М.А. Шишкин. Закономерности эволюции онтогенеза // Современная палеонтология. Методы, направления, проблемы, практическое приложение: Справочное пособие: В 2-х томах. / Под. ред. В.В. Меннера, В.П. Макридина. – М.: Недра, 1988. – Т. 2. с. 169-209.
М.А. Шишкин
ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ ОНТОГЕНЕЗА
ОБЩИЕ ЗАМЕЧАНИЯ
Рассматривая морфологическую сторону эволюции онтогенеза, наиболее существенную для палеонтолога, необходимо прежде всего иметь в виду взаимосвязь между изменениями в индивидуальном развитии и эволюцией взрослого организма. Именно эта проблема составляет, например, суть всего учения о филэмбриогенезах А. Н. Северцова [926, 927, 928]. Хотя такое внимание к итогу развития иногда считают чем-то вроде пережитка геккелевского мышления [163, 540] или, в лучшем случае, допустимым методическим приемом [1661], на деле оно неизбежно, если нас интересует реальный эволюционный процесс. Только во взятом изолированно онтогенетическом цикле взрослая стадия может казаться наименее значимой, поскольку она полностью обусловлена предыдущим развитием, но сама уже не определяет ничего. В эволюционном же аспекте дело представляется совсем иначе. Взрослая стадия — единственная, на которой в норме (по крайней мере, у Metazoa) лежит функция размножения, создающего генетическое разнообразие, т. е. материал эволюционного процесса. От успеха взрослых организмов в борьбе за существование зависит, какого сорта гаметы послужат для создания следующего поколения популяции и, следовательно, с каким фенотипическим материалом будет в дальнейшем иметь дело отбор. В процессе отбора вместе с неудачными фенотипами элиминируются и реализующие их онтогенезы.
Это означает, что какие бы удачные приспособления ни создавались на промежуточных стадиях развития, они не дают преимуществ, если в итоге онтогенез не обеспечивает осуществления нужного фенотипа. Взрослая стадия как бы диктует свои условия всему онтогенезу. Поэтому столь часто критикуемые слова Э. Геккеля о том, что филогения (понимаемая им как совокупность взрослых стадий) есть причина онтогении, в действительности имеют глубокий смысл, хотя и не совсем тот, который подразумевался автором биогенетического закона.
То, что изменяются все стадии онтогенеза, ни у кого не вызывает сомнений. Вопрос в другом — как соотносятся их изменения? Эволюирует ли взрослая стадия самостоятельно или под действием одновременных морфологических изменении в раннем развитии, или же обоими путями сразу? Наконец, возможно ли в филогенезе обратное воздействие поздних изменений на ход предшествующих стадий — т. е. такое, которое немыслимо в пределах одного онтогенеза?
В ответе на первый вопрос обнаруживается поразительное противоречие в существующих взглядах на роль онтогенеза в эволюции, объединяющих, по сути, две несовместимые концепции. С одной стороны, признается, что нормальное развитие есть зарегулированный процесс, устойчиво направленный к достижению конечного результата и стремящийся свести на нет все возникающие на этом пути уклонения [1192, 1196, 2439, 2441]. Отсюда следует, что эти вариации не должны непосредственно влиять на облик взрослого организма. Эволюция последнего должна быть связана с формообразовательными уклонениями, относящимися к концу развития, где возможности регуляции снижаются. И в то же самое время господствует убеждение, что изменения ранних или промежуточных стадий могут служить причиной немедленного отклонения (девиации) всего хода онтогенеза, радикально меняющего облик взрослой формы у потомков. Обычно этого противоречия даже не замечают. Обе концепции существуют давно и одинаково берут начало в классической морфологии, но лишь первая из них, как будет показано, может быть согласована с требованиями эволюционной теории.
Первооснову для нее создали два эмпирических обобщения К. М. Бэра [1291]. Одно из них касается уменьшения эмбриональной изменчивости в последовательных стадиях (говорящее, по мнению К. М. Бэра, о присутствии высшего целеполагающего контроля, регулирующего развитие). Второе составляет известный закон Бэра, или закон зародышевого сходства. В рамках эволюционной теории, как указывал уже Ч. Дарвин [309], это явление означает, что отбор стремится изменять организмы главным образом в позднем возрасте. В дальнейшем эти представления получили поддержку со стороны механики развития, экспериментально показавшей способность морфогенетических процессов к саморегуляции, в результате чего взрослый организм оказывается более устойчивым, чем способ его осуществления (в онтогенезе или при регенерации). Это обобщение, известное как «правило Ру» [921, 2184], или принцип эквифинальности [1524],. подтвердило высокую консервативность ранних стадий развития, показывающую, что эволюция взрослой организации должна происходить преимущественно путем изменений в позднем онтогенезе [2185].
Однако главным источником таких представлений послужил основной биогенетический закон Э. Геккеля [1687], представляющий собой переложение на эволюционный язык старой идеи о параллелизме индивидуального развития и «лестницы существ». Выполнение этого закона (т. е. краткое повторение филогенеза в онтогенезе) невозможно представить себе иначе, как на основе конечных изменений развития.
Поскольку Э. Геккеля интересовала в онтогенезе не столько его эволюция, сколько получение метода для филогенетических реконструкций, то о причинах такого хода изменений мы не находим у него специальных разъяснений, кроме многочисленных и мало что говорящих «законов» наследственности и приспособления. Тем не менее в биогенетическом законе содержится вполне целостная теория эволюции онтогенеза, подразумевающая два положения: а) взрослая стадия эволюирует посредством прибавления к онтогенезу новых конечных стадий, что обеспечивает рекапитуляцию в онтогенезе облика взрослых предков (палингенез); б) промежуточные стадии имеют собственную адаптивную эволюцию, искажающую онтогенетическую запись преобразований взрослой стадии (ценогенез).
Биогенетический закон обычно связывают с механоламаркистскими взглядами Э. Геккеля [359, 565, 1196], иногда утверждая даже, что он был опровергнут самим открытием менделевских факторов и мутационного процесса [1661, 1892]. Однако в действительности вопрос о природе наследственности не играет решающей роли в оценке закона [1822]. В самом деле, независимо от того, связывается ли появление эволюционных новшеств с поздними соматическими изменениями, передаваемыми каким-то образом через половые клетки (ламаркистская концепция) или же с непосредственным изменением самих этих клеток (концепция наследственных факторов), в любом случае новый цикл развития неизбежно должен начинаться с преобразований зиготы, т. е. отличаться от родительского с самого начала.
Э. Геккель ясно понимал это обстоятельство и отнюдь не видел в нем препятствия для биогенетического закона, указывая, что изменения, возникшие в определенный момент развития, и передаваемые через родительские гаметы, обнаруживают себя впервые у потомков на той же самой стадии («закон одновременного наследования» [1687]). В этом он прямо следовал Ч. Дарвину [310], сформулировавшему принцип «наследования в соответствующем возрасте» и подчеркивавшему, что появление наследственного уклонения в гаметах и его видимое проявление в индивидуальном развитии у потомства суть разные вещи. Поэтому неудивительно, что» признание детерминантов или генов в качестве носителей наследственности не мешало многим исследователям признавать биогенетический закон [1380, 1692, 2472] или, по крайней мере, связь эволюционных изменений с поздними стадиями [503, 709].
Противоположный взгляд на эволюцию онтогенеза, допускающий изменение взрослых организмов путем уклонения на ранних стадиях развития, ведет свое начало от Э. Жоффруа Сент-Илера [2198] и Ф. Мюллера [719]. У них и у современных авторов он чаще всего имеет одно и то же обоснование — сравнение нормальных онтогенезов (или морфогенезов гомологических органов) и отождествление наблюдаемой конечной онтогенетической разницы (зародышевой дивергенции Бэра) с реальным эволюционным процессом. Если, например, морфогенезы гомологичных органов А и Б у двух разных форм совпадают до стадии х, а затем расходятся, то утверждают, что орган Б возник из А путем девиации на стадии х [2141]. Методологическая несостоятельность этой аргументации будет нами показана. Представления о возможности или неизбежности такого пути эволюции никогда не составляли в отличие от геккелевских какой-либо целостной теории. Тем не менее уже к началу XX в. они стали преобладающими.
Одной из причин этого было накопление фактов сравнительной эмбриологии, не согласующихся, по мнению исследователей, с биогенетическим законом [При этом упускалось из виду, что неполнота выполнения закона заранее предусмотрена в нем ссылками на ценогенезы и сокращенность повторения филогении. Таким образом, к закону предъявлялись требования, не вытекающие из его содержания]. Признание эволюции путем ранних уклонений развития устраняло тезис о первичной неизбежности рекапитуляции, правда, ценой того, что само их существование превратилось в загадку [565, 1869]. Другой момент в критике геккелевских взглядов был связан с растущим убеждением эмбриологов, что подлинное объяснение хода индивидуального развития следует искать в изучении его непосредственных причинных факторов, а не в конструировании умозрительных исторических принципов, подменяющих опытное знание. Этот подход и привел к появлению экспериментальной (каузальной) эмбриологии, которая выявила чрезвычайное разнообразие и специфичность структуры зародышевых клеток у различных организмов. Последнее с очевидностью показывало, что зигота является таким же продуктом эволюции, как и взрослый организм, и что начальная стадия развития высшего организма не есть повторение его одноклеточного предка. «Яйцо курицы не более соответствует начальному звену филогенетической цепи, чем сама курица» [1733]. Все это привело к убеждению, что эволюция происходит путем изменения всего онтогенетического цикла снизу доверху в каждом очередном поколении, а не за счет прибавления конечных стадий [1448, 1763, 2259] [Столетием раньше эти же доводы использовались сторонниками идеи преформированного развития в их критике теории параллелизма. Указывалось, что развитие организма не может проходить через стадии низших классов, поскольку он отличается от них своим предначертанием уже в зародышевой клетке [540]]. Онтогения, таким образом, творит филогению, а не повторяет ее [1623]. При этом одни авторы имели в виду только постепенные преобразования, другие — возможность резких уклонений, скачкообразно меняющих взрослую форму.
В рамках этих взглядов исчезала, однако, всякая теоретическая возможность появления параллелей между онто- и филогенезом вопреки, казалось бы, очевидным фактам их существования. Это затруднение пытались обойти с помощью утверждения, что такие параллели имеют чисто морфогенетические, а не исторические причини, т. е. что анцестральная взрослая стадия может удерживаться в онтогенезе потомка лишь постольку, поскольку она составляет необходимую базу для реализации последующих стадий [1623]. Другое объяснение состояло в том, что большинство подобных примеров, используемых биогенетическим законом, в действительности относится не к повторению облика взрослых низших форм в онтогенезе высших, а просто к сохранению общих стадий развития у тех и у других, т. е. к проявлениям зародышевого сходства, на что указывал еще К. М. Бэр [1623, 1767, 1970 и др.]. Этот довод, постоянно повторяемый и позднее, например [1317, 1319], обычно считают решающим опровержением геккелевской модели эволюции. Но, как ни странно, при этом не замечают, что при перестройке онтогенеза каждый раз с начальной стадии сохранение зародышевого сходства становится таким же невероятным, как и появление рекапитуляции!
В целом, однако, позиция позднейших исследователей по отношению к биогенетическому закону выглядит более умеренной и допускает возможности разных путей онтогенеза. Соответственно выдвигаются различные классификации этих путей, или модусов [565, 926, 928, 1317, 1319, 1604, 2141, 2124], в которых геккелевский способ эволюции путем поздних надставок занимает то или иное место — от сравнительно большого, например у А. Н. Северцова, до ничтожно малого у Г. де Бира [1317]. Почему онтогенез меняется в разных случаях по-разному, этими взглядами не объясняется (если не считать обычного утверждения, что ранние изменения — это путь к созданию крупных систематических групп). А. Н. Северцов [927] прямо указывает, что ответ на этот вопрос вообще не входит в его задачу, которая состоит лишь в том, чтобы выяснить, как эволюция может идти. Он признает, что выдвигаемые им представления о филэмбриогенезах не зависят от принятия той или иной эволюционной теории.
Само разнообразие выделяемых модусов нередко приводит их авторов к выводу, что все логически мыслимые пути изменений индивидуального развития действительно реализуются в эволюции [2141]. Это равносильно признанию, что в преобразованиях онтогенеза нет сколько-нибудь общих закономерностей. Такой малоутешительный итог столетнего изучения проблемы после Э. Геккеля, в общем, неудивителен в условиях, когда оно основывалось на сравнении одних лишь нормальных онтогенезов, вне связи с анализом механизмов, реализующих материал эволюции — онтогенетическую изменчивость.
Очередным подтверждением этого служит работа С. Гулда [1661] о соотношении онто- и филогенеза, представляющая собой наиболее обширное исследование на эту тему за последние десятилетия. Методология здесь все та же — выделяются эволюционные модусы и оценивается, насколько они отражают ход филогенеза. Попытка связать их с двумя типами адаптивных стратегий (r и K-отбором) не меняет традиционного характера подхода к проблеме. Эволюция онтогенеза остается суммой изолированных процессов, каждый из которых идет в своих особых условиях. Вопрос об их общих первопричинах по-прежнему остается в стороне.
ОНТОГЕНЕЗ И СТАБИЛИЗИРУЮЩИЙ ОТБОР
Морфологическая эволюция онтогенеза не может быть понята, исходя из одних лишь ее адаптивных результатов, т. е. из простого сопоставления «готовых» типов нормального развития. В ее основе лежит процесс эволюции формообразовательных механизмов, преобразующий типичную взрослую организацию (адаптивную норму [1184]), а вместе с ней и стандартный путь ее осуществления в онтогенезе. Каждое элементарное изменение взрослой нормы связано с отбором одного из ее уклонений, т. е. с сохранением любых индивидуальных вариантов развития, реализующих данное выгодное уклонение. Этот отбор варьирующих индивидуальных циклов по принципу однозначности их итога неминуемо должен перестраивать типичный ход развития в целом (см. "Эволюция как эпигенетический процесс"), и главная задача эволюционной теории состоит в уяснении закономерностей этой морфогенетической перестройки. Морфологическая эволюция онтогенеза является лишь ее внешним выражением.
Основа для решения этой проблемы содержится в теории стабилизирующего отбора, или эпигенетической концепции эволюции (см. "Эволюция как эпигенетический процесс"), согласно которой создание элементарного адаптивного изменения выражается в росте устойчивости онтогенетического осуществления исходной для него вариации.
В свете этих взглядов помехоустойчивость типичного развития, вскрываемая экспериментальной эмбриологией, рассматривается как результат отбора на максимальную защиту нормального формообразования. Механизмы этой защиты (саморегуляции) наиболее хорошо изучены для «регуляционных» онтогенезов (позвоночные, иглокожие и т. д.), где развитие на ранних стадиях идет путем индукционного взаимодействия зачатков, определяющего направление их дальнейшей дифференцировки. Устойчивый ход развития поддерживается за счет широкого раздвигания порогов нормального реагирования зачатков по самым различным параметрам: количеству и активности метаболитов, протяженности периодов компетенции (способности к нормальному взаимодействию) и т. д. Поэтому небольшие колебания в этих показателях, вызываемые генетическими или внешними нарушениями, в достаточно широких пределах забуфериваются и не влияют на дальнейший ход развития. Сильные же воздействия, необратимо нарушающие развитие, ведут либо к гибели, либо к дефинитивным аномалиям, снижающим жизнеспособность.
Такая же устойчивость характерна и для онтогенезов «мозаичного» типа (моллюски, членистоногие и др.), где органогенезы протекают в небольших группах клеток, и регуляция, очевидно, имеет место внутри них. Зачатки здесь в принципе так же мультипотентны, как и в «регуляционных» онтогенезах (например, зачаток глаза насекомого, способен развиваться в антенну или конечность), но нормальная детерминация осуществляется столь надежно, что на первый взгляд кажется единственно возможной. Между двумя типами развития нет четкой грани и в других отношениях: «мозаичные» фазы имеют место в «регуляционных» онтогенезах I и наоборот. В целом, благодаря саморегуляции, нормальный онтогенез представляет собой канализированную последовательность событий (креод, или, буквально, «необходимый путь» [2441]), стремящуюся выправить все помехи на пути к осуществлению стандартной организации.
Рассматриваемая нами проблема соотношения эволюции взрослого организма и его онтогенеза вначале должна быть поставлена несколько иначе — каким образом становление новых взрослых особенностей связано с перестройкой их формообразовательных механизмов?
Согласно эпигенетической концепции [1178, 1180, 1181, 1192, 1196], в главных чертах эта связь сводится к следующему. Элементарное эволюционное изменение создается отбором как модификация прежней, фенотипической нормы (см. рис. 26, 27, см. "Эволюция как эпигенетический процесс"), осуществляемая лишь в тех условиях, на фоне которых она имеет адаптивное преимущество. Это — примитивное (зависимое) формообразование, в котором среда выступает прямо или косвенно (через посредство функции) как детерминирующий фактор развития. По мере того как новый признак приобретает значение оптимальной адаптации для любого варианта нормальных условий, его развитие становится все более независимым от специфических внешних стимулов, благодаря установлению регуляторных взаимосвязей с другими формообразовательными процессами, составляющими нормальный онтогенез. Реализация адаптивного изменения становится, таким образом, неотъемлемой составной частью осуществления фенотипической нормы как целого. В итоге становление новой элементарной адаптации есть процесс замены внешних факторов ее развития внутренними.
Идея о таком ходе эволюционных изменений принадлежит, как известно, неоламаркизму, но лишь теория стабилизирующего отбора впервые дала ей рациональное обоснование. Адаптивная модификация не возникает одноактно, а создается путем отбора среди элементарных индивидуальных реакций (морфозов), представляющих собой видоспецифичные уклонения нормального развития (см. "Эволюция как эпигенетический процесс"). Ее последующее фиксирование — результат селективного преобразования частного варианта нормы в безусловную норму, а не выражение «прямого унаследования». Признаки вообще делятся не на «наследственные» и «приобретенные», а лишь на устойчивые к внешним изменениям и зависимые от них [1194, 1196, 2501]. Унаследование новой особенности не означает ничего иного, кроме стабилизации ее онтогенетического осуществления [1180, 1181].
Прямым экспериментальным подтверждением описанного хода эволюционных преобразований являются опыты К. X. Уоддингтона [2441] по генетической ассимиляции морфозов у дрозофилы, в которых индуцированные внешними воздействиями изменения в ходе отбора превращались в стойкие наследственные признаки (подробнее см. "Эволюция как эпигенетический процесс"). На такой путь создания адаптации указывает также и множество наблюдений сравнительного характера. Прежде всего это хорошо известные факты параллелизма между адаптивными модификациями у отдельных видов или рас и устойчивыми признаками у их ближайших родственников, что многократно отмечалось для растений, позвоночных, насекомых и т. д. [405, 1192, 1196]. Естественно предполагать, что мы наблюдаем здесь как бы последовательные стадии стабилизации этих признаков, тем более, что указанные модификации у первых («пластичных») видов обычно возникают при воздействии условий, свойственных в норме для вторых («стабильных»). Другие подобные доказательства дают эмбриология и палеонтология. Например, у позвоночных в позднем развитии очень велика формообразующая роль функции, в связи с чем еще В. Ру выделял в их онтогенезе дофункциональный (автономный) и функциональный периоды. Сравнение показывает, что граница между ними сдвигается в эволюции за счет нарастания автономности развития у более высокоорганизованных форм. Так, у водных хвостатых амфибий образование легочных альвеол обусловлено дыханием; у более наземных жаб оно индуцируется гормонально и лишь в конечных стадиях дифференцировки зависит от функции; наконец, у амниот оно протекает вполне автономно в раннем развитии [632].
Во многих случаях устойчивого формообразования первичная зависимость развития от внешних факторов самоочевидна. Например, образование мозолей у млекопитающих и бегающих птиц есть обычная адаптивная реакция кожи на трение. Однако в участках, подвергающихся трению постоянно (например, подошвы конечностей), утолщение эпидермиса наступает еще в эмбриональный период [1196]. Стертая поверхность зубов у дюгоня явно возникла первично на основе жевательных движений, однако в онтогенезе она развивается вне связи с жеванием, путем эмбриональной резорбции поверхности зубов [1317]. Еще один пример — развитие вдавленности на внутренней стороне оборотов раковины наутилоидей. Первоначально она возникла как пассивный результат плотного смыкания оборотов, так что у девонских форм ее еще не было на ранних стадиях развития раковины, предшествующих смыканию. Но уже у некоторых карбоновых видов, как и у современных, вогнутость появляется независимо от механического воздействия — на стадии несвернутой раковины [1776].
МОРФОЛОГИЧЕСКАЯ ЭВОЛЮЦИЯ ОНТОГЕНЕЗА
Прогрессивные дефинитивные изменения. Проследим теперь морфологические проявления стабилизационного процесса. Некоторые из них самоочевидны. Если новая элементарная адаптация всегда возникает как ответ на определенный внешний фактор, а позднее (в филогенезе) автономизируется от него в своем развитии, появляясь уже на предфункциональной стадии, то ясно, что речь идет об ускорении (акселерации) развития или, вообще говоря, о гетерохронии.
Другие закономерности удобнее пояснить с помощью схемы (рис. 28). Допустим, что морфогенез органа проходит через стадии А, В к взрослому состоянию С. Отбор на расширение адаптивных возможностей приводит к тому, что в последующих циклах развития взрослая стадия при воздействии фактора среды f реагирует новой приспособительной дифференцировкой D(DI). Тем самым стадия С рекапитулируется здесь в почти ненарушенном виде (C1); изменилась лишь ее реактивная способность в определенных условиях (f). По мере совершенствования дифференцировки D в филогенезе (DII) ее развитие стабилизируется и больше не нуждается во внешней индукции. Роль формативной основы стадии DII целиком берет на себя предыдущая стадия, которая снова должна видоизмениться (С2), чтобы удовлетворить этому новому требованию. Рекапитуляция исходного взрослого состояния С тем самым искажается уже заметнее. Дальнейшая перестройка развития в ходе его интеграции с другими морфогенезами может затронуть уже не только непосредственную базу структуры D (достигшей уровня DIII или DIV), но и более глубокие основы организации, благодаря чему появляются стадии C3 и В1, а затем и С4, В2, А1. Все эти изменения могут иметь на первых порах чисто физиологический характер, но по мере их углубления они получают и морфологическое выражение.
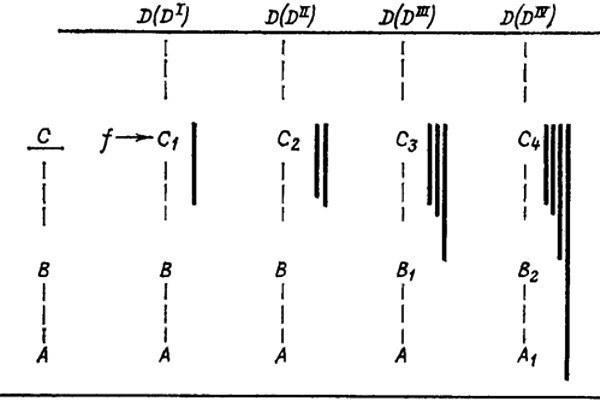
Рис. 28. Перестройка морфогенеза нового признака в процессе его стабилизации. По М. А. Шишкину.
A, A1; В, B1; В2; С, C1–C4 — стадии онтогенетического развития органа; D(DI)–D(DIV) — этапы эволюционной стабилизации нового признака; f—внешний формативный фактор; сплошные вертикальные линии — накопление стабилизационных изменений
Из этой картины ясно, что эволюция поздних стадий не является ни их наращиванием (анаболией), ни замещением (субституцией) в чистом виде, а результатом непрерывного превращения первого процесса во второй. Данный пример, по существу, демонстрирует этапы эволюции онтогенеза при создании единственной надставки D. Поскольку новый признак возникает как формообразовательная реакция прежней взрослой стадии, то он вначале является надстройкой над ней. Но параллельно созданию признака стабилизируется и способ его разбития, что ведет к нарастающему искажению рекапитуляции прежней взрослой стадии.
Таким образом, эволюция путем изменения поздних стадий в равной степени является причиной как появления палингенетических рекапитуляции, так и их последующего искажения. Или, иначе говоря, нарушение биогенетического закона обусловлено в конечном счете той же самой причиной, что и его выполнение. Процесс стабилизационной перестройки, осуществляющий это нарушение, распространяется с затуханием в последовательных онтогенезах от поздней (взрослой) стадии к ранним (рис. 28). Чем дальше он заходит, тем глубже новый цикл развития отличается от исходного. Так, если онтогенез А–DII уклоняется от предкового А–С, начиная со стадии В, то в следующем филетическом звене А–DIII момент расхождения с А–С отодвигается к стадии A, а в дальнейшем (А1–DIV) — к еще более ранним стадиям.
Эти закономерности открывают нам смысл закона К. М. Бэра, говорящего об углублении онтогенетической дивергенции между родственными формами по мере отдаления их родства. На первый взгляд при таких соотношениях между онтогенезами предка и потомка второй из них никогда не может быть продолжением первого. Поэтому неудивительно, что, несмотря на все попытки согласовать закон Бэра с законом Геккеля [930, 1896], они чаще всего считаются «фундаментально несовместимыми» [1661, 1767]. Но при этом не замечают, что эти два обобщения имеют разные области применения, определяемые тем, насколько длительную эволюцию претерпел данный онтогенез по отношению к сравниваемому исходному. Если бы этой эволюции не было, то мы наблюдали бы в развитии любого сколь угодно отдаленного потомка лишь приращение к анцестральному циклу, а не отклонение от него. Закон Бэра мог выполняться бы только для филетически дивергентных форм, причем общий ранний отрезок их развития состоял бы из одних палингенезов (рекапитуляции взрослых общих предков) [Последние могли бы затемняться только эмбриональными адаптациями (ценогенезами)]. И лишь стабилизационные процессы, непрерывно совершенствующие и упрощающие в поколениях путь реализации ранее приобретенных новшеств, приводят к тому, что онтогенетическая запись каждого такого приобретения постепенно преобразуется из «продлевающей» в «замещающую». Другими словами, чем более далекие звенья филогенетической цепи мы сравниваем, тем меньше их онтогенетическая разница подчиняется биогенетическому закону и тем больше — закону Бэра.
Это дает ответ на одно из главных упоминавшихся возражений против биогенетического закона, указывающее, что его классические призеры в действительности относятся к проявлениям зародышевого сходства, которые имеют отношение к особенностям взрослой стадии предка лишь в той мере, в какой они сохранялись у него до конца онтогенеза [1317, 1623]. Например, присутствие у эмбрионов высших позвоночных жаберных щелей повторяет у них в первую очередь эмбриональное, а не взрослое состояние рыбьих предков, и т. д. Неслучайно такие возражения исходят прежде всего от зоологов и эмбриологов, поскольку в качестве моделей предка и потомка им приходится использовать почти исключительно современные формы, представляющие собой продукты дивергентной эволюции с глубоко разошедшимися онтогенезами. Напротив, палеонтологу часто удается наблюдать очень близкие звенья филогенетической цепи и убеждаться, что в этом случае анцестральная взрослая стадия может отражаться в позднем онтогенезе потомка с максимальной полнотой [887]. Причина этого понятна и заключается в том, что морфогенез исторически недавнего приобретения еще мало затронут стабилизационными преобразованиями и в основном сохраняет свой первоначальный вид надстройки над предыдущим взрослым состоянием. Именно этими возможностями, предоставляемыми ископаемым материалом, и объясняется тот странный для многих факт, что, несмотря на многолетнюю уничтожающую критику биогенетического закона, палеонтология по-прежнему остается его оплотом [Иногда даже считают, что теория рекапитуляции имела больше воздействия на палеонтологическую мысль, чем любая другая концепция, за исключением самой идеи органической эволюции [1535]].
Рассмотренные нами закономерности стабилизационных изменений по отдельности не составляют чего-то ранее неизвестного. Мысль о преобразовании надставок развития в субституцию высказывалась С. Г. Крыжановским [540], подчеркивавшим, что прибавление новой стадии в морфологическом смысле означает вытеснение ею предыдущей во временном отношении. Обусловленность наиболее характерных проявлений перестройки онтогенеза (акселерации и спрямления пути развития) состоянием взрослой стадии понималась многими исследователями независимо от того, связывали ли они эти изменения с появлением новых надставок, «впрессовывающих» вглубь прежние стадии (Э. Д. Коп, А. Хайетт, А. Вейсман и др.), или же с самим фактом длительного исторического существования данной взрослой организации (Ф. Мюллер, Ф. Бальфур). В последнем случае интуитивное объяснение, по существу, было очень близко к пониманию процесса стабилизации, так как указывалось, что природа стремится достигнуть желательного конца кратчайшим путем, вызывая укорочение первоначальной онтогенетической записи [1299]. В целом, однако, для гетерохроний (изменения темпов развития) и выпадения стадий выдвигались самые различные объяснительные причины — чисто адаптивные [309, 1775, 2506], механические [2470] или же имеющие особую природу [1457].
Но если указанные явления, с одной стороны, обусловлены эволюцией взрослой стадии, а с другой — сами обусловливают нарушение рекапитуляции, то, очевидно, между двумя последними процессами должна существовать прямая связь. Первым это понял эмбриолог Э. Менерт [1945], установивший, что скорость развития отдельных органов в онтогенезе зависит от степени их филетиче-ского прогресса. Поскольку эта зависимость при неравномерной эволюции органов ведет к нарушению прежних целостных онтогенетических стадий, то изменения, связанные с гетерохрониями, согласно Э. Менерту, не могут считаться фальсификациями исторического развития, а составляют его закономерный результат. В палеонтологии подобные взгляды были почти одновременно высказаны Дж. Смитом [2295].
Наиболее далеко в вопросе о влиянии изменений взрослой стадии на ход онтогенеза продвинулся А. Н. Северцов [927, 928] в раннем варианте своей теории филэмбриогенезов. Анализируя факторы нарушения рекапитуляции, не связанные с провизорными приспособлениями, он выделил в них две группы — первичные и вторичные филэмбриогенезы. Под первыми подразумевались ранние эмбриональные уклонения, преобразующие ход дальнейшего онтогенеза, под вторыми — искажения онтогенетической записи, обусловленные эволюцией конечных надставок, т. е. те самые изменения, которые интерпретированы нами как результат стабилизационной перестройки.
Основываясь на правиле Э. Менерта, А. Н. Северцов показал, что следствием поздних изменений органов является искажение рекапитуляции даже в пределах отдельных органогенезов. Сдвигаясь на другие стадии, закладки эволюирующих органов меняются и сами по различным параметрам (размеры, гистологическая структура, форма, соотношения с другими частями). Распространение этих изменений в направлении ранних стадий (в филогенезе) было продемонстрировано А. Н. Северцовым как для ускорения, так и для замедления развития. Был сделан также вывод о наибольшей полноте рекапитуляции в самых поздних стадиях, еще мало задетых вторичными изменениями. Наконец, было показано, что ускорение (эмбрионализация) развития взрослых черт связано не с позднейшей надстройкой онтогенеза (как гласит обычная интерпретация биогенетического закона), а с длительностью их собственного исторического существования, как полагал еще Ф. Мюллер. Отсюда следовало (хотя и не было четко сформулировано), что эмбрионализация сама по себе есть признак прогрессивной эволюции органов.
Таким образом, А. Н. Северцову уже были ясны основные морфологические закономерности, рассмотренные нами. Отсутствовало лишь представление о факторе эволюции, приводящем их в действие, — стабилизирующем отборе, ведущем к максимальна надежному воспроизведению адаптивного фенотипа в онтогенезе. Вместо этого выдвигались традиционные объяснения, предполагающие компенсацию удлинения онтогенеза и узко адаптивное значение сдвигания стадий для взрослой формы. Поскольку изложенные представления целиком опирались на эмпирическое правило Э. Менерта, понимаемое как принцип «сквозного» изменения темпов развития, то, как только обнаружились случаи, где это правило выполняется не столь жестко (время закладки органа у двух форм может оказаться одинаковым, несмотря на различия в степени его филетического развития), поколебалась и вся основа этих взглядов.
В результате А. Н. Северцов [926, 930] отказался от понятия о вторичных филэмбриогенезах. В его схеме, правда, появились «вторичные архаллаксисы», означающие частный случай того же явления, но они уже не рассматривались как закономерный результат эволюции поздних стадий. В такой же частный модус превратились и гетерохронии. Вместо перестройки, вызываемой надставками, филэмбриогенезами стали называться сами надставки (модус анаболии). Единый процесс распался на изолированные явления. В итоге из двух логически допускавшихся способов перестройки онтогенеза недоказанный путь (ранние изменения) остался в теории А. Н. Северцова, а бесспорный выпал. Теория превратилась в модификацию биогенетического закона, в которой, как и у Э. Геккеля, нарушение рекапитуляции не обусловлено эволюцией взрослой стадии. Разница лишь в том, что в геккелевском варианте все нарушения закона названы ценогенезами, а в северцовском к ним добавлены архаллаксисы и девиации [Насколько нам известно, никто, за исключением отчасти В. Е. Руженцева [887], не обратил внимания на это радикальное изменение содержания теории филэмбриогенезов]. Эта эволюция взглядов А. Н. Северцова лишний раз показывает, насколько судьба правильных эмпирических обобщений может зависеть от их теоретического обоснования.
Регрессивные дефинитивные изменения. Стабилизационные изменения, сопровождающие формирование новых адаптации, связаны с одновременным распадом старой адаптивной нормы. Оба эти явления составляют две неразрывных стороны эволюционного процесса. В любой морфологической структуре прогрессивное развитие одних черт сопровождается деградацией других (например, превращение плавника кистеперой рыбы в пятипалую конечность — это одновременно утрата кожных плавниковых лучей, развитие копыта лошади — это также редукция боковых пальцев и т. д.). Каждый элементарный шаг в этих изменениях имеет одну и ту же основу — отбор на устойчивость преобразованной нормы, ведущий к перестройке ее генотипа как целого, т. е. к изменению пространства индивидуальных геномных вариаций, допускающих ее надежное осуществление ([1180]; см. "Эволюция как эпигенетический процесс").
По отношению к любому признаку, потерявшему свое значение и не поддерживаемому более отбором, это изменение пространства вариаций выглядит как накопление беспорядочных генетических изменений, все более расшатывающих его развитие. Оно выражается в постепенном сдвигании различных морфогенетических параметров к тем порогам, за которыми уже невозможна регуляция нормального развития [1196, рис. 11], в результате чего последнее становится все более изменчивым и неполным. Вначале такие сдвиги задевают лишь позднее развитие; но по мере их накопления у потомков они начинают нарушать и более эффективно регулируемые ранние стадии.
Таким образом, при разрушении формообразовательного процесса обнаруживаются те же характерные черты стабилизационной перестройки, что и при его созидании. В обоих случаях изменения распространяются с затуханием в сторону все более ранних стадий и постепенно автономизируются от влияния факторов среды. Если сначала прежний ход развития преимущественно лишь теряет устойчивость, осуществляясь во все более узком интервале внешних условий, то в последующих поколениях он нарушается уже независимо от их колебаний. И хотя высокая изменчивость рудиментарных органов всегда сохраняется [310, 2141], она приобретает все более неупорядоченный характер. Среди примеров незавершенной рудиментации можно назвать развитие глаз и пигментации кожи у пещерной амфибии Proteus, где утраченная норма все еще в большей мере восстанавливается при выращивании личинок на свету. Напротив, стойкая утрата способности к метаморфозу, характерная в той или иной степени для многих хвостатых амфибий, есть проявление более глубоко стабилизированного недоразвития.
Путь углубления редукционного процесса хорошо виден из сопоставления типов рудиментации конечностей у современных рептилий, изученных А. Н. Северцовым [931]. Сравнение развития геккона (Ascalobotes) и змеевидных ящериц (Seps, Ophisaurus) позволяет считать, что в филогенезе редукция начинается с конечных стадий нормального морфогенеза, а затем захватывает все более ранние стадии, причем поздние к этому времени могут уже совсем выпадать. Так, при относительно меньшей степени редукции (Seps) не возникают фаланги и метаподий 1-го пальца, а также концевые фаланги остальных пальцев (нормально возникающие последними), тогда как проксимальные (рано развивающиеся) элементы закладываются уменьшенными. Дальнейшая рудиментация (Seps, Ophisaurus) ведет к исчезновению почти всех элементов кисти и уже сильному уменьшению зачатка zeugopodium. При этом у обеих форм процесс редукции уже настолько сокращен и ускорен, что целиком осуществляется эмбрионально [А. Н. Северцов [931] называет рудиментацию отрицательным архаллаксисом, поскольку весь процесс здесь выглядит эмбриональным, и в онтогенезе зачатки дегенерирующих органов сразу возникают уменьшенными. Однако вторичность обоих явлений ясно вытекает из закономерностей, вскрытых в этом случае самим же Северцовым. Им показано, что зачаток, уменьшенный как будто бы «сразу», на самом деле становится таковым в филогенезе постепенно («делается все меньше и меньше» [931, с. 514]) и что рудиментация в ходе эволюции распространяется с поздних стадий на ранние. Если на основании этих двух положений экстраполировать к начальному этапу рудиментации, то неизбежен вывод, что ранние зачатки вообще ею не затрагивались, и изменения проявлялись лишь на самых поздних (постэмбриональных) стадиях. Таким образом, не орган уменьшился вследствие уменьшения его зачатка, а наоборот, зачаток стал маленьким вследствие филогенетического углубления редукции органа].
Наиболее характерные проявления недоразвития — выпадение анцестральных конечных стадий и замедление предшествующих преобразований, ведущие к переходу прежних эмбриональных или ювенильных черт во взрослое состояние — описаны под множеством названий; ретардация (Э. Д. Коп, А. Хайетт), брадигенез (А. Грабау), неотения (Ю. Колльман), эпистаз (О. Иекель), фетализация (Л. Больк), педоморфоз (У. Гарстанг), аббревиация (В. Франц), отрицательная анаболия (А. Н. Северцов), протерогенез (О. Шиндевольф) и т. д. На первый взгляд это — процесс, противоположный прогрессивному развитию, но, как мы видели, его эволюция подчиняется тем же самым закономерностям. Он точно так же начинается всегда с изменения (в данном случае — распада) дефинитивного состояния и по мере его усиления распространяется у потомков на все более ранние фазы развития. Стирание рекапитуляции полно развитого состояния органа происходит при его регрессе точно таким же путем, что и в случае его дальнейшего усложнения или специализации. Другими словами, нет принципиальной разницы, затемняется ли в онтогенезе потомков стадия пятипалой когтистой лапы благодаря тотальному недоразвитию конечности (угревидные хвостатые амфибии и ящерицы) или же вследствие ее преобразования в крыло птицы.
Непосредственные адаптивные изменения промежуточных стадий. Все сказанное об эволюции взрослой стадии в равной степени приложимо и к любым другим стадиям. Каждая фаза онтогенеза может изменяться путем отбора ее регулируемых вариаций сколь угодно далеко при условии, что это не нарушает нормального течения последующего развития. Эти регулируемые, т. е. сходящие затем на нет провизорные приспособления, начинаются, подобно дефинитивным, с конечных (по времени) изменений той стадии, на которую направлен отбор; затем их механизм стабилизируется, захватывая и предыдущий отрезок развития. Сравнительно-эмбриологические доказательства поздних изменений провизорных стадий приведены, например, для яиц рыб и личинок амфибий [540, 932]. Стабилизационное ускорение таких изменений прослежено в развитии рисунка окраски у гусениц бражников, в котором более простые предковые типы рекапитулируются и сдвигаются на ранние стадии [2470].
Другой пример — развитие головастиков бесхвостых амфибий. Среди них существуют четыре типа строения жаберных камер, причем только в одном из них имеется парное spiraculum, (рис. 29, A1–Г1). Этот тип почти наверняка является исходным для остальных, которые, очевидно, когда-то возникли у вполне «взрослых» головастиков путем слияния обоих отверстий или зарастания одного из них. Парные отверстия действительно рекапитулируются во всех типах и далее преобразуются обоими указанными путями. Однако у современных форм это происходит на такой ранней стадии, когда опекулярные складки (стенки камер) только начинают развиваться, а самих внутренних жабр, характерных для «взрослого» головастика, еще нет (рис. 29, А–Г ср. [2319]). Очевидно, что ход первично позднего преобразования здесь давно изменен и ускорен в процессе его стабилизации. В основе последней лежит перестройка морфогенетических механизмов, следы которой иногда вполне явственны. Так, у наиболее специализированных головастиков передняя лапка развивается в жаберной камере и выходит наружу, прорывая operculum. Вероятно, вначале этот прорыв осуществлялся за счет механического давления лапки; но у современных форм перфорация operculum — вполне автономный процесс, вызываемый продуктами распада внутренних жабр [2014].
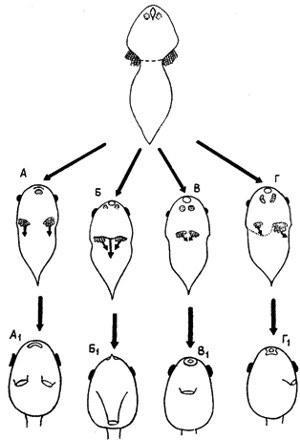
Рис. 29. Типы онтогенетического развития spiraculum у личинок бесхвостых амфибий. По П. X. Старетт.
А, Б, В, Г — ранние стадии, показывающие рекапитуляцию парного spiraculum; A1, Б1, В1, Г1 — стадии «взрослых» головастиков (A–A1 — Pipidae и Rhinophrynidae; Б–Б1 — Microhylidae; B–B1 — Ascaphidae и Discoglossidae; Г–Г1 — остальные бесхвостые)
Таким образом, непосредственная адаптивная эволюция любой стадии онтогенеза протекает по одним и тем же законам, т. е. начинается с ее конечной фазы и затем, по мере стабилизации достигнутого изменения, распространяется вглубь в последующих поколениях. Тем самым исторические преобразования любой отдельной стадии сами по себе могут влиять лишь на предыдущий, но не на последующий ход онтогенеза. Эта значительная самостоятельность эволюции стадий приводит к тому, что чем больше нарастают изменения любой из них, тем более глубокой должна быть перестройка при переходе к следующему отрезку развития, имеющему собственную адаптивную эволюцию. Именно в этом состоит причина резких превращений, свойственных развитию с выраженным метаморфозом, отражающему перестройку между существенно различными возрастными адаптивными типами (у ракообразных, насекомых с полным превращением, бесхвостых амфибий и т. д.). Благодаря такому механизму преобразований онтогенеза его течение характеризуется не просто уменьшением изменчивости к концу, как подчеркивал К. М. Бэр, а чередованием стабильных, адаптивно значимых фаз, или «установок» [415], и более вариабельных отрезков, подготавливающих эти фазы. К числу первых относятся, например, различные специфические личиночные типы. В развитии черепа хвостатых амфибий такими адаптивно важными «установками» являются моменты прорыва рта и перехода к активному питанию, когда основные морфологические изменения, в отличие от предшествующих стадий, оказываются строго скоординированными [566]. Общим итогом развития всегда остается стабильное осуществление взрослой адаптивной нормы. Иными словами, главный принцип эволюции онтогенеза состоит в том, чтобы обеспечить это осуществление вне зависимости от самостоятельных адаптивных изменений его промежуточных стадий.
Провизорные адаптивные изменения вместе с их стабилизационными последствиями составляют то, что может быть названо ценогенезами. Это понятие имеет разные толкования вследствие его двойственности у Э. Геккеля [1688], который отождествил в нем: а) любые нарушения палингенезов и б) приспособления ранних стадий. Для авторов, отрицающих палингенезы, ценогенез как сопряженное понятие вообще не имеет содержания [540, 1623] при использовании только второго из названных критериев [414, 928] оно охватывает и типичные палингенезы (например, жабры у личинок амфибий); если же исходить из первого [1945], то ценогенезом окажется любое стабилизационное изменение типа гетерохроний, несводимость которых к собственно провизорным адаптациям давно отмечена [926, 565, 928, 1317]. Неопределенность употребления термина иногда заходит так далеко, что им обозначают просто любой новый признак [2078]. Введение понятия о стабилизационной перестройке впервые позволяет внести ясность в этот вопрос. Дело в том что ценогенез в представлениях Геккеля был прежде всего равнозначен изменениям, не связанным с эволюцией взрослого организма. Но на самом деле часть нарушений рекапитуляции обусловлена именно последствиями этой эволюции (стабилизацией дефинитивных изменений). Тогда все, что остается за их вычетом и должно составлять ценогенез в идеальном понимании. Сюда должны включаться собственно адаптивные изменения не только палингенетических признаков [358], но и любых других, таким образом, ценогенезы — это любые преобразования, не связанные с эволюцией взрослой стадии и совершенствованием способа ее осуществления.
«ЭМБРИОНАЛЬНЫЕ МОДУСЫ» И ОНТОГЕНЕТИЧЕСКАЯ ЗАПИСЬ
Рассмотрим теперь вопрос о реальности эволюции взрослой стадии путем одновременных изменений в раннем онтогенезе (архаллаксисов и девиаций). Как уже было сказано выше, это представление несовместимо с фактами, говорящими о целенаправленном (канализированном) характере онтогенетического процесса, основанном на регуляции промежуточных вариаций. Среди доводов, выдвигаемых в пользу возможности эволюции путем онтогенетических скачков особое место занимают попытки найти ее следы непосредственно в самих онтогенезах. В этой связи нам придется сначала затронуть проблему исторического истолкования стадии развития в свете представлений о стабилизационных изменениях онтогенеза. Для этого проследим закономерности стабилизации на конкретном схематизированном примере — срастании костей голени в эволюции бесхвостых амфибий (рис. 30).
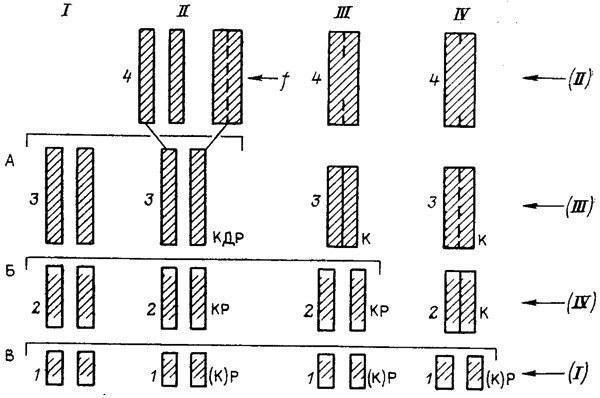
Рис. 30. Эволюция слияния элементов голени в истории бесхвостых амфибий. Составил М. А. Шишкин, оригинал.
I — исходное состояние (пермские рахитомы); II — предки бесхвостых, близкие к раннетриасовому Triadobatrachus; III — промежуточный этап; IV — современные бесхвостые; 1–4 — стадии морфогенезов; А, Б, В — уровни зародышевого сходства между исходным типом развития и морфогенезами II–IV; (I)–(IV) — исторический порядок возникновения отдельных стадий морфогенеза IV; К (полное окостенение), Д (полное удлинение), Р (раздельность элементов голени) — рекапитулируемые признаки взрослого предка; f — внешний фактор; штриховкой показана степень окостенения
Это срастание, как и многие другие скелетные изменения, было связано у бесхвостых с передвижением прыжками, возникшим, по-видимому, как приспособление к мгновенному укрытию в воде от прибрежных хищников [1620]. У древнейших раннетриасовых Salientia (Triadobatrachus), как и у их рахитомных предков, соединения костей голени еще не было или же оно в какой-то мере проявлялось в позднем возрасте [Последнее возможно в том случае, если единственный имеющийся образец Triadobatrachus в действительности представляет собой личинку в стадии позднего метаморфоза [1674]. У современных бесхвостых кости голени на этой стадии уже частично слиты] (см. рис. 30, I, II). Облегчая прыжки, оно должно было в то же время затруднять нормальное четвероногое передвижение, уменьшая подвижность голени. Первоначально оба способа локомоции, очевидно, сосуществовали у древних Salientia, причем степень фиксации взаимоположения костей голени у взрослых животных могла в большой степени определяться интенсивностью функции прыгания, игравшей роль индуктора (см. рис. 30, II). По мере того как новая форма поведения становилась основной, отбор вел к ускоренному соединению, а затем и слиянию берцовых костей, так что оно начиналось уже у самых юных сформированных особей и притом все более автономно (см. рис. 30, III). Следующая эволюционная стадия (см. рис. 30, IV) типична для современных лягушек (Rana), у которых начало слияния приурочено к раннему метаморфозу [1562, 1826].
Этот схематизированный эволюционный ряд, представляющий собой пример гетерохронии, позволяет проиллюстрировать рассмотренные ранее закономерности. Мы видим, что новый признак может почти не меняться в филогенезе (см. рис. 30, II-4–IV-4), но его формообразовательный механизм преобразуется. Эти изменения захватывают все более ранние стадии: если сначала объединение голени, по-видимому, индуцировалось поведением взрослого животного, то у поздних форм оно детерминируется уже на стадии головастиков. Рекапитуляция исходной «двухкостной» взрослой голени, будучи практически полной вначале, у потомков все более обедняется. Если у формы II на стадии 3, предшествующей слиянию, рекапитуляция охватывает три учитываемых признака — окостенение (К), взрослое удлинение (Д) и раздельность составляющих элементов (Р), то в морфогенезе потомков эта стадия выпадает, а ее признаки все более теряют взаимосвязь, распределяясь по другим стадиям. Стабилизация ведет, таким образом, к стиранию палингенеза. Общей для всех форм остается лишь ранняя стадия с ясно выраженным признаком Р, выступающая как элемент зародышевого сходства. В целом видно, что рекапитуляции на поздних стадиях наиболее полны, но исторически недолговечны; напротив, на ранних они максимально обеднены, но устойчивы.
Этот ряд позволяет еще раз показать обусловленность закона К. М. Бэра стабилизационной перестройкой, распространяющейся у потомков «в глубь» онтогенеза. В нашем примере действительно видно (см. рис. 30, А–В), что при сравнении онтогенеза I со все более преобразованными (акселерированными) типами II–IV уровень дивергенции между ними смещается на все более раннюю стадию. И если у формы II конец онтогенеза еще представляет собой надставку над взрослой стадией (3) предка I, то уже у формы III поздний онтогенез (стадии 3, 4) выглядит как замещение этой предковой стадии. Как уже отмечалось нами, это постепенное преобразование онтогенетической записи из продлевающей в дивергентную и есть причина того, что биогенетический закон по отношению к каждому анцестральному состоянию выполняется достаточно полно лишь у ближайших потомков.
Данный пример демонстрирует еще одно важное следствие стабилизационного процесса: несоответствие между порядком появления онтогенетических стадий в филогенезе и онтогенезе [1176, 1178]. В наиболее измененном типе развития IV порядок стадий 2–4 прямо противоположен историческому. Взрослая стадия 4 в основных чертах унаследована здесь от далекого предка (II), тогда как стадия 3 появилась в филогенезе позднее (III), а наиболее ранняя — стадия 2 — только что возникла (IV). И если онтогенетически стадия 4 является следствием стадии 2, то исторически, наоборот, стадия 2 является следствием стабилизации развития стадии 4. И лишь в раннем развитии (тип IV, стадии 1, 2) обе последовательности в нашем примере совпадают.
Подобное несоответствие, в общем, не было новостью уже для Э. Геккеля, который фактически признал его, введя понятие ценогенеза. Об этом же говорит его апелляция к методу тройного параллелизма как средству проверки филогенетических реконструкций, а также его признание, что эти реконструкции всегда гипотетичны и во многом «опираются на интуицию» [цит. по 2079]. На невозможность прямого прочтения онтогенеза как эволюционной последовательности указывалось потом множество раз, главным образом в контексте критики биогенетического закона, причем нередко признают, что единственный способ достоверного истолкования онтогенеза — это знание предшествующего филогенеза [540, 1661, 2079, 2141, 2284]. Причину этого, однако, видят не в стабилизационном процессе, а в том, что эволюция онтогенеза может идти разными путями, но каким именно в каждом конкретном случае — мы не знаем. В дальнейшем будет показано, что это представление в основном есть просто следствие произвольной расшифровки результатов стабилизационного процесса. Именно воздействие последнего (вместе с ценогенезами) и приводит к тому, что в итоге «развитие организма не несет в самом себе метода для определения исторической значимости стадий развития» [540].
Этот вывод имеет решающее значение для оценки существующих представлений об эволюции онтогенеза. Мы можем убедиться, что мнение о возможности эволюции взрослой стадии посредством единовременных эмбриональных изменений, бытующее уже полтора века, в действительности почти целиком основано на примерах прямого исторического прочтения онтогенезов, т. е. на том же самом принципе, что и биогенетический закон, но доведенном до абсолюта. А именно: если два онтогенеза дивергируют от некоторой общей ранней стадии, то заключают, что одна из сравниваемых форм возникла из другой путем девиации на этой общей стадии [926, 2141]. В нашем примере (см. рис. 30) это означает, что из прямого сопоставления морфогенезов I и IV, дивергирующих на стадии начального окостенения разделенных элементов голени (1), мы должны были бы заключить, что слияние голени в эволюции лягушек произошло скачкообразно путем объединения ее зачатков у головастиков! Возможность того, что взрослый признак может быть древнее, чем его эмбриональная стадия, обычно не принимается во внимание, если нет прямых доказательств этого; уклонение онтогенеза потомка отождествляется с историей возникновения этого потомка.
Возникает парадоксальное противоречие, которого не замечают. С одной стороны, история первого появления нового взрослого признака есть свершившийся факт, который меняться не может; с другой стороны, разница между первичной и преобразованной онтогенетической записью этого события нарастает в эволюции непрерывно, по мере стабилизации (эмбрионализации) признака у все более отдаленных потомков. Например, на рис. 30 истинная история слияния голени у лягушек иллюстрируется филогенетическими этапами I и II, но она будет выглядеть каждый раз по-новому при сравнении состояния I с этапами II, III и IV, если считать, что разница исходного и итогового онтогенеза показывает нам в каждом случае «модус» возникновения признака. Указанное противоречие не возникает лишь в том случае, если морфогенез раз возникшего признака никогда более не менялся бы.
Самое удивительное, однако, то, что реконструирование онтофилогенетических событий указанным способом не мешает исследователям признавать гетерохронии, выпадения стадий и другие вторичные изменения морфогенезов, искажающие подлинную картину возникновения новых взрослых структур. Другими словами, вполне однотипные явления онтогенетической дивергенции между предков и потомком могут истолковываться в одних случаях как доказательство происхождения потомка путем девиации, а в других, если имеются переходные онтогенезы, — как результат эмбрионализации изменений, первично возникших в позднем развитии.
В нашем примере (см. рис. 30) это означает, что один и тот же онтогенез IV может быть расценен двояко: либо как результат ускорения развития взрослой стадии 4, возникшей в онтогенезе II, — если мы знаем всю последовательность I–IV; либо как изменение онтогенеза I путем девиации на стадии 1, если известны только I и IV. При этом выбор интерпретации не имеет никакой иной основы кроме наличия или отсутствия фактического материала по промежуточным историческим этапам. Этот дуализм методологии типичен для исследований, классифицирующих эволюцию онтогенеза по модусам. Например, у А. Н. Северцова в одной и той же работе можно найти аргументы для обоих взаимоисключающих способов интерпретации онтогенеза. С одной стороны, говорится, что древний взрослый признак со временем передвигается на ранние стадии онтогенеза [928, с. 141], с другой же, утверждается, что «если признак появляется сразу в зачатке данного органа... и сохраняется... до взрослого состояния, то мы можем с большой вероятностью сказать, что этот признак филогенетически возник путем эмбрионального изменения» [928, с. 129].
Возникает вопрос — на чем же тогда следует основывать конкретный выбор между этими двумя правилами? Другими словами, как гарантировать, что между сравниваемыми типами развития не существовала в каждом случае цепь онтогенезов, показывающих первично взрослое происхождение нашего «эмбрионально возникшего» признака? Этот вопрос, однако, не ставится, и в большинстве приводимых примеров, в частности в случае смещения вперед брюшных плавников у костистых рыб [930], мы находим один и тот же аргумент: если бы строение потомка возникло путем надставки (анаболии), то мы бы видели у него рекапитуляции предшествующих взрослых состояний; а раз этого нет, то имело место эмбриональное изменение. Этот довод означает на самом деле молчаливое допущение, что никакой перестройки онтогенеза при его длительном конечном наращивании не происходит — ибо только в этом случае можно было бы ожидать полного отражения всего ряда предков в онтогенезе потомков. И, однако, наряду с этим признаются «вторичные архаллаксисы» [926], или изменения, основанные на вторичных искажениях записи надставок, т. е. как раз на том явлении, которое неявно отрицается при доказательстве «истинных» эмбриональных модусов. Эта произвольность интерпретации еще больше бросается в глаза, когда она оказывается разной для вполне однотипных явлений. Так, утрата в развитии конечности лошади пятипалой стадии расценивается как вторичное искажение анаболии, а аналогичная утрата закладок боковых пальцев у змеевидных ящериц — как истинный архаллаксис [ср. 928, с. 158; 930, с. 428].
Обоснование «эмбриональных модусов» у других авторов строятся на той же самой предпосылке — представлении о неизменности предшествующей онтогенетической записи при эволюции путем надставок. Например, А. Ремане [2141], также предлагающий систему модусов, на первый взгляд признает, что разница между двумя онтогенезами (или морфогенезами гомологичных органов) не может служить прямым указанием на ход филогении. Однако в противоречии с этим он тут же пытается доказать существование девиаций именно из сопоставления двух онтогенезов. Рассуждения А. Ремане в схематической форме повторяют ход рассуждений А. Н. Северцова [926] при доказательстве им девиации в развитии ноздрей у рыб семейства Scombresocidae и сводятся к следующему. Если онтогенезы двух форм дивергируют от стадии А, то в принципе эта может быть результатом их дивергентной эволюции по способу анаболии от общего предка, у которого взрослый облик соответствовал стадии А, а онтогенез — предшествующим общим стадиям двух форм. Однако подобное расхождение двух онтогенезов часто происходит на столь недифференцированной стадии, что она заведомо не может быть рекапитуляцией взрослого предка. В этом случае следует признать происхождение одной формы из другой путем эмбриональной девиации на стадии А.
Из этих соображений видно, что в представлении А. Ремане анаболия означает полное сохранение стадии взрослого предка (и его онтогенеза) в онтогенезе сколь угодно отдаленного потомка, и нарушение этой картины для него представляется не иначе, чем результатом девиации. Нетрудно видеть, к каким выводам мы можем отсюда придти. Если, например, в развитии крокодила и птицы нет стадии, представляющей их общего архозаврового предка, то мы должны признать, что либо обе формы произошли путем девиации от эмбрионов каких-то третьих форм, либо что птицы произошли от эмбрионов крокодилов. На этом же основании можно вывести какой угодно организм из какого угодно, поскольку здесь требуется лишь чисто негативный критерий — отсутствие общей стадии онтогенеза, соответствующей общему взрослому предку!
Приведенные соображения ясно показывают неприемлемость сравнительно-онтогенетического обоснования «эмбриональных модусов». Видеть в эмбриональном появлении взрослого признака прямое указание на способ его появления в филогенезе — значит судить об эволюции онтогенеза, исходя из отсутствия такой эволюции. Этот способ реконструкции исторических событий предполагает столь буквальное и ненарушимое их отражение в онтогенезе, на какое никогда не претендовал даже биогенетический закон.
Все сказанное относится также к «архаллаксисам» и «девиациям», описанным у ископаемых животных. Методология здесь та же самая — прямое отождествление онтогенетического различия с историческим событием. Если новый взрослый признак проявляется в онтогенезе рано, то нередко заключают, что именно так он возник в филогенезе — без скидок на возможность его вторичной акселерации или, наоборот, ретардации прежнего ювенильного состояния. И это несмотря на то, что проявления гетерохроний наиболее обычны именно на палеонтологическом материале, где они встречаются, по словам В. Е. Руженцева [887], на каждом шагу [Палеонтологам еще со времен А. Хайэтта известно о выпадении ранних стадий при акселерации («вторичные архаллаксисы» Северцова); это явление позднее называли липогенезом, или липопалингенезом [1386, 1863]]. Даже признавая, что предложенные А. Н. Северцовым критерии для распознания первичных и вторичных эмбриональных модусов по сути ничем не различаются, исследователи все же иногда продолжают выделять «истинные» архаллаксисы и девиации [708, с. 127]. Более того, в тех же самых работах, где описывается «скачкообразное появление» новых взрослых признаков в раннем онтогенезе, нередко можно найти примеры, когда эти же признаки у близких групп формируются постепенно, так что вывод об их вторичном сдвигании на ранние стадии как бы напрашивается сам собой. Например, Д. М. Раузер-Черноусова [861] описывает как девиацию раннее (в онтогенезе) появление субцилиндрических камер взрослого типа у фузулиниды Wedekindelia (Foraminifera), но тут же указывает, что типичным для фузулинид является позднее развитие таких камер. Точно так же выглядит описанная этим автором «девиация» (появление крупных камер) у Nodosaria nechaevi на фоне онтогенеза у других видов этого рода.
В. Е. Руженцев [887] и Б. И. Богословский [91] считают девиацией дорсальное смещение сифона у климений (Ammonoidea), несмотря на то что среди ранних аммоноидей (гониатитов) подобное же, хотя и менее устойчивое, смещение проявляется постепенно — либо на поздних оборотах раковины (в подотряде Tornoceratina), либо только на средних, т. е. ценогенетичесет (в подотряде Goniatitina) [343]. Логично предположить, что климениевый способ развития возник либо путем акселерации первого варианта, либо фетализации второго. Эти допущения невозможно устранить на том лишь основании, что между гониатитами и климениями, вероятно, нет прямой связи [91]. Речь здесь идет о типичном способе эволюции данного морфогенеза, который мог осуществляться параллельно в разных филетических рядах.
И все же палеонтологи, начиная с А. Хайэтта и Л. Вюртенбергера, в большинстве своем признают, что основным способом эволюции является изменение (надставка или выпадение) поздних стадий развития [291]. Это. мнение преобладает и в современных работах, например [306, 343, 627, 703, 861, 887, 965]. Из них видно, что все преобразования онтогенезов, реально наблюдаемые у ископаемых форм (помимо смены конечных стадий), — это в действительности либо провизорные ценогенетические изменения, либо различные гетерохронии и их последствия. «Девиации» палеонтологов — это конечные результаты гетерохроний, скрытых от нас неполнотой ископаемой документации.
Из сказанного понятно, что, говоря о роли раннеонтогенетических изменений в эволюции взрослого организма, следует различать две стороны проблемы: 1) вероятность их прямого воздействия на взрослую стадию в пределах одного онтогенетического цикла (т. е. вопрос о реальности девиаций развития) и 2) возможность постепенного перехода таких изменений во взрослое состояние в филогенезе по мере замедления развития и выпадения его конечных стадий у потомков (фетализация). Первая из этих возможностей, как мы видели, не согласуется с фактами, говорящими о канализированности (забуференности) развития, и принципиально не доказуема путем сопоставления дивергентных онтогенезов. Напротив, фетализация составляет хорошо известное явление, неразрывно связанное с прогрессивной эволюцией и подчиняющееся тем же самым закономерностям, т. е. распространяющееся в филогенезе от поздних стадий развития к ранним.
Принципиальная разница между «всплыванием» ранних стадий при фетализации и гипотетическими «эмбриональными модусами» часто не осознается исследователями, и первый процесс либо объединяют под общим названием с девиацией (педоморфозов понимании Г.де Вира) [1317], либо считают разновидностью последней—«девиация с запаздыванием» у фораминифер [261, 667, 629], либо отождествляют с архаллаксисом [527, 887, 1984]. Главная причина этого, по-видимому, в том, что при недоразвитии материалом для изменений взрослого состояния служат уже существующие провизорные морфологические особенности и их «всплывание» нередко понимают как скачкообразную замену одного фенотипа другим [1185, 2420]. В действительности это не так. Реализация ювенильного признака в онтогенезе и его дефинитивное проявление в ходе эволюции — это совершенно разные события, из которых первое совершается во всех индивидуальных циклах по заданной программе, а второе — достигается в поколениях шаг за шагом. Всякое адаптивное изменение взрослого фенотипа связано с последовательным отбором и стабилизацией мелких онтогенетических вариаций, направленных в сторону создаваемого изменения. Это касается и замедления развития, что в итоге и обусловливает постепенность утраты прежней взрослой стадии. Реальность такой изменчивости, составляющей потенциальный материал для регрессивных преобразований, показана, например, для келловейского аммонита Kepplerites, у которого «субкосмоцерасовая» стадия онтогенеза (соответствующая взрослому состоянию его потомка Kosmoceras) обнаруживает индивидуальные уклонения в сторону увеличения ее длительности за счет взрослой кепплеритной стадии [412].
Что же касается явлений, как будто бы показывающих «скачкообразную остановку» развития (на деле всегда осложненную гетерохрониями в развитии репродуктивной системы и другими перестройками), то они представляют собой в действительности следствие эволюции, а не ее причину. Эти явления существуют лишь в виде факультативной неотении, т. е. разновидности созданного отбором модификационного полиморфизма, когда развитие в зависимости от внешние условий либо останавливается на определенной стадии, либо идет до конца. Общеизвестный пример — мексиканская саламандра Ambystoma tigrinum среди хвостатых амфибий. Как и все адаптивные модификации, «скачки» такого рода легко обратимы в поколениях, и соответственно в плейстоценовой истории мексиканской амбистомы, прослеженной на ископаемом материале, отмечается чередование двух ее адаптивных состояний в зависимости от смены похолодании и потеплении [2383].
Следует упомянуть о том, что, признавая раннеонтогенетические сальтации в качестве модуса эволюции, исследователи не обязательно имеют в виду одновременное распространение их действия на поздние стадии, т. е., по существу, могут подразумевать просто одноактное возникновение ценогенетических изменений. Иногда эти гипотетические явления достаточно четко отделяются от «классических» архаллаксисов и девиаций. Например, у О. Шиндевольфа [2223] такой скачок, или «типогенез», предшествует постепенному переходу онтогенетического новшества во взрослое состояние. А. С. Северцов [932] выделяет такие события как особые модусы филэмбриогенеза, действующие как архаллаксисы и девиации, но только в отношении промежуточных стадий, а не взрослой. Иногда, однако, эти различия не акцентируются. Так, у колониальных кораллов в качестве архаллаксиса [101] был описан переход личинки-основательницы от бокового прикрепления к осевому и превращение протокораллита-капсулы в базальный диск — безотносительно к ходу позднего развития, которое может при этом не затрагиваться изменениями.
Во всех подобных случаях вывод о способе эволюции ранних стадий развития основан на том же самом подходе, что и при обосновании скачкообразных изменений взрослой стадии, а именно, на отождествлении различия двух онтогенезов с историческим событием. В действительности, как уже было показано выше, каждый отрезок онтогенеза, завершающийся более или менее устойчивым морфологическим типом, эволюирует в общем случае, как и взрослая стадия, путем терминальных изменений и их последующего стабилизационного углубления.
«ЭМБРИОНАЛЬНЫЕ МОДУСЫ» И МУТАЦИОНИЗМ
Представления об эволюционно значимых изменениях раннего онтогенеза, одновременно меняющих взрослую стадию, основываются отнюдь не только на сравнительно-эмбриологических доводах. По существу, речь должна идти об обоснованиях всей совокупности мутационистских взглядов на эволюцию, ибо предполагаемое ими скачкообразное изменение организмов невозможно представить себе иначе, как результат ранних индивидуальных нарушений нормального хода онтогенеза. Необходимость допущения таких уклонений или даже перестройки всего развития подчеркивается многими авторами, признающими эволюционные сальтации, — например, когда говорится о «системных мутациях» [1650] или «онтомутациях» (1482], или же когда глубокие мутационные аномалии прямо отождествляются с арахаллаксисами А. Н. Северцова [2142]. Однако чаще этот аспект проблемы остается без внимания.
Независимо от способов аргументации в основе мутационизма всегда лежит явное или неосознанное убеждение, что между нормальным и аномальным итогом онтогенетического развития нет принципиальной разницы в отношении эволюционной устойчивости. Неприемлемость этого взгляда (см. "Эволюция как эпигенетический процесс") делает концепцию скачков несостоятельной для объяснения магистрального эволюционного процесса; однако следует помнить, что мир природных явлений бесконечно разнообразен, и нельзя исключить априори возможность осуществления в нем относительно маловероятных событий. К тому же граница между «ранним» и «поздним» онтогенезом, отделяющая область регулируемых или летальных уклонений от области эволюционно значимых вариаций, меняющих взрослый фенотип, всегда достаточно условна, и, строго говоря, мы можем лишь утверждать, что вероятность сохранения отбором онтогенетических изменений снижается тем резче, чем раньше они наступают. Поэтому важно определить, в каких случаях теоретически возникает наибольшая вероятность уклонения от типичных общих закономерностей онтогенетической эволюции, рассмотренных выше. Рассмотрим основные принципиальные аргументы в пользу мутационизма, относящиеся к собственно морфологической эволюции.
На первый взгляд, первостепенная роль здесь принадлежит палеонтологии, так как, начиная со времен Ж. Кювье и А. д’Орбиньи, она многократно порождала сальтационистские концепции (неокатастрофистские идеи О. Иекеля, Д. Н. Соболева, О. Шиндевольфа, Р. Ведекинда, К. Бойрлена, Э. Даккэ и др. [291, 318]) и продолжает выдвигать их и сегодня, например [527, 1878]. Но на деле используемые ею факты не могут играть в нашем вопросе решающей роли в силу самой специфики палеонтологической летописи, не позволяющей с уверенностью отличать истинную смену поколений от случайной преемственности, созданной седиментационными проблемами и локальными миграциями. Между тем для доказательства эволюционно значимых скачков необходимым (хотя и не достаточным) условием является именно регистрация их у непосредственных потомков. С этой точки зрения более существенны свидетельства, относящиеся к рецентным организмам.
Один из самых традиционных доводов в пользу скачкообразной эволюции, ведущий начало от Э. Жоффруа Сент-Илера, заключается в указаниях на появление в природе резких аномалий и, вообще, дискретных изменений как на путь возникновения новых форм (А. Келликер, У. Бэтсон и др.). Особое развитие он получил в контексте критики дарвиновской теории, когда ее главному принципу отбора мелких вариаций стали противопоставляться бесспорные случаи спонтанного появления у животных и особенно у растений (преимущественно домашних или культивируемых) крупных наследственных уклонений, давших начало новым устойчивым формам (С. И. Коржинский, Г. де Фриз и др.). Но еще Ч. Дарвин [310] указывал, что такие аномалии, используемые в практике селекции, имеют мало шансов закрепиться в природных условиях в связи с их редкостью и нарушением в них нормального развития, ведущим к снижению приспособленности.
Это возражение, в основном сохраняющее свою силу и сегодня для мира животных, часто считается менее весомым в отношении растений в связи с накоплением данных, свидетельствующих о большое жизнеспособности многих растительных мутантов в природе. Последнее заставляет многих авторов, даже мало склонных к мутационистским представлениям, допускать скачкообразное возникновение отдельных растительных групп через посредство таких аномалий [1194, 2249, 2520].
В последнее время оспаривают также и другое возражение Ч. Дарвина — о единичности и малой вероятности сохранения крупных аномалий в природе. При этом приводят случаи, когда подобные аномалии (большей частью экзогенного характера) дают широкие вспышки в небольших популяциях и могут удерживаться достаточно долго. Сюда относятся случаи массовой полимелии (появления лишних конечностей) и полидактилии (многопалости) у современных амфибий. На этой основе Л. Ван Валеном выдвинута новая модель эволюции, в которой подобные массовые аберрации служат объектом отбора (генетической ассимиляции) и, таким образом, становятся не зависимыми от первично вызвавшего их внешнего фактора [2420].
Весьма существенным доводом в пользу спонтанного появления новых таксонов у животных и растений считают факты близкого параллелизма между аномалиями одних родов и видов и нормальной организацией у других [1174, 1650, 2323]. Например, бескрылые виды мух по типу рудиментации крыльев могут очень напоминать соответствующих мутантов у дрозофилы. Особенно удивительными эти параллелизмы кажутся исследователям в тех случаях, когда речь идет об изменении числа однотипных органов или о переносе полового признака на противоположный пол. Так, по мнению В. М. Шимкевича [1174], наличие лишних ротовых отверстий у одного из реснитчатых червей или добавочной пары рогов у антилопы Tetraceros указывают на их происхождение из мутаций, аналогичных таким же уродствам у близких таксонов.
Но, пожалуй, самым распространенным источником представлений о скачкообразных эволюционных изменениях является господствующая сегодня синтетическая теория микроэволюции. Согласно ей, новые формы или расы во многих случаях возникают из старых путем единичной мутации, которая затем распространяется в популяции, приобретая ранг нормы [269, 1509]. С этих позиций обычно трактуются и популяционные изменения у ископаемых форм, когда распространение какой-либо особенности — например, новой складки зубов в эволюции лошадеобразных или новой формы раковины у двустворок, — расценивается как результат отбора в пользу мутантного гена [1713, 2287]. Сторонники этих взглядов обычно даже не осознают своей приверженности к мутационизму, полагая, что последний непременно связан с допущением крупных скачков типа «системных мутаций» Р. Гольдшмидта [594, 1509]. Но в действительности сущность мутационизма выражается не в масштабах допускаемого эволюционного уклонения, а в том, что оно мыслится возникающим в готовом виде независимо от естественного отбора, который затем лишь увеличивает его частоту.
Наиболее очевидными случаями спонтанных изменений, расцениваемыми нередко как единственная бесспорная возможность мгновенного видообразования [594], большинство эволюционистов считает межвидовую гибридизацию и полиплоидию (т. е. кратное увеличение числа хромосомных наборов в клетках). Различают прежде всего авто- и аллополиплоидию, т. е. соответственно изменения на основе одного видового генома или же двух (и более), объединенных у гибридов. Говоря о таких скачках, обычно имеют в виду не столько обязательное появление морфологических отличий, сколько внезапность репродуктивной изоляции от родителей.
Здесь снова обнаруживается поразительное различие между животным и растительным царствами. Если у растений полиплоиды обоих типов охватывают, по некоторым оценкам, до половины всех покрытосеменных и до 95 % папоротникообразных [269], то у животных они намного более редки и охватывают лишь группы небольшого ранга (у наземных позвоночных — не выше вида). Обычно это автополиплоиды, связанные в огромном большинстве случаев с партеногенезом, но иногда включающие и группы бисексуальных видов, например у бесхвостых амфибий [ср. 594]. Аллополиплоиды (обычно триплоиды) нередко встречаются у животных среди межвидовых гибридов [111], которые в целом также характеризуются партеногенезом или другими формами клонирования (гибридогенез, гиногенез). Переход этих гибридов к однополости объясняется неизбежными нарушениями у них гаметообразования при конъюгации в мейозе нетождественных родительских хромосом, что в свою очередь нарушает оплодотворение.
Попробуем теперь оценить степень доказательности указанных фактов. Прежде всего очевидна неубедительность многочисленных примеров «включения полезной мутации в норму», используемых синтетической теорией эволюции, поскольку под мутациями здесь в действительности подразумеваются уже сложившиеся адаптивные морфы или экотипы, стабилизированные в данное время (или в данной популяции) на низком частотном уровне, — например «мутации» меланизма у грызунов или жилкования крыльев у земляных ос [244, 2519]. Их дальнейшая экспансия означает просто перераспределение частот фенотипов в полиморфных системах. О предыстории таких «мутантных» фенотипов мы ничего не знаем; там же, где она известна хотя бы отчасти (как в случае меланистической морфы у бабочки Biston betularia), факты говорят об их постепенном (а не скачкообразном) формировании [1181].
Наибольший интерес для решения нашей проблемы представляют явления параллелизма между аномалиями и нормальными признаками у разных таксонов. Исследователи, усматривающие в этом доказательство скачкообразного возникновения новых форм, не учитывают, однако, что подобие или тождество двух фенотипов не означает здесь ни их одинаковой устойчивости в поколениях, ни идентичности способов их онтогенетической реализации. С точки зрения представлений о пространстве (ландшафте) системы развития ([1180, 2441], рис. 25, 26, 31, а–в, ср. "Эволюция как эпигенетический процесс") эти способы глубоко различны. Аномалия соответствует индивидуальному уклонению развития от канализированного пути (см. рис. 31, б), означающему снижение его устойчивости, а значит, и воспроизводимости конечного результата. Напротив, эволюционный сдвиг в пользу того же фенотипа есть результат постепенной перестройки самого ландшафта системы, ведущего к стабилизации прежней аберративной траектории. Этот сдвиг суммируется из ряда более мелких шагов, каждый из которых связан с небольшим уклонением развития и его последующей стабилизацией, создающей основу для нового подобного шага (см. рис. 31, а).
В соответствии с этой моделью в онтогенетических системах между нормой и выраженной аномалией (прообразом возможного эволюционного изменения) обычно в самом деле существует ряд непрерывных или дискретных промежуточных состояний, например последовательные градации выражения Крыловой мутации vestigial у дрозофилы, связанные со все более ранними нарушениями онтогенеза [1650]. Поэтому приписывать, например, появление бескрылых видов насекомых скачкообразной утрате крыльев (и вообще отождествлять эволюционное изменение хода развития со скачком), значит, по меньшей мере не замечать реальных морфогенетических возможностей для постепенного приобретения нового признака. И действительно, во многих случаях природной редукции крыльев (среди жесткокрылых) могут наблюдаться самые различные степени ее выражения — как в пределах изменчивости одного вида, так и среди видов или родов одного семейства [1650]. К тому же аналогия между эволюционной и аномальной (мутантной) редукцией нередко оказывается достаточно поверхностной и на деле осложненной многими адаптивными изменениями в нормальном типе, как показывает, например, строение рудиментов крыльев у мухи Amalopteryx mаritima, параллелизуемой Р. Гольдшмидтом с мутацией vestigial у дрозофилы [806].
Рис. 31. Обычный (а) и гипотетические сальтационные (б, в) пути эволюции онтогенеза. Составил М. А. Шишкин, оригинал.
а — постепенное преобразование фенотипа А в В2 путем смещения развития на промежуточные траекторий В, В1 и их последовательной стабилизации (соответствующие типы развития F1, Fn, Fx, Fy разделены рядом преобразуемых отбором поколений); б — скачкообразное уклонение от исходного пути развития А на глубокодивергентную траекторию В в ходе одной смены поколений F1–F2, без участия отбора (классическое мутационистское представление о механизме эволюции); в — такое же, но неустойчивое скачкообразное уклонение с последующей стабилизацией его траектории путем отбора в ряду поколений F2–Fn (модель скачкообразной эволюции Ван Валена)
Все сказанное относится и к специальным случаям подобных параллелизмов, связанных, например, с изменением числа нормальных органов. Мутантные и экзогенные аномалии такого типа обычно также показывают самые разные градации выражения, причем они могут выражаться асимметрично на разных сторонах тела — например, удвоение крыльев у дрозофилы, полимастия и полидактилия у млекопитающих и т. д. [30]. Поэтому трудно усмотреть здесь какие-либо доказательства скачкообразных изменений.
Как уже отмечалось, становление всякого эволюционного новшества неизбежно связано с отбором на канализированность (помехоустойчивость) его развития. Однако остается нерассмотренным вопрос—всегда ли оно идет путем накопления и фиксирования мелких изменений, задевающих конец онтогенеза, или же наряду с этим возможна непосредственная стабилизация (в ряду поколений) резкого одноактного раннеонтогенетического уклонения? Такая постановка вопроса как будто бы примиряет два противоположных подхода, совмещая крупномасштабность изменения с постепенностью его закрепления в ходе отбора, как это, в частности, предполагается упомянутой гипотезой Л. Ван Валена ([2420]; ср. рис. 31, в).
В том, что прямая стабилизация крупных аномалий принципиально возможна, нет, конечно, никаких сомнений. К ней, например, сводится процедура выведения в генетике «чистых линий» мутантов, а также механизм генетической ассимиляции индуцированных аномалий. К этой же категории явлений относятся и многочисленные примеры выведения новых сортов и пород на основе внезапных уклонений, традиционно используемые как возражение против дарвинизма.
Однако все эти примеры, в сущности, подчеркивают одно и то же: поразительный контраст между возможностью стабилизации крупных аномалий при искусственном отборе и отсутствием или редкостью достоверных случаев такого рода в естественных условиях. Причины этого достаточно понятны. Нарушение адаптивного фенотипа, порождаемое глубокой девиацией развития, в общем случае имеет шансы сохраниться и стабилизироваться лишь при одновременном резком и необратимом специфическом изменении внешних условий, дающем такой аномалии адаптивное преимущество. Вероятность подобного события очень мала. Такие ситуации пытались моделировать экспериментально, например, культивируя смешанные популяции дрозофилы в условиях постоянного ветра, когда крыловые мутанты, не способные летать, получали преимущество и вытесняли сдуваемых нормальных особей [1731]; подобные же явления отмечались и в узко локальных сезонных очагах в природе [2]. Однако эти наблюдения имеют не большую доказательную силу, чем любые случаи Искусственной или вообще кратковременной в исторических масштабах селекции. Отбор в пользу любого фенотипа стабилизирует последний лишь применительно к условиям отбора; их исчезновение неминуемо ведет к расшатыванию и утрате этого фенотипа в последующих поколениях. Чтобы понять, насколько природная норма, сбалансированная по отношению к сложному комплексу условий, устойчивее временно стабилизированной аномалии, достаточно вспомнить многократно отмечавшиеся генетиками факты возвращения или приближения «чистых линий» мутантов к нормальному фенотипу при разведении их в свободной культуре или перенесении в природную среду [243, 707]. Отбор в исторически типичных условиях, таким образом, легко сводит на нет эпизодическое закрепление онтогенетических девиаций, лишая его эволюционной перспективы.
Следующий важный вопрос касается возможности скачкообразных изменений через посредство полиплоидии и гибридизации. Наименее значимой в этом плане следует считать автополиплоидию. Если в норме она связана с изменением кариотипа гамет, то это еще не доказывает, что именно так она и возникает в эволюции. Как у животных, так и у растений часто встречается соматическая автополиплоидия (эндополиплоидия), обусловленная изменениями митоза и закономерно охватывающая определенные ткани и органы, преимущественно в конце их развития [979]. Она может проявляться в соматических клетках и как обычная вариация, например, у потомства самок шелковичного червя, искусственно стимулированных к партеногенезу [31].
Поэтому случаи спонтанного появления полиплоидных гамет в природе не могут приниматься априори за достоверную модель эволюционного события. С не меньшей вероятностью мы можем считать, что кратное увеличение гаметного генома, наблюдаемое при типичной автополиплоидии, является в действительности не причиной ее, а, наоборот, конечным этапом ее эволюционного становления. В этом случае она должна сначала возникать митотически на поздних стадиях развития диплоидной формы и лишь затем сдвигаться в поколениях в глубь онтогенеза вплоть до зародышевых клеток.
Единственной очевидной возможностью преобразований, основанных на внезапном изменении типичной структуры зародышевой клетки (зиготы), является межвидовая гибридизация. Однако с точки зрения результата развития это событие в действительности не доказывает ни онтогенетического скачка, ни тем более создания какого-то новшества. Всякое нарушение нормального развития — вызвано ли оно абиотическим фактором, или влиянием чуждой гаметы — означает дестабилизацию, т. е. снижение жизнеспособности. Поэтому теоретически наибольшие шансы закончить развитие и дать нормальное потомство имеют те из гибридных особей, у которых ход онтогенеза остается предельно близким к одному из родительских вариантов, тогда как влияние противоположной гаметы низведено до уровня регулируемой помехи. Иначе говоря, отбор должен вначале благоприятствовать особям, фенотипически максимально близким к одному из родителей.
Косвенным подтверждением этих представлений служат многочисленные экспериментальные факты, показывающие, что ранний эмбриогенез у межвидовых гибридов (иглокожие, рыбы, амфибии) протекает обычно целиком по материнскому типу [357]. Очевидно, что чем позднее это доминирование нарушается, тем менее дестабилизированным оказывается итог развития. Следует добавить, что и при нормальном оплодотворении у огромного большинства животных раннее развитие (иногда вплоть до гаструлы) контролируется исключительно организацией материнской клетки, причем начальный этап функционирования диплоидного генома, по-видимому, сводится лишь к синтезу продуктов, уже существующих в этой клетке [333]. Все это заставляет думать, что у природных гибридных видов сохранение фенотипической близости к одному из родителей (описанное для ряда случаев) может служить существенным указанием в пользу их недавнего возникновения. Вероятное отсутствие скачка при формировании гибридных видов у животных касается не только их морфологии, но и пути перехода к типичному для них однополому размножению. В природе сохраняются зафиксированными различные стадии этого перехода, а именно, гибридогенез (скрещивание гибридов с самцами родительского вида, с последующим уничтожением мужского генома в оогенезе дочерних особей [111]) и гиногенез, где роль отцовской гаметы сведена лишь к стимуляции развития. Мы видим, что механизм включения развития лишь постепенно освобождается здесь от необходимости акта оплодотворения, делая существование гибридной формы все более независимым от контакта с родительскими видами.
Огромное различие в частоте случаев гибридного видообразования у животных и растений, несомненно, заключается в большей пластичности способов размножения у последних [31, 378]. Растительные гибриды через развитие соматической полиплоидии могут дать вегетативным путем обоеполое аллополиплоидное потомство, способное к нормальному гаметообразованию и продуктивному скрещиванию [269]. Напротив, для гибридов животных подобная возможность крайне затруднена, поскольку их существование поддерживается либо возвратным скрещиванием с родителями, либо однополым размножением. Ни в том, ни в другом случае не возникает эквивалентного по кариотипу полового партнера. Поэтому неудивительно, что двуполая аллополиплоидия у животных до сих пор достоверно неизвестна за пределами эксперимента [31, 111].
Проведенный обзор показывает отсутствие сколько-нибудь убедительных доказательств в пользу скачкообразной эволюции онтогенеза. Все приводившиеся с этой целью доводы основаны так или иначе на смещении в сознании исследователей продуктов эволюции с ее сырым материалом (истолкование адаптивный морф в качестве «мутаций», отождествление нормального развития с аберративным, отождествление дискретности нормальных фенотипов с онтогенетическим скачком между ними и т. д.). Верно лишь то, что многие изменения в механизме онтогенеза или в структуре зародышевой клетки создают потенциальную возможность для ускоренного преобразования взрослой организации у потомков, но само оно осуществляется лишь путем последовательной стабилизации в ходе отбора. Среди «подготавливающих» изменений такого рода наиболее специфический случай составляет, несомненно, межвидовая гибридизация, получившая массовое распространение в мире растений. Но поскольку, как мы видели, сама по себе она не доказывает одновременного уклонения взрослой стадии за рамки одного из родительских фенотипов, то возникает вопрос — действительно ли специфика онтогенеза у растений облегчает для них возможность скачкообразной эволюции, как это принято считать?
Этот взгляд кажется сильным упрощением. Систематика растений была бы невозможна, если бы их виды не обладали в общем случае определенной устойчивостью, которая и является выражением саморегуляции их онтогенеза. Последняя же как раз и представляет собой механизм, направленный на эффективное погашение ранних аберраций. Слабая морфогенетическая интегрированность растений, обычно подчеркиваемая как фактор, способствующий их сальтационной эволюции, в действительности играет здесь несколько иную роль. Чем менее интегрирована данная система, тем менее контрастны в ней различия между равновесным состоянием и отклонениями от него и тем более длителен для нее период восстановления нарушенного равновесия. В применении к живым системам это означает, что у растений отклонения от равновесного фенотипа (нормы) оказываются относительно «терпимыми» (жизнеспособными) в более широких пределах, чем у животных, и могут дольше удерживаться в поколениях.
Поэтому масштабы уклонений, для которых потенциально возможно закрепление отбором, у растений значительно шире. Иначе говоря, здесь возрастает вероятность случаев эволюции по схеме Л. Ван Валена — путем стабилизации относительно ранних аберраций. Эти случаи, однако, не означают девиации в общепринятом смысле, поскольку возникающие уклонения составляют здесь лишь сырой материал для создания устойчивой нормы в последующих поколениях. Тем более это не меняет общих закономерностей, рассмотренных нами ранее.
Сравнительный анализ приводит многих ботаников к выводу о преобладании в эволюции онтогенеза растений конечных изменений и о вторичном распространении их на более ранние стадии [2322, 2518], как это имеет место и у животных. К этому надо добавить, что сама по себе возможность утилизации отбором относительно ранних уклонений у растений еще не означает, что она доказана фактически. Наблюдая аномалии развития, мы не можем судить об их эволюционной судьбе; в то же время для уже возникших видов исходный тип развития никогда не может быть установлен вполне достоверно.
О НЕКОТОРЫХ ПРИМЕРАХ «СКАЧКООБРАЗНЫХ ИЗМЕНЕНИЙ»
Рассмотрим подробнее некоторые из перечисленных ранее явлений, используемых для доказательства эмбриональных скачков и относящихся к преобразованиям в рамках уже существующих структур.
Изменения в числе. Сюда относятся классические архаллаксисы А. Н. Северцова (увеличение числа метамеров тела у змей, изменение иннервации брюшных плавников у костистых рыб) и иные примеры — изменение числа щупалец у Cnidaria и т. д. [2141]. Признание в этих явлениях эмбриональных скачков основывается на убеждении, что подобные структуры по своей природе могут возникать лишь дискретно и эмбрионально, как, например, новые сегменты тела у змей по мере его филетического удлинения [928].
В действительности мы снова видим здесь неоправданную параллелизацию хода нормального развития с историческим событием. При этом не учитывается, что на популяционном уровне объектом вариаций является не только число метамерных структур, но и время их закладки, а также длительность проявления. Отсюда очевидны две возможности возникновения таких структур, не отражаемые непосредственно в итоговом онтогенезе: 1) новый метамер может развиваться эмбрионально как регулируемая (временная) вариация, которая в последующих поколениях сохраняется все дольше, пока не переходит, наконец, во взрослое состояние; 2) допустимо думать, что у рептилий время завершения сегментации в хвостовом отделе (где она наступает позднее всего) может варьировать, вплоть до индивидуального появления лишнего метамера в постнатальный период (в филогенезе эта вариация стабилизируется и эмбрионализируется); в пользу этого говорят соотношения у амфибий, у которых число хвостовых метамеров может в норме резко возрастать уже после метаморфоза [2014].
Перенос половых признаков на противоположный пол. Это явление встречается в норме у многих животных, и обычно полагают, что оно происходит в эволюции скачкообразно [1174]. По мнению И. И. Шмальгаузена, более общим правилом является внезапное появление таких «заимствованных» признаков в эмбриональном периоде и лишь затем переход их у потомков во взрослое состояние. В то же время для многих случаев им фактически признается одномоментная перестройка всего развития — появление самцовой окраски у самок насекомых, ящериц, птиц и т. д. [см. 1192]. Механизм переноса объяснялся на основе данных по морфогенезу вторичных самцовых признаков у птиц, показывающих зависимость их детерминации от чувствительности тканей к уровню концентрации мужского (стимулирующего) и женского (тормозящего) гормонов. И. И. Шмальгаузен полагал, что эволюционная стабилизация исходно самцовых признаков, связанная с повышением их чувствительности к действию активирующих факторов и одновременной автономизацией от влияния тормозящих факторов, в конце концов приводит к тому, что концентрации этих веществ у самок делают для них возможной формообразовательную реакцию по самцовому типу, что и вызывает у них скачкообразное изменение.
Между тем вызывает возражения само представление о скачкообразности такого переноса. И. И. Шмальгаузен видит ее доказательство в том, что в популяциях многих видов среди самок типичного облика встречаются с той или иной частотой особи с признаками другого пола — например, самцовая окраска у ряда насекомых, рептилий и птиц; рогатые особи среди безрогих самок копытных и т. д. Но, как уже было сказано, подобные «мутанты» могут представлять собой относительно стабилизированные адаптивные типы внутри полиморфных систем. Спорадичность их проявления нельзя отождествлять со скачкообразностью эволюционного возникновения. Кроме того, множество наблюдений показывает, что переносимые половые признаки могут обнаруживать все градации в степени выражения — от едва заметных зачатков до полного развития, как, например, рога самок у разных видов антилоп или матка у самцов дельфина Phocaena сотmunis. Потенциальная способность любой зиготы к реализации обоих полов, а также различных морфологических переходов между ними («интерсексов») показана многими экспериментальными исследованиями, в особенности по межрасовым скрещиваниям — у шелковичного червя, бесхвостых амфибий и т. д.; при это чем менее значительно уклонение в сторону противоположного пола, тем более поздние стадии развития оно задевает [29, 257].
Все это показывает, что механизмы детерминации половых признаков обеспечивают возможность постепенного перехода между крайними альтернативами и что она действительно реализуется в эволюции путем последовательного накопления и закрепления промежуточных вариаций. Данный процесс может быть сопряжен с поздними онтогенетическими изменениями, которые затем углубляются в поколениях по мере усиления уклонения взрослого признака в сторону противоположного состояния (рис. 32). Или же, наоборот, переносимый признак обнаруживается сначала как временное уклонение раннего развития, а затем постепенно «всплывает» во взрослую стадию.
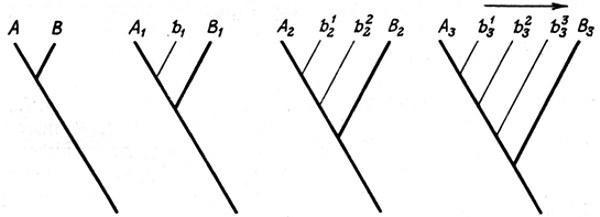
Рис. 32. Эволюционное расширение спектра промежуточных уклонений b1, b2, b3 по мере эмбрионализации развития альтернативных половых признаков A и В и углубления онтогенетической дивергенции между ними — до состояний A3 и В3. Составил М. A. Шишкин, оригинал.
Непосредственный скачок от A3 к B3 возможен лишь как аномалия развития; эволюционно значимое изменение в направлении В3 осуществляется лишь через последовательную стабилизацию промежуточных уклонений (показана стрелкой)
Не подлежит сомнению, что явление переноса половых признаков существует так же давно, как и сам половой диморфизм. А поскольку половые признаки по своему происхождению связаны со взрослой стадией, то и древнейшие их нарушения должны были ограничиваться поздним онтогенезом. По мере филетической эмбрионализации, углубляющей дивергенцию между двумя альтернативными вариантами осуществления полового признака, все более расширяется спектр возможных нарушений выбора между ними. Чем раньше в онтогенезе наступает такое аномальное уклонение, тем больше его результат фенотипически приближается к противоположному нормальному состоянию (см. рис. 32). Нет никаких оснований думать, что эволюция идет путем непосредственного закрепления наиболее крупных из этих аномалий; факты по изменчивости выражения переносимых признаков говорят об обратном. Вместе с тем эмбрионализация развития полового признака создает условия для формирования в его раннем морфогенезе временных аберраций в сторону противоположного пола (таких, как, например, провизорная закладка яйцеводов у самцов млекопитающих).
Таким образом, наиболее вероятно, что эволюционный перенос половых признаков осуществляется путем последовательного закрепления и суммирования отбором небольших уклонений, начинающихся либо непосредственно с исходного взрослого состояния признака, с последующим распространением изменения на предыдущие стадии, либо с одной из ранних стадий, изменение которой может в дальнейшем постепенно перейти в дефинитивное состояние путем фетализации.
Возникновение зеркально-симметричных вариантов. Подобные инверсии, в особенности появление отдельных левозакрученных особей среди гастропод с типичной правозакрученной раковиной (или, реже, наоборот) — один из популярных примеров «архаллаксиса» [359, 2141; 2287]. Различие между двумя вариантами прослеживается здесь с первых же дроблений и может детерминироваться еще в материнском яичнике; скрещивание таких форм дает простое менделевское расщепление [2343]. Морфологические переходы между этими двумя типами невозможны, и обычно принимается, что один возникает из другого путем мутации. Так объясняют появление у гастропод левозавернутых форм.
На самом деле, однако, здесь нет ничего, что говорило бы о скачкообразных эволюционных изменениях. Наличие правильного расщепления ясно указывает, что речь идет о сложившейся полиморфной системе, и вопрос состоит в том, как исторически она возникла. Само по себе наличие право- и левозакрученности (и соответствующих типов асимметрии внутренних органов) — есть следствие конического (турбоспирального) навивания раковины, которое возникло у древнейших гастропод из симметричного планоспирального навивания [61, 1997]. Естественно считать, что исходным материалом для такого изменения послужили незначительные отклонения устья раковины от плоскости спирали и что лишь в последующих поколениях этот процесс стал захватывать все более ранние обороты, вплоть до макушки, соответствующей личиночному протоконху.
Уже первые неустойчивые отклонения такого рода неизбежно должны были осуществляться в двух вариантах — «правом» и «левом», причем их адаптивная ценность едва ли могла заметно различаться (рис. 33). Каждый шаг стабилизации этих отклонений в ходе отбора открывал путь к их дальнейшему усилению благодаря постепенному увеличению амплитуды возможных для них новых вариаций. Таким образом, могло идти параллельное преобразование двух зеркально-симметричных морф и углубление онтогенетической дивергенции между ними (см. рис. 33, I–VI). Реально в эволюции отдельных филумов (или на отдельных ее отрезках) частота одной из таких морф обычно остается очень низкой, что означает ее меньшую адаптивную ценность и недостаточную стабилизацию ее онтогенеза. Но, очевидно, всегда сохраняется потенциальная возможность для усиления отбора в пользу этой морфы (при соответствующем изменении условий) и перехода ее тем самым в статус основной нормы. Вероятный пример таких преобразований показывает для кайнозоя Англии морская гастропода Trophon aniiquum, у которой в плиоцене в условиях относительно теплого климата преобладала левозавернутая форма, тогда как ныне доминирует правозавернутая [2419]. Пространственные изменения частот двух соответствующих морф известны у ряда современных видов (например, Fructicicola lantzi [218].
Все это позволяет предполагать, что перераспределение адаптивных ролей между двумя типами навивания раковины является не только обратимым процессом, но и происходит на фоне их непрерывной параллельной эволюции. По мере углубления и спрямления онтогенетических преобразований, связанных с диссиметри-зацией внутренних органов в этих двух альтернативных вариантах, их дивергенция должна была начинаться в развитии все раньше, пройдя в итоге путь от дефинитивной стадии у билатерально-симметричного предка до первых фаз дробления у потомков.
Рис. 33. Коэволюция двух возможных симметричных состояний фенотипической нормы (на примере право- и левозавернутости у гастропод). Составил М. A. Шишкин, оригинал.
I–VI — последовательные исторические этапы; A1–A6, B1–B6 — последовательные стадии эволюции и эмбрионализации развития альтернативных морф (А и В). Заштрихованы отрезки онтогенеза, следующие за моментом дивергенции двух типов развития. Новый адаптивный признак (А–В) возникает в виде двух слабых и неустойчивых противоположных уклонений взрослой стадии на этапе I, усиливающихся и ускоряющихся в процессе дальнейшей эволюции этапы II–VI. Сужение нормы в пользу стабилизируемого варианта А ведет к снижению частоты симметричной ему морфы В, сохраняющей пониженную устойчивость осуществления (наследования, II– IV). Последующий сдвиг нормы в сторону морфы В стабилизирует ее и дестабилизирует фенотип A (V–VI)
Таким образом, наличие у гастропод двух глубоко дискретных типов развития с наибольшей вероятностью может рассматриваться как итог их постепенного исторического становления. Их предопределенность в яйце говорит об эволюционном скачке не больше, чем хромосомная детерминация пола в зиготе говорит о скачкообразном происхождении полового диморфизма.
Изложенная картина возникновения диморфизма раковины на основе планоспиральной организации взрослого предка требует некоторых дополнительных обоснований. В развитии асимметрии тела гастропод выделяют два основных момента: 1) торсионный процесс, т. е. поворот раковины с внутренностным мешком относительно головы и ноги на 180° и 2) приобретение раковиной турбоспиральной формы и отведение оси ее навивания назад, что вызывает одностороннюю редукцию парных органов. Независимо от того, развивались ли эти изменения синхронно или последовательно, есть все основания связывать их начало с переходом взрослого предка от пелагического обитания к жизни на дне [61, 430, 1997]. Весь ход развития гастропод несет на себе признаки филетического ускорения — «адьюльтации» [1785], приведшего к появлению зачатков ряда взрослых органов (нога, радула, раковина) на личиночных или эмбриональных стадиях. Поэтому и приуроченность торсионного процесса к концу личиночной жизни логично связывать с таким же сдвигом. В пользу справедливости этого мнения говорит сравнение разных типов личинок и в особенности соотношения у примитивного переднежаберного Haliotis, где вторая фаза скручивания наступает лишь после перехода к обитанию на дне и продолжается весьма долго [430]. Мнение У. Гарстанга [1624] о том, что скручивание возникло еще у личинок как результат мутации, вызвавшей одностороннюю редукцию мышц, представляет собой типичный пример отождествления истории возникновения признака с его специализированным морфогенезом [Другое возражение против взрослого происхождения скручивания [695], указывающее на невозможность втягивания тела в раковину при наличии перекреста первично парных мускулов, основано на представлении, что торсионный процесс осуществлялся у предка сразу же во всей полноте. Между тем речь должна идти о его постепенном филетическом усилении, параллельно с редукцией одного из мускулов].
Все это позволяет считать, что сопутствовавший скручиванию переход к турбоспиральному типу навивания (а с ним и появление двух его зеркальных вариантов) начал впервые формироваться на поздних стадиях развития планоспирального предка.
Приведенные данные показывают, что реальная возможность эволюционных изменений на основе онтогенетических скачков крайне мала и при этом, вопреки распространенным представлениям, не имеет достоверных фактических доказательств. Однако из этого вовсе не следует, что эволюция имеет монотонный (градуальный) характер. Дело обстоит как раз наоборот. Каждый элементарный шаг в изменении адаптивной организации является результатом двух событий: 1) дестабилизации прежнего фенотипа, ведущей к уклонениям позднего онтогенеза (повышению изменчивости) и 2) стабилизации одного из таких уклонений в качестве новой нормы. Следовательно, на этом уровне рассмотрения эволюция предстает как постоянное чередование фаз устойчивости и неустойчивости (снижения адаптивности) или соответственно фаз персистирования и направленного изменения ([1180,1181]; см. "Эволюция как эпигенетический процесс"). Отбор направлен на поддержание первых, пока они сохраняют свое адаптивное значение, и скорейшее преодоление вторых. В этом смысле эволюция всегда прерывиста, ибо переход между двумя стабильными состояниями невозможен без снижения стабильности в промежутке. На макроэволюционном уровне этот механизм сдвига нормы служит основой как для постепенных («филетический градуализм»), так и более неравномерных по своим темпам изменений. Однако ни на макро-, ни на микроуровне эта качественная неоднородность эволюционного процесса не имеет отношения к онтогенетическим скачкам (девиациям) и не подтверждает их существования.
Список литературы
2. Аберративный полиморфизм у Drosophila fasciata Meig. (syn. — melanogaster Meig) / Н.П. Дубинин, Д.Д, Ромашев, М.А. Гептиер и др. – Биол. журн., 1937, т. 6, № 2, с. 311-354.
5. Агаев М.Г. Экспериментальная эволюция. Л., Изд-во ЛГУ, 1978.
7. Айала Ф. Механизмы эволюции. – В кн.: Эволюция. М., 1981, с. 33-65.
29. Астауров Б.Л. Генетика пола. – В кн.: Актуальные вопросы современной генетики. М., 1966, с. 65-113.
30. Астауров Б.Л. Исследование наследственных нарушений билатеральной симметрии в связи с изменчивостью одинаковых структур в пределах организма. – В кн.: Наследственность и развитие. М., 1974, с. 54-109.
31. Астауров Б.Л. Экспериментальная модель происхождения раздельнополых полиплоидных видов у животных. - В кн.: Наследственность и развитие. М., 1974, с. 151-165.
39. Бабков В.В. Системный стиль в изучении естественного отбора. – В кн.: Системные исследования. М., 1981, с. 404-419.
43. Балкашина Е.И., Ромашев Д.Д. Генетическое строение популяции Drosophila. – Биол. журн., 1935, т. 4, №1, с. 81-106.
61. Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. М., Наука, 1964, т. 1, 2.
66. Белоусов Л.Д. Целостные и структурно-динамические подходы к онтогенезу. – Журн. общ. биол., 1979, т. 40, №4, с. 514-529.
91. Богословский Б.И. Ранний онтогенез и происхождение климений. – Палеонтол. журн., 1976, №2, с. 41-50.
101.Бондаренко О.Б. Соотношение онто-, гистеро-, асто- и филогенеза у ископаемых колониальных кораллов. – В кн.: Морфогенез и пути развития ископаемых беспозвоночных. М., 1982, с. 3-25.
111. Боркин Л.Я., Даревский И.С. Сетчатое (гибридогенное) видообразование у позвоночных. – Журн. общ. биол., 1980, т.41, №4, с. 485-506.
159. Вейсман А. Лекции по эволюционной теории. Пг., А.Ф. Девриен, 1918.
163. Веригин Б.В. Эмбриогенез рыб и общая теория индивидуального развития организмов. – Вопр. ихтиологии, 1970, т. 10, вып. 2 (61), с. 237-254.
200. Вулых П.В. Морфогенез Janetella gen. nov. нового семейства гелиотоид. – Палеонтол. журн., 1980, №4, с. 39-44.
214. Галл Я.М. И.И. Шмальгаузен и проблема факторов эволюции. Историко-биол. исслед. М., 1980, №8, с. 106-123.
218. Гаузе Г.Ф., Смарагдова Н.П. Потеря в весе и смертность у правозавитых и левозавитых особей улитки Fructicicola lantzi. – Зоол. журн., 1939, т. 18, №2, с. 154-161.
242. Гершензон С.М. «Мобилизационный резерв» внутривидовой изменчивости. – Журн. общ. биол., 1941, т. 2, № 1, с. 85-107.
243. Гершензон С.М. Экспериментальное исследование естественного отбора в мутантных популяциях Drosophila viriles. – Журн. общ. биол., 1941, т.2, №3, с. 395-415.
244. Гершензон С.М. Роль естественного отбора в распространении и динамике меланизма у хомяков (Cricetus cricetus L.). – журн. общ. биол., 1946, т.7, №2, с. 97-127.
245. Гершкович И. Генетика. М., Наука, 1968.
255. Голубовский М.Д. Некоторые аспекты взаимодействия генетики и теории эволюции. – В кн.: Методологические и философские проблемы биологии. Новосибирск, 1981, с. 69-92.
256. Голубовский М.Д. Критические исследования в области генетики. – В кн.: Александр Александрович Любищев. Л., 1982, с. 52-65.
257. Гольдшмидт Р. Механизм и физиология определения пола. М.-Пг., 1923.
261. Горбачик Т.Н. Основные закономерности развития среднеюрских–раннемеловых планктонных фораминифер. – В кн.: Закономерности исторического развития ископаемых организмов. М., 1982, с. 53-68.
269. Грант В. Эволюция организмов. М., Мир, 1980.
272. Григорьева А.Д. Фенетические исследования ископаемых популяций. – В кн.: Фенетика популяций. М., 1982, с. 24-29.
280. Гурвич А.Г. Теория биологического поля. М., Сов. наука, 1944.
291. Давиташвили Л.Ш. История эволюционной палеонтологии от Дарвина до наших дней. М., Изд-во АН СССР, 1948.
306.Дагис А.С. Постэмбриональное развитие брахидия позднепалеозойских и раннемезозойских Terebratulida. – В кн.: Морфологические и филогенетические вопросы палеонтологии. Новосибирск, 1972, с. 22-58 (тр. ИГиГ и СО АН СССР, вып.112).
309. Дарвин Ч. Очерк 1842. – Сочинения, т. III. М.-Л., 1939, с. 82-112.
310. Дарвин Ч. Происхождение видов путем естественного отбора. М., Сельхозгиз, 1952.
311. Действие повышенной концентрации ионов магния и натрия на трансляционном и посттрансляционном уровнях у дрожжей Sacharomyces cerevisae/ Н.Н. Михайлова, В.В. Симапров, В.Л. Тихомирова и др. – Исследования по генетике, 1981, №9, с. 65-76.
318.Дмитриев А.Н., Журавлев Д.И., Кренделев Ф.П. О математических принципах классификации предметов и явлений. – Дискретный анализ, 1966, вып. 7, с. 3-15.
333. Дондуа А.К. Репрограммирование контроля над развитием в раннем онтогенезе многоклеточных животных. – Журн. общ. биол., 1979, т. 40, № 4, с. 530-543.
343. Друщиц В.В., Богословская М.Ф., Догужаева Л.А. Эволюция септальных трубок у аммоноидей. – Палеонтол. журн., 1976, №1, с. 41-56.
353.Дубинин Н.П. Эволюция популяций и радиация. М., Атомиздат, 1966.
354. Дубинин Н.П. О сущности явлений наследственности. – Журн. общ. биол., 1973, т. 34, № 1, с. 3-13.
357. Дэвидсон Э. Действие генов в раннем развитии. М., Мир, 1972.
358. Ежиков И.И. К теории рекапитуляции. – Зоол. журн., 1933, т. 12, вып. 4, с. 57-76.
359. Ежиков И.И. Соотношение онтогенеза и филогенеза. – Успехи совр. биол., 1939, т. 11, вып.2, с. 217-240.
378. Завадский К.М., Колчинский Э.И. Эволюция эволюции. Л., Наука, 1977.
405. Захаров Ю.Д. Онтогенез цератитов рода Pinacoceras и особенности развития подотряда Pinacoceratina. - Палеонтол. журн., 1977, № 4, с. 59-66
412. Иванов А.Н. К вопросу о так называемой профетической фазе в эволюции. – Бюл. МОИП. Отд. геол. 1945, т. 20, № 1, 2, с. 11-32.
414. Иванов П.П. Эмбриональные черты в строении человека. – Человек, 1928, № 2-4, с. 117-133.
415. Иванов П.П. Общая и сравнительная эмбриология. М.-Л., Биомедгиз, 1937.
430. Иванова-Казас О.М. Сравнительная эмбриология беспозвоночных животных. Трохофорные, щупальцевые, щетинкочелюстные, погонофоры. М., Наука, 1977.
441. Инге-Вечтомов С.Г. Система генотипа. – В кн.: Физиологическая генетика. Л., 1976, с. 57-113.
442. Инге-Вечтомов С.Г., Сойдла Т.Р. Эволюционные аспекты проблемы доминирования и молекулярные взаимодействия. – В кн.: Итоги науки и техники. Общая генетика. М., 1978, т. 3, с. 7-37.
443. Иогансен В. Элементы точного изучения наследственности и изменчивости. М.-Л., Сельхозгиз, 1933.
449. Исследование синхронных и параллельных изменений генофондов в природных популяциях плодовых мух Drosophila melanogaster / М.Д. Голубовский, Ю.Н. Иванов, И.К. Захаров и др. – Генетика, 1974, т. 10, №4, с. 73-81.
461. Камшилов М.М. Генотип как целое. – Успехи совр. биол., 1934, т. 3, №2, с. 181-207.
462. Камшилов М.М. Является ли плейотропия свойством гена? – Биол. журн., 1935, т. 4, №1, с. 113-144.
463. Камшилов М.М. Изменчивость и проявление. Проблема нормального фенотипа. – Докл. АН СССР, 1940, т.29, №3, 239-243.
464. Камшилов М.М. К вопросу об отборе на холодоустойчивость. – Журн. общ. биол., 1941, т. 2, № 2, с. 211-227.
465. Камшилов М.М. Эволюция биосферы. М., Наука, 1979.
478. Кастлер Г. Возникновение биологической организации. М., Мир, 1967.
487. Кирпичников В.С. У истоков теории стабилизирующего отбора. – История и теория эволюционного учения, 1974, №2, с. 61-67.
498. Кожанчиков И.В. Об условиях возникновения биологических форм. – Тр. Зоол. ин-та АН СССР, 1941, т. 6, №4, с. 16-32.
503. Кольцов Н.К. Организация клетки. М.-Л., Биомедгиз, 1936.
505. Конюхов Б.В., Нончев С.Г. Экспрессия доминантных и рецессивных признаков в онтогенезе млекопитающих. – Журн. общ. биол., 1981, т. 42, №3, с. 325-334.
527. Красилов В.А. Эволюция и биостратиграфия. М., Наука, 1977.
540. Крыжановский С.Г. Принцип рекапитуляции и условия исторического понимания развития. – В кн.: Памяти академика А.Н. Северцова. М.-Л., 1939, с. 281-366.
565. Лебедкин С.И. БІягенетичны закон І тэория рэкапІтуляцый. – Менск, Изд-во Бел. Ан, 1936. 386 с.
566. Лебедкина Н.С. Эволюция черепа амфибий. М., Наука, 1979.
594.Майр Э. Зоологический вид и эволюция. М., Мир, 1968.
596. Майр Э. Популяции, виды и эволюция. Мир, 1974.
597. Майр Э. Эволюция. – В кн.: Эволюция. М., 1981, с. 11-32.
627. Маслакова Н.И. Индивидуальное развитие глоботрунканид. – Вопр. микропалеонтологии, 1969, № 12, с. 95-107.
629. Маслакова Н.И. Некоторые особенности морфогенеза раковин меловых планктонных фораминифер семейства Globotruncanidae. – В кн.: Закономерности исторического развития ископаемых организмов. М., 1982, с. 35-52.
632. Машковцев А.А. Смена эндогенных и экзогенных факторов эмбрионального развития в онтогенезе и филогенезе. – Изв. АН СССР. Сер. биол., 1936, №5, с. 945-997.
667. Мейен С.В., Шрейдер Ю.А. Методологические аспекты теории классификации. – Вопр. философии, 1976, № 12, с. 67-79.
669. Мейнард-Смит Дж. Эволюция полового размножения М., Мир, 1981.
670. Мейстер Г.К. Критический анализ основных понятий генетики. М.-Л., Сельхозгиз, 1934.
695. Миничев Ю.С., Старобогатов Я.И. Проблема торсионного процесса и проморфологические перестройки у личинок трохофорных животных. – Зоол. журн., 1972, т.51, с. 1437-1449.
703. Михайлова И.А. Соотношение онтогенеза и филогенеза на примере меловых аммоноидей. – В кн.: Морфогенез и пути развития ископаемых беспозвоночных. М., 1982, с. 125-148.
707. Морган Т.Г. Структурные основы наследственности. М.-Л., Госиздат, 1924.
708. Морган Т.Г. Значение генетики для физиологии и медицины (Нобелевская лекция). – В кн.: Избранные работы по генетике. М.-Л., 1937, с. 256-271.
709. Морган Т.Г. Развитие и наследственность. М.-Л., Биомедгиз, 1937.
719. Мюллер Ф. За Дарвина. – В кн.: Мюллер Ф., Геккель Э. Основной биогенетический закон. М.-Л., 1940. с. 61-166.
750. Новинский Ч. Теория эволюции как теория процесса самоорганизации. – В кн.: Проблемы взаимосвязи организации и эволюции в биологии. М., 1978, с. 72-102.
777. Оленов Ю.М., Хармац И.С. Динамика генного состава природной популяции Drosophila melanogaster. Докл. АН СССР, 1938, т. 19, №5, с. 409-415.
806. Парамонов А.А. Дарвинизм. М., Просвещение, 1978.
848. Промптов А.Н. Ген и признак в онтогенезе. – Успехи совр. биол., 1934, т. 3, №2, с. 145-180.
861. Раузер-Черноусова Д.М. Об онтогенезе некоторых палеозойский фораминифер. – В кн.: Памяти академика А.А. Борисяка. М.-Л., 1949, с. 339-353 (Тр. ПИН АН СССР, т. 20).
887. Руженцев В.Е. Принципы систематики, система и филогения палеозойсикй аммоноидей. М.-Л., Изд-во АН СССР, 1960 (Тр. ПИН АН СССР, т. 83).
898. Рьюз М. Философия биологии. М., Прогресс, 1977.
909. Самохвалова Г.В. Получение наследственных изменений у тлей при перемене кормовых растений. – Журн. общ. биол., 1951, т. 12, №3, с. 176-191.
920. Светлов П.Г. О целостном и элементаристическом методах в эмбриологии. – Архив анат., гист. и эмбриол., 1964, т. 46, №4, с. 3-26.
921. Светлов П.Г. Онтогенез как целенаправленный (телеономический) процесс. – Архив анат., гист. и эмбриол., 1972, т. 63, №8, с. 5-16.
922. Светлов П.Г. Физиология (механика) развития. Л., Наука, 1978.
926. Северцов А.Н. Морфологические закономерности эволюции. М.-Л., Изд-во АН СССР, 1939.
927. Северцов А.Н. Эволюция и эмбриология. – В кн.: Северцов А.Н. Собр. соч., т.3. М.-Л., 1945, с. 7-18.
928. Северцов А.Н. Этюды по теории эволюции. – В кн.: Северцов А.Н. Собр. соч., т.3. М.-Л., 1945, с. 19-216.
930. Северцов А.Н. О соотношениях между онтогенезом и филогенезом у животных. – В кн.: Северцов А.Н. Собр. соч., т.3. М.-Л., 1945, с. 316-452.
931. Северцов А.Н. Этюды по редукции органов позвоночных животных. – В кн.: Северцов А.Н. Собр. соч., т. 3. М.-Л., 1945. с. 453-515.
932. Северцов А.С. К вопросу об эволюции онтогенеза. – Журн. общ. биол., 1970, т.31, №2, с. 222-234.
952. Симпсон Дж.Г. Темпы и формы эволюции. М., Изд-во иностр. лит. 1948.
965. Смирнова Т.Н. Некоторые особенности филогенетического развития семейства Dalliniidae Beecher. – В кн.: Закономерности развития органического мира по данным палеонтологии. Л., 1968, с. 63-73 (Тр. XII сессии Всесоюзн. палеонтол. об-ва).
979.Сойдла Т.Р., Вахтин Ю.Б. Генетические основы дифференцировки. – В кн.: Физиологическая генетика. Л., 1976, с. 191-253.
999. Солбриг О., Солбриг Д. Популяционная биология и эволюция. М., Мир, 1982.
1057. Тимофеев-Ресовский Н.В., Иванов В.И. Некоторые вопросы феногенетики. – В кн.: Актуальные вопросы современной генетики. М., 1966, с. 114-130.
1078. Уоддингтон К.Х. Зависит ли эволюция от случайного поиска? – В кн.: На пути к теоретической биологии. М., 1970, с. 11-38.
1079. Уоддингтон К.Х. Основные биологические концепции. - В кн.: На пути к теоретической биологии. М., 1970, с. 108-115.
1104. Филипченко Ю.А. Эволюционная идея в биологии. М., М. и С. Сабашниковы, 1926.
1141. Четвериков С.С. О некотрых моментах эволюционного процесса с точки зрения современной генетики. – Журн. эксперимент. биол., 1926, сер. А, №2, с. 91-96.
1147. Шапошников Г.Х. Специфичность возникновения адаптаций к новым хозяевам у тлей (Homoptera, Aphidoidea) в процессе естественного отбора (экспериментальные исследования). – Энтомол. обозр. 1961. т.40, №4, с. 739.
1148. Шапошников Г.Х. Морфологическая дивергенция и конвергенция в эксперименте с тлями (Homoptera, Aphidoidea). – Энтомол. обозр. 1965. т.44, №1, с. 3-25.
1149. Шапошников Г.Х. Возникновение и утрата репродуктивной изоляции критерий вида. – Энтомол. обозр. 1966. т.45, №1, с. 3-35.
1150. Шапошников Г.Х. Динамика клонов, популяций и видов и эволюция. – Журн. общ. биол., 1978, т. 39, №1, с. 15-33.
1164. Шеппард Ф.М. Естественный отбор и наследственность. М., Просвещение, 1970.
1174. Шимкевич В.М. Уродства и происхождение видов. СПб.-М., Вольф, 1907.
1176. Шишкин М.А. Морфология древних земноводных и проблемы эволюции низших тетрапод. М., Наука, 1973 (Тр. ПИН АН СССР, т. 137).
1177. Шишкин М.А. О роли модификаций в микроэволюции. – В кн.: Микро- и макроэволюция. Тарту, 1980, с. 136-139.
1178. Шишкин М.а. Закономерности эволюции онтогенеза. – Журн. общ. биол., 1981, т.42, №1, с. 38-54.
1179. Шишкин М.А. Морфологическая эволюция онтогенеза. – В кн.: Современные проблемы эволюционной морфологии животных. 1981, с. 119-120 (Тез. докл. междунар. симпозиума).
1180. Шишкин М.А. Индивидуальное развитие и естественный отбор. – Онтогенез, 1984, т. 15, №2, с. 115-136.
1181. Шишкин М.А. Фенотипические реакции и эволюционный процесс. – В кн.: Экология и эволюционная теория. Л., 1984, с. 196-216.
1182. Шишкин М.А. Эпигенетическая система и менделевский фактор. – В кн.: Методологические проблемы эволюционной теории. Тарту, 1984, с. 74-76.
1184. Шмальгаузен И.И. Изменчивость и смена адаптивных норм в процессе эволюции. – Журн. общ. биол., 1940, т.1, №4, с. 509-528.
1185. Шмальгаузен И.И. Пути и закономерности эволюционного процесса. М.-Л., Изд-во АН СССР, 1940.
1186. Шмальгаузен И.И. Стабилизирующий отбор и его место среди факторов эволюции. – Журн. общ. биол., 1941, т.2, №3, с. 307-354.
1192. Шмальгаузен И.И. Факторы эволюции. Теория стабилизирующего отбора. М., Наука, 1968.
1194. Шмальгаузен И.И. Проблемы дарвинизма. Л., Наука, 1969.
1196. Шмальгаузен И.И. Организм как целое в индивидуальном и историческом развитии. М., Наука, 1982.
1199. Шпеманн Г. Наследственность и механика развития. – Журн. эксперим. биол., 1925, Сер Б., т.1, № 3-4, с. 119-144.
1224. Эшби У.Р. Конституция мозга. М., Изд-во иностр. лит., 1962.
1261. Alberch P. Ontogenesis and morphological diversification. – Amer. Zool., 1980, v. 20, p. 653-667.
1262. Alberch P. Developmental constraints in evolutionary processes. – In: Evolution and development / Ed. J.T. Bonner. Heidelberg, Springer Verlag, 1982, p. 313-332.
1291. Baer C. Ueber Entwicklungsgeschichte der Tiere. Koenigsberg, Borntraeger, 1828, Bd.1.
1299. Balfour F.M. Address to the Department of the British Association for the Advancement of Scince. – In: The works of F.M. Balfour. London, Macmillan, 1880, v.1, p. 698-713.
1317. Beer G.R., de. Embryology and evolution. Oxford, Clarendon Press, 1930.
1319. Beer G. R., de. Embryos and ancestors. London, Oxford Univ. Press, 1958.
1377. Bridges C. The origin of variations in sexual and sexlimited characters. – Amer. Natur., 1922, v. 56, p. 51-63.
1380. Broman I. Das sogenannte “Biogenetische Grundgesetz” und die moderne Ergeblichkeitlehre. Munchen, 1920.
1386. Buchman S.S. Type ammonites. New York, Cramer, 1920, v. 3.
1410. Carter G.S. Animal evolution: a study of recent views of its causes. London, Sedgwick et Jackson, 1951.
1448. Conclin E.G. The organization of celi-lineage of the ascidian egg. – J. Acad. Natur. Sci. Philadelphia, 1905, v.13, p. 1-119.
1457. Cope E.D. The primary factors of organic evolution. – Chicago, Open. Court Publ. Co., 1904.
1482. Dalcq A. Les ontomutation a l’origine des mammiferes. – Bull. Soc. zool. France, 1954, t. 79, N 1, p. 240-255.
1509. Dobzhansky Th. Genetics and the origin of species. New York, Columbia Univ. Press., 1947.
1524. Driesch H. The science and philosophy of the organism. London, Black, 1908.
1535. Easton W.H. Invertebrate paleontology. New York, Harper and Row, 1960.
1562. Erdmann K. Zur Entwicklung des Knochernen Skeletts von Triton und Rana unter besonderer Berucksichtigung der Zeitfolge der Ossifikationen. – Z. Anat. Entwicklungsgesch., 1933, Bd. 101, N 5-6, S. 566-651.
1604. Franz V. Ontogenie und Phylogenie. – Abh. Theorie organisch. Entwickl., 1927, N.F., N 3.
1620. Gans C., Parsons T.S. On the origin of the jumping mechanism in frogs. – Evolution, 1966, v.20, p. 92-99.
1623. Garstang W. Theory of recapitulation. A critical restatement of the biogenetical law. – J. Linn. Soc. London, 1922, v. 35, p. 81-103.
1624. Garstang W. The origin and evolution of larval forms. – Rep. Brit. Assoc., 1928, v.77, p. 1-98.
1649. Goldschmidt R. Physiological genetics. New York-London, McGraw-Hill, 1938.
1650. Goldschmidt R. The material basis of evolution. New Haven, Yale Univ. Press, 1940.
1651. Goldschmidt R. Theoretical genetics. Los Angeles, Univ. Calif. Press, 1955.
1661. Gould S.J. Ontogeny and phylogeny. Cambridge – London, Belknap Press, 1977.
1674. Griffiths J. The phylogeny of Salientia. – Biol. Rev., 1963, v.38, p. 241-292.
1683. Gurwitsch A. Ueber Determination, Normierung und Zufall in der Ontogenese. – Roux Arch., 1910, Bd. 30, S. 133-193.
1684. Gurwitsch A. Die Vererbung als Verwirklichungsvorgang. – Biol. Zbl., 1912, Bd. 32, S. 458-486.
1687. Haeckel E. Generelle Morphologie der Organismen. Berlin, G. Reimer, 1866. Bd. 1, 2.
1688. Haeckel E. Die Gastrula und die Eifurchung der Thiere. – Jenaische Z. Naturwiss., 1875, Bd. 9, S. 402-508.
1692. Haldane J.B.S. The time of action of genes and its bearing on some evolutionary problems. – Amer. Natur., 1932, v. 66, p. 5-24.
1713. Hayami I. Possibility of the “palaeogenetics”. – J. Geol. Soc. Japan, 1972, v. 78, N9, p. 495-506.
1719. Hecht M.K. Morphological transformation, the fossil record and the mechanisms of evolution: A debate. Pt. 1. The statement and the critique. – Evol. Biol., 1974, v. 7, p. 215-303.
1731. L’Heritier Ph., Neefs Y., Teissier G. Apterisme des insects et selection naturelle. – C. r. Acad. Sci., 1936, t. 204. p. 907-909.
1733. Hertwig O. Uber die Stellung der vergleichenden Anatomie zur Systematik und Deszendenztheorie. – In: Handb. der Vergleichend u. experimentellen Entwicklungslehre der Wirbelthiere. Jena, Fisher, 1906, Bd. 3, S. 149-180.
1741. Ho M.-W., Saunders P.T. Beyond neo-Darwinism – an epigenetic approach to evolution. – J. Theor. Biol., 1979, v. 78, p. 573-591.
1745. Hoffman A. Paleobiology at the crossroads: a critique of some modern paleobiological research programs. – In: Dimensions of Darwinism / Ed. M. Grene. Cambridge, Cambridge Univ. Press, 1983, p. 241-271.
1763. Hughes N.F. Palaeosuccession of earliest angiosperm evolution. – Bot. Rev., 1977, v. 43, N 1, p. 105-127.
1767. Hurst C.H. The recapitulation theory. – Natur. Sci., 1983, v.2, p. 195-200, 364-369.
1775. Hyatt nA. Genesis of the Arietidae. – Smiths. Comtrib. knowl., 1889, v. 26, N 673, p. 1-238.
1776. Hyatt A. The phylogeny of an acquired characteristic. – Proc. Amer. Phil. Soc., 1894, v. 32, N 143, p. 349-647.
1783. Jaanusson V. Morphological discontinuities in the evolution of graptolite colonies. – In: Animal Colonies/Ed. R.S. Boardman, A.H. Chutham, W.A. Oliver. Stroudsburg, Dowden, Hutchinson and Ross, 1973,p. 515-521.
1785. Jagersten G. Evolution of the metazoan life cycle. London, Academic Press, 1972.
1822. Keibel F. Das biogenetische Grubdgesetz und die Caenogenese. – Ergebn. Anat. und Entwicklungsgesch., 1989, Bd. 7, S. 722-792.
1826. Kemp N.E., Hoyt J.A. Sequence of ossification in the skeleton of growing and metamorphosing tadpoles of Rana pipiens. – J. Morphol., 1969, v. 129, N 4, p. 415-444.
1839. Kojima K.I. Is there a constant fitness value for a given genotype? – Evolution, 1971, v. 25, p. 281-285.
1863. Lang W.D. The evolution of ammonites. Proc. Geol. Assoc., 1919, v. 30, p.49.
1869. Lebedkin S.I. The recapitulation problem. Pt. 1. – Biol. Gen., 1937, Bd. 13, p. 391-417.
1878. Lewin R. Evolutionary theory under fire. – Science, 1980, v. 210, N 4472, p. 883-887.
1879. Lewontin R.C. On the irrelevance of genes. – In: Towards a theoretical biology / Ed. C. Waddington. Chicago, Aldine, Co., 1970, p. 67-88.
1892. Lovejoy A.O. Recent criticism of the Darwin theory of recapitulation: its grounds and its initiators. – In: Forerunners of Darwin. Baltimore, J. Hopkins Unniv. Press, 1959, p. 438-458.
1895. Lovtrup S. Epigenetics. New York, J. Wiley, 1974.
1896. Lovtrup S. On von Baerian and Haeckalian recapitulation. – Syst. Zool., 1978, v. 27, N 3, p. 348-352.
1910. Mac Gillavry H.J. Mutation pressure: a possible cause of directional evolution in Cycloclypeus Carpenter (Foraminifera). -–Verhandl. Koniklijk Nederlandse Geol. – Mijnbouwkundig Genootschap (Geol. Ser.), 1956, v. 16, p. 296-308.
1945. Mehnert E. Biomechanik, erschlossen aus den Prinzipien der Organogenese. Jena, Fisher, 1898.
1970. Morgan T.H. A critique of the theory of evolution. Princeton, Princeton Univ. Press, 1916.
1984. Muller A.H. Lehrbuch der Palaozoologie. Bd. 1. Allgemeine Grundlagen. Jena, Fischer, 1976.
1985. Muller A.H. Abberant Graptoloidea (Pterobranchiata) aus dem Silur (Teil 2). – Z. geol. Wiss, 1977, Bd. 5, N 1, S. 83-93.
1997. Naef A. Studien zur generellen Morphologie der Mollusken. – Ergebn. und Forschr. Zool., 1913, Bd. 3, S. 329-462.
2014. Noble G.K. The biology of the Amphibia. New York, 1931.
2021. O’Donald P. Genetic load of an evolutionary change. – Nature, 1967, v. 216, N 5122, p. 1348.
2027. Olson E.C. Fauna of the upper Vall and Choza. 6. Diplocaulus. – Fieldiana. Geol., 1952, v. 10, p. 147-166.
2059. Ottow R. Die erbbendingte Osteogenesis dysplastico-exostotica der ausgerotteten flugunfahigen Reisentauber Pezophaps solitaria der Maskareneninsel Rodriguez. – K. Svenska Vetensk. Akad. Handl., 1950, Avd. 4, v. 1, N 9, S. 1-37.
2069.Patterson B. Rates of evolution in taeniodonts. – In: Genetics, paleontology and evolution?Ed. G.L. Jepsen, E. Mayr, G.G. Simpson. Princeton, Princeton Univ. Press, 1949, p. 243-278.
2078. Peter K. Palingenese und Coenogenese in der Embryologie. – Gegenbaurs Morphol. Jahrb., 1955, Bd. 94, S. 65-110.
2079. Peters D.S. Braucht man das “Biogenetische Grundgesetz?” – Aufsatze und Reden Senckenberg Naturforsch. Ges., 1975, N 27, S. 16-24.
2115. Rachootin S., Thomson K.S. Epigenetics, paleontology and evolution. – In : Evolution today/ Ed. G.G. Scudder, J.L. Reveal. Pittsburg, Carnegie-Mellon Univ., 1981, p. 181-193.
2124. Raup D.M. Geometric analysis of shell coiling: general problem. – J. Paleontol., 1967, v. 41, N 1, p. 43-65.
2141. Remane A. Die Grundladen des naturlichen System, der vergleichenden Anatomie und der Phylogenetik. Theoretische Morphologie und Systematik 1. Leipzig, Akad. Verlagsgesellschaft, Geest und Portig K.-G., 1956.
2142. Rensch B. Evolution above the species level. New York, Columbia Univ. Press, 1960.
2184. Roux W. Gesammelte Abhandlungen uber Entwicklungsmechanik der Organismen. – Leipzig, Engelmann, 1895, Bd. 1, 2.
2185. Roux W. Terminologie der Entwicklungsmechanik der Tiere und Pflanzen. Leipzig, Engelmann, 1912.
2198. Saint-Hilaire G. Sur le degree d’influence du monde ambient pour modifier les formes animals, question interessant l’origine des especes teleosauriennes et successivement celle des animaux de l’epoque actuelle. – Mem. Acad. Sci., 1833, t. 12, p. 63.
2223. Schindewolf O. Grundfragen der Palaontologie. Geologische Zeitmessung. Organische Stammesentwicklung. Biologische Systematik. Stuttgart, Springer, Verl., 1950.
2249. Schwanitz F. Genetik und Evolutionforschung bei Pflanzen. – In: Die Evolution der Organismen. Bd. 2. Stuttgart, Fischer, 1974.
2259. Sedgwick A. The influence of Darwin on the study of animal embryology. – In: Darwin and modern science/ Ed. A. C. Seward. Cambridge, 1910, p. 171-184.
2284. Siewing H. Lehrbuch der vergleichenden Entwicklugsgeschichte der Tiere. Hamburg-Berlin, 1969.
2287. Simpson G.G. The major features of evolution. New York, Columbia Univ. Press, 1953.
2295. Smith J.P. The biogenetic law from Stadpoint of paleontology. – J. Geol., 1900, v.8, N 1, p. 413-425.
2319. Starett P.H. Evolutionary patterns in larval morphology. – In: Evolutionary biology of the anurans/ Ed. J.L. Vial. Columbia, 1973, p. 251-271.
2322. Stebbins G.L. Flowering plants. Evolution above the species level. Cambridge Mass., Belknap Press of Harvard Univ. Press, 1974.
2323. Steenis C.G. Plant speciation in Malesia, with special reference to the theory of non-adaptive salutatory evolution. – Biol. J. Linn. Soc., 1969, v.1, p. 97-133.
2343. Sturtevant A.K. Inheritance of direction of coiling in Limnea. – Science, 1923, v. 58, N 1501, p. 269-270.
2362. Tasnadi-Kubacska A. Palaopatalogie. Jena, Fischer, 1962, Bd. 1.
2375. The role of development in macroevolutionary change/ P.F. Maderson, P. Alberch, B.C. Goodwin et al. – In: Evolution and development/ Ed. J.T.Bonner. Heidelber, Sprinder Verlag, 1982, p. 279-312.
2383. Tiffney B.H., Barghoorn E.S. The fossil record of the Fungi. – Occas. Pap. Farlow Herbar. Cryptogam. Bot., 1974, N 7, p. 1-42.
2389. Tompa A. Fossil eggs of the land snail genus Vallonia (Pulmonata: Valloniidae). – Nautilus, 1976, v. 90, p. 5-7/
2419. Van Valen L. Variation genetics of extinct animals. – Amer Natur., 1969, v. 103, N 931, p. 193-224.
2420. Van Valen L. A natural model for the origin of some higher taxa. – J. Herpetol., 1974, v. 8, p. 109-121.
2439. Waddington C.H. Canalization of development and the inheritance of acquired characters. – Nature, 1942, v. 150, N 3811, p. 563-565.
2440. Waddington C.H. Genetic assimilation of an acquired characters. – Evolution, 1953, v. 7, p. 118-126.
2441. Waddington C.H. The strategy of the genes: a discussion of some asoects of the theoretical biology. London, Allen and Unwin, 1957.
2242. Waddington C.H. Principles of development and differentiation. New York, Macmillan, 1966.
2470. Weismann A. The origin of the markings of caterpillars. – In: A. Eismann. Studies in the theory of descendant. London, Sampson Law, Marston, Searle and Livington, 1881, p. 161-389.
2471. Weismann A. The germ-plasm. A theory of heredity. New York, Scribner, 1898.
2472. Weismann A. Vortrage uber Descendenztheorie. Jena, Fischer., 1904, Bd. 2.
2501. Woodger J.H. On biological transformations. – In: Essays on growth and form presented to d’Arcy Wentworth Thompson/Ed. W.E. Le Gross Clark, P.B. Madawar. Pxford, Clarendon Press, 1945, p. 94-120.
2505. Wright S. An intensive study of the inheritance of color. – Carnegie Inst. Washigton Publ., 1916, v. 241, pt. 11, p. 59-160.
2506. Wurtenberger L. Studien uber die Stammesgeschichte der Ammoniten. Ein geologischer Beweis fur die Darwinische Theorie. Leipzig, 1880.
2518. Zimmermann W. Arbeitswiese der botanischen Phylogenetik und anderer Gruppierungwissenschaften. – In: Handbuch der biologische Arbeitsmethoden, Abt. IX, Bd. 3, Berlin-Wien, 1931. S. 941-1053.
2519. Zimmermann W. Ueber Mutationen in wilden Populationen. – Mitt. Zool. Mus. Berlin, 1933, Bd. 19, S. 439-452.
2520. Zimmermann W. Grundlagen der Evolution. Frankfurt a/M, Klostermann, 1948.