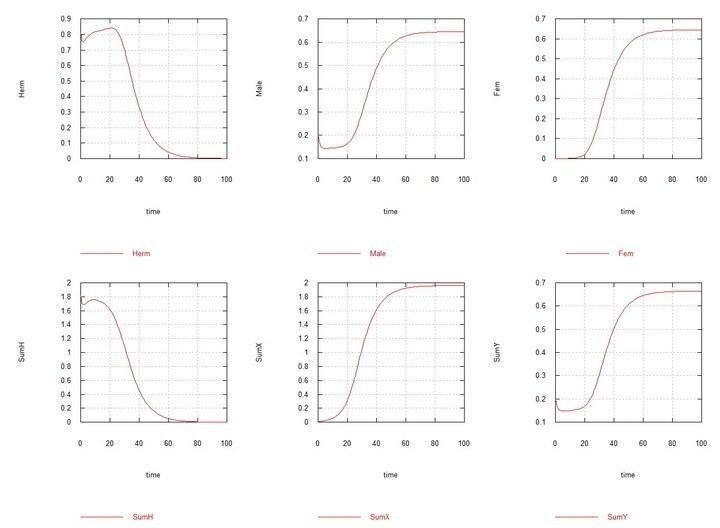
Рис. 9 и 10 (Fig07s и Fig08s) – Кинетические кривые при добавлении 1% гермафродитов HX в равновесную систему гермафродитов (HH 80%) и самцов (HY 20%). SumH = 2*HH + HX + HY + SSHe + H; SumX = HX + 2*XX + XY + SSXe + X ; SumY = HY + XY + Y; Herm = HH + HX; Male = HY + XY; Fem = XX.
Выводы
В результате моделирования получилось, что появление в популяции гермафродитов HH мутанта HY, обладающего прежней скоростью сперматогенеза (т.е. как у HH), мутанты быстро вытесняются гермафродитами. Однако, если самцы HY тратят на сперматогенез столько же сил, сколько гермафродит на оба вида гамет суммарно, то устанавливается некоторое равновесие, в котором, все же, гермафродиты значительно превосходят в численности самцов. Это ожидаемо, поскольку, самцы в своем потомстве имеют равное число самцов и гермафродитов, а гермафродиты между собой имеют еще и дополнительное количество потомков-гермафродитов.
При появлении в популяции гермафродитов HH мутантных гермафродитов HX, и присвоении самкам XX такой же способности производить яйцеклетки, как и гермафродитам HH и HX, хромосома X так же быстро вытесняется хромосомой H, как это происходило со слабой хромосомой Y. Но если самкам XX присваивается способность производить более жизнеспособные ооциты (за счет того, что они тратят на их производство столько же сил, сколько гермафродиты тратят на производство обоих типов гамет в сумме), то и здесь устанавливается равновесие, в котором численно преобладают гермафродиты (и, как легко видеть, по тем же причинам).
Однако, когда в равновесной системе гермафродиты-самцы HH-HY (с преобладанием, как было сказано, гермафродитов HH), появляется мутант HX, то происходит дивергентный рост численности самцов XY и самок XX, полное вытеснение гермафродитов всех видов и самой хромосомы H, и устанавливается равновесие с равной численностью самцов и самок.
P.S. Возможно, вместо разговора о H, X, Y, как о хромосомах, следовало бы говорить о них, как об аллелях одного гена, как делаете это Вы, но я исходил из того, что гены, определяющие мужской и женский пол – это разные гены, и ген на хромосоме Y антагонистичен к самому себе и не способен к образованию пары YY. Впрочем, все это для меня не очень ясно, а Вы, как специалист в этой области, можете судить намного компетентней.