Українська мова (найновіша версія) / Русский язык (обновление прекращено)
4.13. Конкуренция и экологические ниши
Как мы установили, конкуренцией называется такое отношение между двумя популяциями, при котором увеличение каждой из них вызывает снижение численности другой. Впрочем, это определение касается только межвидовой конкуренции, а кроме нее существует также внутривидовая конкуренция, которая разворачивается внутри одной популяции.
При конкуренции две популяции могут неблагоприятно влиять друг на друга двумя разными способами. Первый (косвенная, или эксплуатационная конкуренция) связан с тем, что две популяции используют один и тот же ресурс. При возрастании численности одной из этих популяций усилится потребление ею общего ресурса, и другой достанется меньшее его количество. Второй (прямая, или интерференционная конкуренция) связан с затратами энергии особей на причинение друг другу ущерба. Примером прямой конкуренции может быть аллелопатия — выделение растениями веществ, угнетающих другие виды.
А зачем организмы идут на затраты энергии в ходе интерференционной конкуренции? Это явление не могло бы наблюдаться, если бы такие затраты не решали какую-то важную для конкурирующих популяций задачу. Эта задача — уменьшение ущерба от эксплуатационной конкуренции. Итак, хотя прямая конкуренция, которая может сопровождаться конфликтами между особями разных популяций, кажется более «явной» и зрелищной, она является лишь следствием скрытой, косвенной конкуренции.
Какие виды сильнее конкурируют друг с другом: подобные или отличающиеся? Чем более сходны виды, тем сильнее пересекаются их потребности в ресурсах, тем острее будет конкуренция между ними. Для описания отношения конкуренции очень полезным оказывается понятие экологической ниши.
Мы уже говорили, что многие ключевые термины употребляются в экологии в различных смыслах. Вероятно, понятие экологической ниши является в этом отношении «рекордсменом».
Впервые словосочетание «экологическая ниша» употребил Дж. Гринелл в 1917 году. Он обозначал таким образом характерное местообитание вида, совокупность условий, в которых какой-то вид встречается в природе. Ч. Эльтон в 1927 году определил экологическую нишу как место вида в сообществе, его положение в структуре пищевых взаимоотношений. Дж. Хатчинсон в 1957 году представил экологическую нишу как совокупность всех значений экологических факторов, допускающих существование вида (см. подробнее дополнение 6.3). Наконец, Ю. Одум пояснил, что ниша является характеристикой требований вида к окружающей среде и присущего ему образа жизни.
Например, мы можем оценить, как использует ресурсы среды тот или иной вид, и использовать эту оценку как срез его экологической ниши.
На рис. 4.13.1 показана двумерная ниша одной из насекомоядных птиц американских дубрав — сине-зеленого мошколова. Как вы видите, его ниша охарактеризована здесь по двум параметрам: какую добычу ловит рассматриваемый вид птиц и где (на какой высоте) он это делает. Можно предположить, что для любого другого вида птиц подобная картина выглядела бы иначе.
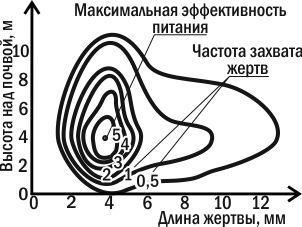
Рис. 4.13.1. Двумерная экологическая ниша сине-зеленого мошколова; показана частота захвата насекомых разного размера на разной высоте от земли
«Экологическая ниша некоторого организма зависит не только от того, где он живет, но и от того, что он делает (как преобразует энергию, каково его поведение, как он реагирует на физическую и биологическую среду и изменяет ее) и как он ограничен другими видами. Можно привести такую аналогию: местообитание — это «адрес» организма, а экологическая ниша — это, говоря биологически, его «профессия» (Одум, 1975).
Объединяя столь разнородные подходы, мы можем сказать, что экологическая ниша — это комплексная характеристика образа жизни вида, включающая потребляемые им ресурсы, а также те условия, при которых он может существовать.
Ценность понятия экологической ниши состоит в том, что оно позволяет сравнивать особенности образа жизни разных видов. Так, если два вида могут обитать в сходных условиях, но отличаются по потребляемому ими ресурсу (например, по размеру своих жертв), мы можем сравнивать ниши этих видов именно по этому параметру.
Теперь вам станет ясно, почему мы заговорили о понятии ниши, когда говорили о конкуренции. За ту часть ресурса, которую потребляет только один вид, конкуренция невозможна. Зато для той части разнообразия ресурса, которую могут употреблять два вида (то есть той части, в которой ниши этих видов пересекаются), между этими видами будет возникать конкуренция (рис. 4.13.2).
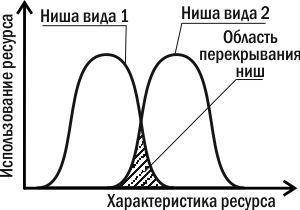
Рис. 4.13.2. Сравнение экологических ниш двух видов по тому параметру, по которому они отличаются (например, по размеру потребляемых хищниками жертв)
Чем более похожи два вида, тем сильнее пересекаются их потребности в ресурсах и тем сильнее конкуренция между ними. Самая острая конкуренция — внутривидовая. Но особи одного вида, хотя и конкурируют весьма остро за ресурсы, совместно формируют следующее поколение. А что будет, если конкуренция двух разных видов достигнет той же остроты, что и внутривидовая?
В 1931–1935 годах молодой советский биолог Георг Францевич Гаузе пытался экспериментально проверить уравнения конкуренции, выведенные В. Вольтерра. Гаузе понял, что для экспериментов в области популяционной биологии хорошо подходят простейшие: им требуется совсем небольшой объем среды, а смена их поколений происходит намного быстрее, чем у многоклеточных животных. Эксперименты Гаузе, прославившее его имя, выполнены на инфузории туфельке (Paramecium caudatum) и ее ближайших родственниках (рис. 4.13.3).
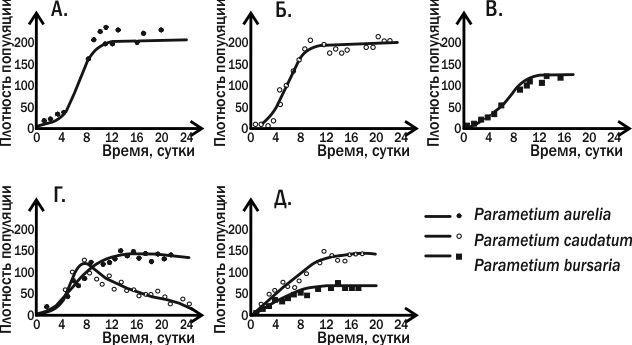
Рис. 4.13.3. Динамика численности трех видов инфузорий в опытах Г.Ф. Гаузе. В вариантах экспериментов А., Б. и В. виды выращивали поодиночке; в случаях Г. и Д. в пробирку заселяли сразу два вида инфузорий
Гаузе выращивал три вида инфузорий рода Paramecium в пробирках, куда он добавлял овсяную муку и откуда периодически удалял отходы. На овсяной муке развивались дрожжи и бактерии, которыми и питались инфузории. Три вида рода Paramecium успешно развивались в монокультуре в таких условиях. При совместном содержании P. aurelia всегда полностью вытесняла P. caudatum. Однако при совместном содержании P. aurelia и P. bursaria два вида могли существовать вместе. Как установил Гаузе, эти два вида отличаются по своему способу питания: первый вид питался бактериями в толще жидкости, второй — дрожжевыми клетками у дна.
На основании описанных наблюдений был сформулирован принцип конкурентного исключения, или правило Гаузе: два вида, занимающие одну и ту же экологическую нишу, не могут устойчиво сосуществовать в одном местообитании; сосуществование видов возможно благодаря разделению их ниш.
Правило конкурентного исключения является одной из возможных причин, объясняющих чрезвычайное разнообразие видов в природе. Если две популяции в одном местообитании не могут занимать одну нишу, им приходится ее разделять. Со временем каждая из таких популяций специализируется в своей нише и может дать начало новому специализированному виду.
Экологически проверить действие правила Гаузе в естественных условиях достаточно тяжело, однако некоторые примеры его действия биологам известны. С конца XIX века на большей части Европы широкопалый рак (Astacus astacus) вытеснен длиннопалым раком (A. leptodactylus). Экологическая ниша длиннопалого рака перекрывает нишу короткопалого, и при этом вид-победитель более плодовит. Плотва вытесняет во многих водоемах красноперку и окуня. Экологические ниши взрослых особей разобщены, а мальков — перекрываются. Мальки плотвы оказываются более конкурентоспособными. Однако наличие даже многих примеров конкурентного вытеснения не означает, что оно происходит всегда. За то время, в течение которого биологи исследуют действие правила Гаузе, удалось найти как примеры его действия, так и ситуации, когда оно не «срабатывает».
Например, в сообществе планктонных водорослей разделения ниш не обнаруживается. Причиной этого является быстро меняющаяся среда, где условия дают преимущество то одному, то другому виду, а также интенсивное воздействие со стороны хищников (зоопланктона и разнообразных фильтраторов), которые не дают конкурентному исключению стать главным фактором в формировании сообщества. Однако в некоторых случаях конкурентное вытеснение происходит и в планктонном сообществе. Это происходит при так называемом «цветении» воды, когда планктонные водоросли вырываются из-под ограничивающего их численность действия хищников. В результате «цветения» воды в ней становятся массовыми всего несколько видов планктонных водорослей, которые отличаются друг от друга по своему образу жизни.
Некоторые виды отношений могут связывать не только особей из разных популяций, но и сородичей, входящих в состав одной популяции. Такова, например, конкуренция. Действие внутривидовой и межвидовой конкуренции сходно тем, что и та и другая ограничивают количество ресурсов, доступных для особи. Однако их влияние на ширину экологических ниш противонаправлено. Внутривидовая конкуренция расширяет экологические ниши: всем особям не хватает оптимальных для вида ресурсов, и некоторые вытесняются «на край», использовать хоть как-то подходящие ресурсы. Межвидовая конкуренция, согласно правилу Гаузе, наоборот, сужает экологические ниши. Наблюдаемая нами в природе ширина ниш является, таким образом, результатом уравновешивания двух противонаправленных процессов, их расширения из-за внутривидовой конкуренции и сужения вследствие межвидовой.
Интересный пример конкурентного исключения был исследован в опытах Т. Парка с мучными хрущаками. Два вида этих жуков из семейства чернотелок (Tribolium confusum и T. castaneum) содержали в ящиках с мукой. Эти виды конкурировали за пищу (муку) и, кроме того, могли питаться друг другом, причем жуки обоих видов поедали преимущественно особей вида-конкурента, а не своего вида (такое сочетание конкуренции с хищничеством называется антагонизмом). Парку удалось подобрать условия, при которых закономерно побеждал первый или второй вид. Однако интереснее всего оказалось исследовать итог конкуренции при промежуточных условиях, не обеспечивавших безусловного перевеса одного из видов (таблица 4.13.1.).
Таблица 4.13.1. Количество побед двух видов мучных хрущаков в экспериментах Т. Парка
|
«Климат» (условия эксперимента) |
% побед |
|
|
Tribolium confusum |
Tribolium castaneum |
|
|
Жаркий влажный |
0 |
100 |
|
Умеренный влажный |
14 |
86 |
|
Холодный влажный |
71 |
29 |
|
Жаркий сухой |
90 |
10 |
|
Умеренный сухой |
87 |
13 |
|
Холодный сухой |
100 |
0 |
Повторяя опыт при одинаковых условиях, экспериментатор регистрировал то победу одного, то победу другого вида с определенной частотой. Изменение условий выращивания меняло шансы каждого вида на победу, но в достаточно широком диапазоне температуры и влажности с определенной вероятностью победить могли оба вида. Постоянным оставалось только одно: через какое-то время в среде оставался лишь один вид, а второй исчезал.
Отличия между видами одной гильдии (совокупности видов, использующих один и тот же ресурс) могут быть опосредованы влиянием хищника. Так, явное отличие от соседа может вывести вид из сферы внимания питающегося соседом хищника! В Юго-Восточной Азии сетчатые питоны живут прямо в поселениях человека, воруют и едят коз, свиней и собак и крайне редко нападают на человека. Это связано с тем, что человек не похож на своих ходящих на четырех ногах родственников и не воспринимается как жертва.
Внутривидовая конкуренция приводит к расширению ниши, стремящейся расшириться до границ фундаментальной и даже далее (в результате эволюции).
Оценить степень конкуренции между видами можно, предположив наличие некого лимитирующего уровня сходства. Если уровни различий между соседними членами гильдии при сравнимом уровне внутривидовой изменчивости оказываются примерно одинаковыми, это свидетельствует о высокой роли конкуренции в дифференциации ниш.Хатчинсон продемонстрировал сходство уровня дивергенции морфологических признаков видов в одной гильдии на примере отношения размеров ротовых частей (тесно связанных с параметрами добычи) у сосуществующих насекомых, птиц и млекопитающих. Выяснилось, что однотипные структуры каждого следующего вида больше, чем у предыдущего в 1,1—1,4 раза, в среднем — в 1,28 раз. Это разница в размерах, обеспечивающая разницу в весе в 2 (2,09) раза. Это правило касается размеров пищедобывательных структур и всего тела у самых разнообразных животных: пауков, жуков-скакунов, ящериц, саламандр, белок, летучих мышей, пустынных грызунов, плодоядных голубей. Выясняется, что при коэффициенте вариации (CV) 5,5 для двух видов с отношением размеров 1,28 перекрывание ниш составит 1–2%. Виды распределяются вдоль градиента ресурса неслучайным образом. Отношение, близкое к 1,3 применимо для описания различий в сериях магнитофонов, станков и велосипедов! Возможно, это соотношение отражает особенности нашего восприятия, вследствие которых инженеры и маркетологи создают классы изделий, «естественно» отличающиеся друг от друга.
Следует учитывать, что, в отличие от станков и велосипедов, представители видов с более крупными размерами тела зачастую проходят в ходе онтогенеза через размерные классы, свойственные меньшим видам. В правиле Хатчинсона не учтены закономерности аллометрического роста.
Сообщество можно представить себе как n-мерное пространство, занимаемое экологическими нишами разных видов. При диффузной конкуренции каждый вид взаимодействует со множеством других, конкурируя с ними по разным факторам среды. Успех или неудача закрепления вида в сообществе в первую очередь зависит от отношения конкуренции и эксплуатации с другими видами. Представьте себе ящик, заполненный надутыми воздушными шариками: каждый из них давит на все остальные. Если популяция какого-то вида (шарик) «проиграет», то есть не сможет себя воспроизводить (лопнет), эффект от ее исчезновения приведет к сдвигу равновесия между всеми остальными популяциями (перемещению остальных шариков).
С другой стороны, пары видов, сильно перекрывающиеся по одному из измерений ниши, могут существенно отличаться по другому, ослабляя тем самым конкуренцию. Можно говорить о дифференциальном перекрывании, при котором диффузно конкурирующие виды делят нишу по разным параметрам. Например, хищники чаще делят ресурсы по времени суток, так как их добычи тоже могут характеризоваться определенной суточной активностью. Для растительноядных животных расделение по времени суток мало свойственно. Пойкилотермные животные более дифференциально используют сутки, чем гомойотермные.
Чем ближе суммарная численность членов гильдии к емкости среды, тем большую роль может играть конкуренция. Можно грубо предположить, что на более высоких трофических уровнях конкуренция острее.
Степень разделения экологических ниш растений в целом ниже, чем животных.
Поскольку внутривидовая конкуренция оказывается сильнее межвидовой, развитие с метаморфозом или экологический половой диморфизм можно рассматривать как меры, уменьшающие внутривидовую конкуренцию.
Дополнительные материалы:
Учебная модель: Конкуренция между видами и разделение экологических ниш