Боркин Л. Я., Безман-Мосейко О. С., Мазепа Г. А., Зиненко А. И., Коршунов А. В., Лада Г. А., Шабанов Д. А., Литвинчук С. Н., Розанов Ю. М. О южной границе распространения гибридной Rana esculenta (Ranidae, Anura, Amphibia) на территории Украины и Молдовы: данные проточной ДНК-цитометрии // Праці Українського герпетологічного товариства. – № 1, 2008. – С. 3-8.
УДК 595.6, 597.94
О ЮЖНОЙ ГРАНИЦЕ РАСПРОСТРАНЕНИЯ ГИБРИДНОЙ RANA ESCULENTA (RANIDAE, ANURA, AMPHIBIA) НА ТЕРРИТОРИИ УКРАИНЫ И МОЛДОВЫ: ДАННЫЕ ПРОТОЧНОЙ ДНК-ЦИТОМЕТРИИ
Л. Я. Боркин1, О. С. Безман-Мосейко1, Г. А. Мазепа2, А. И. Зиненко3, А. В. Коршунов2, Г. А. Лада4, Д. А. Шабанов2, С. Н. Литвинчук5, Ю. М. Розанов5
1 3оологический институт Российской академии наук, Университетская набережная, 1, Санкт-Петербург, 199034 Россия; E-mail: lacerta@zin.ru
2 Харьковский национальный университет им. В. Н. Каразина, пл. Свободы, 4, Харьков, 61077 Украина; E-mail: d_sh@i.ua
3 Музей природы Харьковского национального университета им. В.Н. Каразина, ул. Тринклера, 8, Харьков, 61022 Украина; E-mail: zinenkoa@yahoo.com
4 Тамбовский государственный университет, ул. Интернациональная 33, Тамбов, 392000 Россия; E-mail: esculenta@mail.ru
5 Институт цитологии Российской академии наук, Тихорецкий пр., 4, Санкт-Петербург, 194064 Россия; E-mail: slitvinchuk@yahoo.com
О южной границе распространения гибридной Rana esculenta (Ranidae, Anura, Amphibia) на территории Украины и Молдовы: данные проточной ДНК-цитометрии. Боркин Л. Я., Безман- Мосейко О. С., Мазепа Г. А., Зиненко А. И., Коршунов А. В., Лада Г. А., Шабанов Д. А., Литвинчук С. Н., Розанов Ю. М.— С помощью проточной ДНК-цитометрии было выявлено распространение гибридогенного вида Rana esculenta на территории Молдовы и Украины. Согласно полученным данным, южная граница распространения R. esculenta может быть проведена с запада на восток через следующие пункты: Вилково (дельта р. Дунай), Вулканешты (Молдова), Дойбаны, Тирасполь и Кицканы (р. Днестр), Березово (р. Тилигул), Голопристанский р-н (р. Днепр), Русский Орчик (Харьковская обл., Украина) и вдоль средней части реки Северский Донец от северо-востока Харьковской обл. (Украина) до Ростовской обл. (Россия). В подавляющем большинстве случаев R. esculenta была представлена только диплоидными особями, а популяционные системы RE-типом. Только в Русском Орчике нами была достоверно обнаружена REL-система. Находки R. esculenta в Херсонской обл. географически изолированы от популяций, населяющих северную и центральную части Украины. Однако этот большой разрыв в распространении, по-видимому, скорее связан с плохой изученностью данной территории, чем с реликтовой природой этих наиболее южных популяций.
Ключевые слова: Rana esculenta, размер генома, Украина, Молдова
On the Southern Limit of Range of Hybrid Rana esculenta (Ranidae, Anura, Amphibia) in Ukraine and Moldova: DNA Flow Cytometry Evidence. Borkin L. J., Bezman-Moseiko O. S., Mazepa G. A., Zinenko A. I., Korshunov A. V., Lada G. A., Shabanov D. A., Litvinchuk S. N., Rozanov J. M.— Based on DNA flow cytometry, we identified green frog localities with hybrid Rana esculenta. We found or confirmed some southernmost localities of the species situated in Moldavia and Ukraine. According to our data, the southern distributional border of R. esculenta is demarcated as follows from the west to the east: Vilkovo (Danube River Delta), Vulcanesti (Moldova), Doibani, Tiraspol', and Kitscani (Dniestr River), Beryozovo (Tiligul River), Golaya Pristan District (Dnieper River), Russky Orchik (Kharkov Province, Ukraine) as well as along the middle part of Seversky Donets River in north-eastern Ukraine and Rostov Province in adjacent Russia. The majority of samples contained diploid hybrid frogs from the RE kind of population systems. The REL system was identified in Russky Orchik. The records of R. esculenta in Kherson Province of Ukraine are geographically far isolated from populations in northern and central parts of Ukraine. However, this large gap seems to be resulted from inadequate frog sampling and species identification rather than from relic nature of the southernmost records.
Введение
В последние десятилетия гибридогенный комплекс Rana esculenta привлекает к себе пристальное внимание исследователей разных стран. Выяснилось, что он обладает необычными эволюционно-генетическими механизмами, изучение которых имеет большое значение для понимания процессов видообразования, причем не только среди амфибий. Это, в первую очередь, гибридизация, не-менделевское (мероклональное) наследование, полиплоидия и разнообразные типы популяционных систем, включающие как однополых (или самцы, или самки), так и двуполых гибридов.
Распространение гибридогенного комплекса на территории европейской части бывшего СССР изучено явно недостаточно. Во многом это обусловлено ненадежностью определения видов по традиционным диагностическим признакам, поскольку пределы изменчивости пропорций тела и окраски у гибридной съедобной лягушки(Rana esculenta Linnaeus, 1758) и у родительских видов (Rana lessonae Camerano, 1882 и Rana ridibunda Pallas, 1771) заметно перекрываются, в особенности у триплоидных особей (Planeretal., 1994; Ladaetal., 1995; Schruer, 1996; Борисовский и др., 2001; Ручин и др., 2005). Массовая встречаемость полиплоидов (3nи 4n) недавно была обнаружена на северо-востоке Украины и в Ростовской области России (Borkin et al., 2004, 2006; Боркин и др., 2005). В связи с вышесказанным возникает вопрос о достоверности литературных данных, полученных при использовании внешних морфологических признаков.
В настоящее время адекватное определение видов, входящих в гибридогенный комплекс, возможно с помощью таких биохимических методов, как электрофорез белков (см. Межжерин и др., 2007) и ДНК-цитометрия (см. Боркин и др., 1987; Borkin et al., 2004). В последние десятилетия нами было изучено большое число зеленых лягушек из многих районов европейской части бывшего СССР. Подавляющая часть выборок была исследована с помощью проточной ДНК-цитометрии, позволяющей не только точно определить видовую принадлежность каждой особи, но также установить ее плоидность, а для самцов еще и тип гамет. В случае полиплоидии можно также определить геномный состав каждой особи. С учетом этих данных нам удалось в значительной степени уточнить распространение R. esculenta. В данной статье мы представляем наши материалы, характеризующие южную границу ареала этого гибридного вида на территории Украины и Молдовы.
Материал и методы
Видовую идентификацию и уровень плоидности лягушек осуществляли по размеру генома, определенному методом проточной ДНК-цитометрии (Borkin et al., 2004). После анестезии эритроциты брали непосредственно из сердца. Тестируемые клетки смешивались с клетками репера и анализировались совместно. Поэтому в такой смеси как изучаемые клетки, так и клетки репера красили и измеряли в одних и тех же условиях. В качестве репера использовались эритроциты травяных лягушек (R. temporaria), собранных в Ленинградской и Псковской областях России. Детали этого метода были описаны ранее (Vinogradov et al., 1990; Розанов, Виноградов, 1998; Borkin et al., 2001).
Результаты и обсуждение
Среди 1497 изученных особей зеленых лягушек, отловленных в 131 пункте, расположенном на территории Молдовы и Украины, гибридными оказались 774 особи из 76 мест. Наиболее южными из них были (рис. 1): г. Вилково (Килийский р-н, Одесская обл., Украина), с. Вулканешты (Ниспоренский р-н, Молдова), с. Дойбаны 2-е (Дубоссарский р-н, Приднестровье, Молдова), г. Тирасполь (Тираспольский р-н, Приднестровье, Молдова), с. Кицканы (Слободзейский р-н, Приднестровье, Молдова), Березовский лес (Березовский р-н, Одесская обл., Украина), с. Подтёмное и с. Рыбальче (Голопристанский р- н, Херсонская обл., Украина), урочище Русский Орчик (Зачепиловский р-н, Харьковская обл., Украина), с. Великая Гомольша (Змиёвский р-н, Харьковская обл., Украина), с. Крейдянка (Балаклейский р-н, Харьковская обл., Украина), пос. Червоный Шахтер и с. Яремовка (Изюмский р-н, Харьковская обл., Украина), г. Святогорск (Святогорский р-н, Донецкая обл., Украина), с. Станично-Луганское (Станично-Луганский р-н, Луганская обл., Украина).
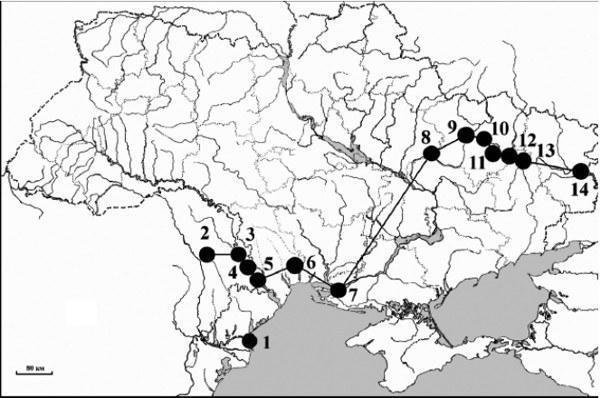
Рис. 1. Южная граница распространения Rana esculenta на территории Украины и Молдовы. 1 — Вилково; 2 — Вулканешты; 3 — Дойбаны 2-е; 4 — Тирасполь; 5 — Кицканы; 6 — Березовский лес; 7 — Голопристанский р-н; 8 — Русский Орчик; 9 — Великая Гомольша; 10 — Крейдянка; 11 — Червонный Шахтер; 12 — Яремовка; 13 — Святогорск; 14 — Станично-Луганское.
В кандидатской диссертации Н. М. Сурядной (2005, с. 277 и рис. 7.9) представлен перечень находок R. esculenta на территории Украины. Помимо основного массива точек на севере и в центральной части страны, этим автором показаны также два района, географически изолированные друг от друга и от основного массива местонахождений вида. Это — дельта р. Дунай (Вилково) и низовья р. Днепр (села Старая Збурьевка, Забарино и Геройское Голопристанского р-на Херсонской обл.). Примерно такой же тип распространения изображен и для прудовой лягушки R. lessonae (кроме пункта Геройское). Идентификация обоих видов была сделана автором по внешней морфологии и частично по морфометрии хромосом. Наши данные по проточной ДНК-цитометрии подтверждают обитание R. esculenta в этих районах. Следует также заметить, что гибриды из дельты р. Дунай были обнаружены еще ранее с помощью электрофореза белков (Gatheretal., 1991; Межжерин и др., 2007).
Для территории Молдовы ряд авторов (см. Borkin et al., 1997) указывал два вида зеленых лягушек: R. ridibunda и R. «esculenta» (= R. lessonae). Любопытно, что в кандидатской диссертации В. Е. Тофана (1966) последний вид был указан для с. Суклея Тираспольского р-на (Приднестровье), с. Карбуна Ново-Аненского р- на, сел Быковец и Чутешты Каларашского р-на и с. Филипены Леовского р-на (Молдова). Однако ранее диплоидные гибриды были найдены нами лишь в окр. с. Вулканешты Ниспоренского р-на (Borkinetal., 1997). Rana lessonae здесь обнаружить не удалось. В 2006 г. в выборке из 36 особей, пойманных в г. Тирасполь, были идентифицированы 2 диплоидных гибрида; все остальные особи оказались R. ridibunda. Специальное обследование, предпринятое нами в 2007 г., показало довольно широкое распространение R. esculenta в Слободзейском, Тираспольском и Дубоссарском районах Приднестровья; однако R. lessonae найти не удалось. Все гибридные экземпляры (n= 31) оказались также диплоидами. Таким образом, эти наши данные позволяют провести южную границу распространения R. esculenta по низовьям р. Днестр.
В 2007 г. на р. Тилигул в Березовском лесу (Березовский р-н, Одесская обл., Украина) были отловлены 10 особей, 9 из которых оказались диплоидными гибридами, а одна особь — R. ridibunda. Особенностью этой находки можно считать то, что она не принадлежит к бассейнам рек Днестр и Днепр и занимает географически промежуточное положение между ними. Отметим, что достоверные находки многочисленных гибридов в Приднестровье и бассейне р. Тилигул географически связывают R. esculenta из таких удаленных друг от друга районов, как дельты рек Дунай и Днепр.
В 2007 г. нам также удалось доказать обитание R. esculenta (3 диплоида) в двух пунктах (Подтемное и Рыбальче) Голопристанского р-на Херсонской обл. Кроме того, по акустическим и внешним морфологическим признакам этот вид был идентифицирован в окрестностях сел Коханы (Голопристанский р-н) и Песчановка (Цюрупинский р-н). В последнем пункте некоторые экземпляры зеленых лягушек были определены нами как R. lessonae. Ранее в других местах этой же области (Херсон, Голая Пристань и Цюрупинск) нами была отмечена только R. ridibunda.
Следующие наши цитометрические данные относятся к северо-восточной Украине. Этот регион, как это теперь хорошо известно, характеризуется широким распространением R. esculenta. Уникальной особенностью его (в рамках европейской части бывшего СССР) является массовая полиплоидия среди гибридов, найденных вдоль среднего течения реки Северский Донец на протяжении 480 км (Borkin et al., 2004, 2006; Боркин и др., 2005). Из целого ряда новых точек, которые нами были обследованы, две в контексте данной статьи заслуживают особого внимания. В урочище Русский Орчик (Капинерное озеро, с. Заречье и с. Даманский) нами были идентифицированы все три вида комплекса R. esculenta (все гибриды диплоидные), что подтверждает прежние данные В. И. Ведмедери (1984) и Г. А. Лады (Borkinetal., 2004), основанные на исследовании внешних признаков. В другом местонахождении (с. Яремовка) были отловлены 4 особи, которые по размеру генома были классифицированы как триплоидные гибриды RRL (n= 3) и RLL (n= 1) типов. Другие новые находки, о которых будет сказано в отдельной статье, не выходили за известные ранее географические рамки.
Наиболее юго-восточная находка R. esculenta была сделана нами в России на территории Ростовской обл., примыкающей к восточной части Украины (Луганская обл.). Ближайшая находка этого вида на территории России была известна до этого в 320 км к северу (Borkin et al., 2006).
Для полноты картины здесь важно упомянуть еще о нескольких находках зеленых лягушек на территории Украины, которые по внешним признакам были идентифицированы как R. esculenta или R. lessonae: с. Крымка (Первомайский р- н, Николаевская обл.; R. esculenta — Кузьмин, 1999), с. Александровка (Вознесенский р-н, Николаевская обл.;R. lessonae — наши данные) и Черный лес у с. Знаменка (Знаменский р-н, Кировоградской обл.; R. esculenta, R. lessonae и R. ridibunda — наши данные).
Таким образом, подводя итог вышесказанному, можно заключить, что южная граница ареала съедобной лягушки (как, видимо, и прудовой) проходит параллельно границе Лесостепи и Степи, смещаясь вместе с ней к северу по мере увеличения континентальности климата и заходя далеко на юг вдоль таких крупных рек как Дунай, Днестр и Днепр.
В рамках анализа популяционных систем следует отметить, что подавляющее большинство выборок, расположенных на южной границе ареала R. esculenta, следует отнести к RE-типу. Исключение составляют популяции из урочища Русский Орчик и, вероятно, Вилково (REL-тип). Во всех наиболее южных местообитаниях гибриды, как правило, были представлены только самцами, исключение составляет северо-восток Украины, где популяции R. esculenta могут характеризоваться обоими полами или только самками.
Исходя из имеющихся сведений (Сурядная, 2005, рис. 7.9; Писанец, 2007, рис. 100; наши данные), на территории Украины существует «разрыв» в распространении R. esculenta между северными и центральными районами, с одной стороны, и самым югом, с другой. Однако, на наш взгляд, эта «картографическая дизъюнкция» обусловлена скорее отсутствием адекватных сборов и их правильной идентификацией, нежели только реликтовой природой южных популяций. Поэтому это «белое пятно» может исчезнуть в будущем при более тщательном изучении распространения зеленых лягушек.
Авторы выражают благодарность В. Г. Дядичко (Одесский филиал ИНБЮМ им. А.О. Ковалевского НАН Украины) за помощь при сборе материала. С российской стороны работа была поддержана грантами НШ-4212.2006.4, РФФИ 05—04—48403 и Центра коллективных исследований «Материаловедение и диагностика в передовых технологиях», с украинской стороны — грантом INTAS Ref. Nr05-1000008-8147.
Борисовский А. Г., Боркин Л. Я., Литвинчук С. Р., Розанов Ю.М. Морфометрическая характеристика зеленых лягушек (комплекс Rana esculenta) Удмуртии // Вестн. Удмурт. ун-та. — 2000. — № 5. - С. 70-75.
Боркин Л. Я., Виноградов А. Е., Розанов Ю. М, Цауне И. А. Полуклональное наследование в гибридогенном комплексе Rana esculenta: доказательство методом проточной ДНК-цитометрии // Докл. АН СССР. - 1987. -295, № 5. - С. 1261-1264.
Боркин Л. Я., Зиненко А. И., Коршунов А. В. и др. Массовая полиплоидия в гибридогенном комплексе Rana esculenta (Ranidae, Anura, Amphibia) на востоке Украины // Матеріали Першої конф. Українського герпетол. тов-ва (10-12 жовтня 2005 р., Київ). - К. : Зоомузей ННПМ НАН України, 2005. - С. 23-26.
Ведмедеря В. И. Некоторые данные о лягушках рода Rana в Харьковской области (по материалам Музея природы ХГУ) // Вестн. Харьков. ун-та. - 1984. - № 262. - С. 99-101.
Кузьмин С. Л. Земноводные бывшего СССР. - М. : Товарищество научных изданий КМК, 1999. - 298 с.
Межжерин С. В., Морозов-Леонов С. Ю., Некрасова О. Д., Куртяк Ф. Ф. и др. Эволюционно- генетические аспекты полуклонального воспроизводства гибридной формы Rana kl. esculenta (Amphibia, Ranidae) // Наук. вісн. Ужгород. ун-ту. Сер. биол. - 2007. - Вип. 1. - С. 79-84.
Писанец Е. М. Амфибии Украины. Справочник-определитель земноводных Украины и сопредельных территорий. - Киев : Зоол. музей ННПМ НАН Украины, 2007. - 312 с.
Розанов Ю. М, Виноградов А. Е. Прецизионная ДНК-цитометрия: исследование индивидуальной вариабельности размера генома животных // Цитология. - 1998. -40, № 8/9. - С. 792-799.
Ручин А. Б., Боркин Л. Я., Лада Г. А. и др. Морфологическая изменчивость, размер генома и популяционные системы зеленых лягушек (Rana esculenta complex) Мордовии // Бюл. Моск. об-ва испыт. прир. Отд. биол. - 2005. -110, вып. 2. - С. 3-10.
Сурядная Н. Н. Зеленые лягушки фауны Украины: морфологическая изменчивость, кариология и особенности биологии : Дис. ... канд. биол. наук. - Киев, 2005. - 27 с.
Тофан В. Е. Фауна земноводных и пресмыкающихся Молдавии : Дис. ... канд. биол. наук. - Л.; 1966. - 350 с.
Borkin L. J., Korshunov A. V., Lada G. A. et al. Mass occurrence of polyploid green frogs (Rana esculenta complex) in eastern Ukraine // Russ. J. Herpetol. - 2004. - 11, N 3. - P. 203-222.
Borkin L. J., Lada G. A., Litvinchuk S. N. et al. The first record of mass polyploidy in hybridogenetic green frog Rana esculenta in Russia (Rostov Oblast') // Russ. J. Herpetol. - 2006. -13, N 1. - P. 77-82.
Borkin L. J., Litvinchuk S. N, Rosanov Y. M. Amphibians and reptiles of Moldavia: additions and corrections, with a list of species // Russ. J. Herpetol. — 1997. —4, N 1. — P. 50—62.
Borkin L. J., Litvinchuk S. N, Rosanov J. M, Milto K.D. Cryptic speciation in Pelobates fuscus (Anura, Pelobatidae): evidence from DNA flow cytometry // Amphibia—Reptilia — 2001. —22, N 4. — P. 387-396.
Gbuther R., Plotner J., Tetzlaff I. Zu einigen Merkmalen derWasserfrasche (Rana synkl. esculenta) des Donau-Deltas // Salamandra. - 1991. -27, H. 4. - S. 246-265.
Lada G. A., Borkin L. J., Vinogradov A. E.Distribution, population systems and reproductive behavior of green frogs (hybridogenetic Rana esculenta complex) in the Central Chernozem Territory of Russia // Russ. J. Herpetol. - 1995. -2, N 1. - P. 46-57.
Plotner J., Becker C., Plutner K. Morphometric and DNA investigations into European water frogs (Rana kl. esculenta Synklepton (Anura, Ranidae)) from different population systems // J. Zool. Syst. Evol. Research. - 1994. -32. - P. 193-210.
SchruerT. Morphologie und Ploidiegrade von Wasserfraschen aus unterschiedlichen Populationssystem in Nordost-Polen // Zeitschrift &r Feldherpetologie. - 1996. -3. - S. 133-150.
Vinogradov A. E, Borkin L. J., Gbnther R., Rosanov J. M. Genome elimination in diploid and triploid Rana esculenta males: cytological evidence from DNA flow cytometry // Genome. - 1990. -33, N 5. - P. 619-627