СИСТЕМНАЯ ОБУСЛОВЛЕННОСТЬ ФОРМООБРАЗОВАНИЯ И ЕЕ ПРОЯВЛЕНИЯ В ПАЛЕОНТОЛОГИЧЕСКОЙ ЛЕТОПИСИ
© 2012 г. М. А. Шишкин
Общей основой механизмов формообразования является обеспечение направленности индивидуального развития как целого к взрослой норме. Устойчивость этого целеполагания есть главное свойство живой организации, требующее объяснения. Каждый шаг эволюции есть вынужденная реорганизация онтогенеза как системы в направлении новой конечной цели (равновесного состояния), т.е. новой нормы. Рост устойчивости ее осуществления в последовательных жизненных циклах под действием отбора ведет к перестройке развития данного эволюционного новшества, распространяющейся в направлении ранних стадий. В итоге различие между записью развития новшества и исходным анцестральным онтогенезом все более преобразуется из пролонгирующего (рекапитулирующего) в дивергентное, соответствующее закону Бэра. Изменения, связанные с этим стабилизационным процессом, фиксируются в палеонтологической летописи в виде ряда характерных проявлений — исходной неустойчивости новоприобретений, перестройке их морфогенеза с течением времени и преимущественного сохранения рекапитуляций анцестральных черт у наиболее близких потомков.
I. Целостность развития как контролирующий фактор морфогенеза
Поиск объяснения любого феномена возможен на разных уровнях рассмотрения, отвечающих его расчленению на все более частные компоненты. Выбор каждого из них может быть оправдан для определенных исследовательских задач. Но содержательные законы, описывающие сущность явления как целого, относятся к наиболее верхним из этих уровней. Только в этом свете может быть оценена природа причинных связей, вскрываемых при более частных редукционистских подходах. Суть научного познания как раз и состоит в том, что для явлений, казавшихся независимыми, обнаруживается взаимосвязь и соподчинение общим законам.
Осознание такого порядка вещей неизбежно для любой области исследований, и теоретическая биология является тому ярким примером. Так, попытки свести эмбриональное развитие к мозаике путей клеточной детерминации, характерные для второй половины XIX века, сменились представлением о едином морфогенетическом поле, контролирующем развитие на всех его этапах. В наше время подобную же перестройку испытывает господствующее эволюционное мышление, еще сорок лет назад верившее во всесилие редукционистских принципов неодарвинизма. На смену этому приходит осознание, что “макроэволюция не может быть сведена к теории, где исходными понятиями являются свойства индивидуальных генов”, и что надлежащая теория должна “учитывать требования верхних уровней рассмотрения” (Maderson et al., 1982).
Сказанное предопределяет должный подход к пониманию действия и становления формообразовательных механизмов индивидуального развития — идет ли речь о процессах, наблюдаемых у современных организмов или об их следах в палеонтологической летописи. Все разнообразие формативных актов, осуществляемых в ходе нормального развития, так или иначе гармонизировано в его рамках, т.е. подчинено закономерностям, управляющим онтогенезом как целым. Среди последних наиболее очевидной характеристикой является направленность всего процесса к заданному конечному итогу, т.е. присутствие в нем целеполагающего (телеономического) контроля, как это было впервые подчеркнуто К. Бэром. Это интегральное свойство развития и должно служить отправной точкой для анализа всех его частных процессов.
Целеполагающее поведение есть свойство системных объектов, выражающее их способность к саморегуляции. Нет сомнения, что индивидуальное развитие есть целостная динамическая система, последовательно корректирующая свое состояние через обратные связи в сторону конечного равновесия, т.е. взрослой нормы (Waddington, 1957; Светлов, 1972; Шишкин, 2006).
Все проявления целостности развития, выявляемые эмбриологией, в конечном счете являются выражением его целенаправленности. Прежде всего, это уменьшение размаха изменчивости в ходе развития, установленное еще Бэром. Во-вторых, это явление эквифинальности или устойчивости итога процесса по отношению к способам его осуществления (Roux, 1896; Driesch, 1908). Данное свойство имеет множество проявлений, касающихся как взрослых, так и “установочных” провизорных стадий развития, например, способа гаструляции у многих первичноротых. Эти свойства могут обнаруживаться как экспериментально (разные пути регенерации особи, развитие полноценных зародышей из разделенных бластомеров или же самосборка последних в одно целое), так и в нормальном развитии. К последнему случаю, прежде всего, относятся альтернативные пути размножения (половое и вегетативное) или типы развития (прямое или с метаморфозом) у одних и тех же или близких видов, причем исходные стадии онтогенетического цикла могут быть предельно различны.
В более широком смысле феномен эквифинальности проявляется на уровне исторических изменений, когда одно и то же свойство организации может осуществляться в развитии у предка и потомков на разной формативной основе (см. ниже).
Если говорить о частных процессах, составляющих развитие, то их целеполагающий характер наиболее отчетлив в морфогенезах регуляционного типа, проявляясь, например, в таких феноменах как множественное обеспечение формообразовательных актов или способность компетентного зачатка к одной и той же реакции в ответ на широкий спектр экспериментальных индукционных воздействий.
Главное обобщение эмбриологии, касающееся направленности онтогенеза, одновременно выражает свойственное ей понимание механизма этого процесса. Это представление о целостной детерминации развития, присутствующей уже в зиготе и предопределяющей формирование всех черт организации по принципу от общего к частному (Baer, 1828; Гертвиг, 1895; Светлов, 1964). Весь ход событий, по выражению П.Г. Светлова, рассматривается здесь как “врисовывание” последовательных актов дифференциации в рамки контролирующего их общего плана развития. Развитие понимается как эпигенез, т.е. нарастание многообразия составляющих его факторов в рамках их интегральной обусловленности. Этот принцип подразумевает отсутствие прямых причинных зависимостей между свойствами организма и частными структурами зародышевой клетки.
Как известно, поиски таких зависимостей составляли суть ранних гипотез о механизме развития, основанньк на “целлюлярном” преформизме, т.е. сведении всего онтогенеза к мозаичной совокупности типов клеточной дифференциации. Предполагалось, что они контролируются по отдельности теми или иными клеточными частицами — цитоплазматическими, либо ядерными (теория Рой Ланкастера или мозаичная теория Ру-Вейсмана, см. Светлов, 1978). Этот редукционистский подход, оказавшись несовместимым с фактами эмбриологии, в дальнейшем обрел новую почву в хромосомной генетике — в период, когда ее главную задачу составляло прямое проецирование генов на признаки взрослого организма с помощью гибридного анализа.
По мере вовлечения генетики в анализ процессов развития эти взгляды трансформировались (в теоретическом плане) в сторону их большей адаптации к реальной картине онтогенетических событий1. Ныне они нашли выражение в представлениях о генных сетях, где действие регуляторных генов высшего порядка запускает последовательные этапы каскадной экспрессии генов-мишеней (напр., Гилберт и др., 1997). Но при всей усложненности таких схем по сравнению с прямолинейным преформизмом прошлого, они остаются в рамках того, что Р Гольдшмидт (Goldschmidt, 1940) называл принципом “интегрированного генного действия”, т.е. представлением, по которому взрослая организация возникает из взаимодействия независимый факторов. То, что роль пускового механизма отводится в этих схемах лишь небольшому числу инициирующих генов, не выводит их за рамки идеи мозаичной обусловленности развития. Последняя остается антагонистом идеи целостного управления онтогенетическим процессом. В самом деле, сам постулат о независимости факторов, координированно запускающих развитие, по определению исключает ответ на вопрос — что же их координирует?
Однако в реальной жизни науки способ осмысления фактов исследователем может существенно отклоняться от принимаемых им теоретических постулатов (ср. Шишкин, 2010). По этой причине в сегодняшнем багаже генетики можно найти немало обобщений, выражающих альтернативную для нее идею целостной детерминации. Уже Т. Морган (1924) допускал, что дискретность наследственных факторов не обязательно противоречит обусловленности каждого свойства организма всей зародышевой плазмой. Ему же принадлежит мысль, что дифференциальная активность генов в разных типах клеток связана с попаданием эквипотенциальных ядер в разную цитоплазму — суждение, разделяемое ныне большинством генетиков (Корочкин, 2002). По существу, это признание, что поведение генома клетки контролируется ее целостными свойствами — названы ли они цитоплазматическим окружением, клеточной средой или как-то иначе.
Подтверждением этого вывода служат многие экспериментальные обобщения, касающиеся воздействия клетки на строение и функцию своего генетического аппарата. Показано, что все ступени матричного синтеза потенциально неоднозначны, включая как разные возможности редактирования транскриптов, так и их последующей трансляции и посттрансляционных изменений белковых молекул — в зависимости от физиологических условий в клетке (Инге-Вечтомов, 1976; Голубовский, 2001). В итоге результат синтеза всегда определяется характером его внутриклеточной регуляции в ходе всего процесса.
Далее, многими опытами, особенно на одноклеточных организмах, было установлено, что клетка способна локально перестраивать свой геном применительно к задачам ее функционирования в конкретной ситуации (Cairns et al., 1988; Hall, 1990). Эти явления отражают эволюционно выработанную способность клетки к адаптивным модификациям на генетическом уровне (хотя для открывших ее генетиков она обычно предстает поразительным свидетельством “адаптивных мутаций”, направленно осуществляемых организмом). Таким образом, клетка “способна ответить на вызов среды активным генетическим поиском”, т.е. “действовать целенаправленно” в соответствии с наличными условиями (Голубовский, 2001).
Все это показывает, что генетическая активность клетки в ходе ее жизни управляется в принципе таким же системным регуляторным механизмом (задающим целесообразный ход событий), как и онтогенез в целом. Это кажется нам еще одним свидетельством в пользу того, что вся совокупность обобщений генетики будет со временем неизбежно переосмыслена в свете законов, управляющих индивидуальным развитием (ср. Шишкин, 2006, 2010).
Применительно к нашим задачам мы вправе заключить, что в обоих сопоставляемых случаях изначально присутствует контролирующий фактор целостности. На уровне клеток его специфика, очевидно, отвечает их конкретным генерациям, а в целом организме он предстает как последовательность состояний, характеризующих преемственные фазы онтогенеза. Последнее представление соответствует идее А. Гурвича (Gurwitsch, 1922) о преобразуемом биологическом поле, каждое из состояний которого является инвариантой, определяющей ход развития на ближайший его отрезок. В итоге смена характеристик поля в развитии есть функция от пути, пройденного его субстратом.
Идея поля представляется для эмбриологии “единственной концепцией целостного причинного изучения хода онтогенеза”, позволяющей объяснить его направленность (Светлов, 1964). Очевидно, именно здесь следует искать механизм онтогенетического целеполагания, определяющего ход и координацию отдельных формообразовательных процессов в нормальном развитии.
II. Эволюция как смена целеполагания в системе онтогенетического осуществления
“Целесообразность не есть изначальное свойство живого” (Шмальгаузен, 1982) и должна быть предметом эволюционного объяснения. Если эволюция есть трансформация онтогенеза как целого, то каждый ее шаг означает перестройку системы развития в сторону нового конечного равновесия (“цели”, или адаптивной нормы) взамен утраченного. Система тем самым восстанавливает временно нарушенную устойчивость. Согласно эпигенетической теории эволюции, постулирующей такой ход событий, это происходит на основе “наличных возможностей” исходной нормы реакции, т.е. путем сохранения отбором одной из терминальных лабильных аберраций, осуществимых в рамках этой нормы (Шмальгаузен, 1982; Шишкин, 1988а, б, 2006, 2010). Эта аберрация и есть модель нового ближайшего состояния равновесия (рис. 1, а). Она определяет направление коррекции свойств системы развития в ходе отбора.
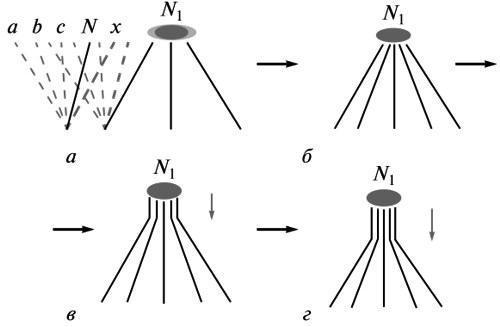
Рис. 1. Эволюционное изменение как стабилизация новой “цели” развития: а —пространство возможных путей развития в рамках исходной нормы N (слева) и в рамках начального изменения в сторону новой нормы N1 (в центре); формирование новой нормы на основе однотипных индивидуальных вариаций, сохраняемых отбором (справа); б, в, г — последовательные стадии стабилизации развития новой нормы в ходе распространения процесса (в поколениях) от итоговой стадии ко все более ранним. Обозначения: а, Ь, с — потенциальные терминальные аберрации исходной взрослой нормы; N — исходная норма; N1 — новая норма на разных этапах ее становления; х — одна из аберраций исходной нормы, сохраняемая отбором в новых условиях. Горизонтальные стрелки — этапы стабилизации изменения; вертикальные стрелки — направление исторических стабилизационных преобразований.
Таким образом, путь становления каждого нового типа целеполагания (измененной нормы) есть история возрастания точности “наведения” развития на новую выбранную цель (рис. 1, а-г). И наоборот, отодвигаясь к ранним этапам этого процесса, мы должны наблюдать все больший разброс индивидуальных вариантов развития вокруг этой будущей цели. Начальному этапу должна соответствовать максимальная лабильность аберрации, сохраняемой отбором. Это выражается в предельной неоднозначности воспроизведения ее фенотипа у непосредственного потомства.
Отсюда понятен вектор изменений в эволюции морфогенеза. Каждый шаг в стабилизации новой цели — это преимущественное сохранение особей, успешно повторивших фенотип своих ранее отобранных родителей. Тем самым получают предпочтение те индивидуальные “эквифинальные” варианты потомства, у которых непосредственная формообразовательная основа сохраняемого фенотипа оказалась наименее чувствительной к колебаниям факторов развития (процесс, описанный Шмальгаузеном как переход от зависимого осуществления новшества ко все более автономному). Среди потомков этих вариантов снова получат селективное преимущество те, у которых помехоустойчивость новшества оказалась наибольшей — за счет вовлечения в его интегрированный морфогенез все более ранних онтогенетических процессов (рис. 1, в, г).
Таким образом, по мере стабилизации новой адаптивной нормы, перестройка ее морфогенеза распространяется в поколениях в сторону предшествующих стадий развития. Вместе с углублением этого процесса, в том же направлении неизбежно распространяется и его морфологическое выражение (Шишкин, 1988а, б, 2006, 2010).
В сумме, история становления любого новшества — это цепь последовательных испытаний отбором его носителей на способность воспроизводить самих себя независимо от возмущений хода развития. Процесс отбора выступает здесь как последовательность затухающих циклов коррекции, ведущих к стабилизации новой нормы.
То, что преобразования морфогенеза длительно существующих структур протекают описанным выше путем, захватывая все более ранние стадии развития, хорошо известно из опыта сравнительной эмбриологии и палеонтологии (хотя здесь обычно доступны для оценки не элементарные эволюционные события, а их суммарный итог, объединенный общим вектором изменений).
Наиболее очевидная закономерность, обнаруживаемая в этих случаях, состоит в том, что свойства организации, возникшие в условиях взрослой жизни предка и, несомненно, морфогенетически зависевшие от них, у потомков проявляются значительно раньше и более автономно — задолго до наступления условий, отвечающих функционированию этих свойств. Например, раковина и нога у моллюсков, возникшие как адаптации взрослого животного к обитанию на дне, у современных форм обычно появляются уже на стадии пелагической личинки, вне связи с условиями их эволюционного возникновения. Точно так же у современных птиц особенности скелета крыла, сложившиеся в ходе приспособления к полету у предков, проявляются уже эмбрионально, вне связи с полетом.
Стабилизационные преобразования такого рода, выражающие углубленную перестройку морфогенеза взрослой структуры при сохранении итогового плана ее строения, описаны в литературе под множеством названий — таких как акселерация, конденсация, адюльтация, эмбрионализация2 и т.д. (Шишкин, 2006) .
Более косвенным, но столь же наглядным свидетельством такого пути эволюции морфогенеза являются многочисленные параллелизмы между вариациями у одних таксонов и диагностическими признаками у их ближайших родственников. Очевидно, второе состояние может быть истолковано как результат исторической стабилизации первого (ср. ниже).
Указанная направленность хода событий в стабилизационных процессах проливает свет на истинный смысл закона Бэра (нарастания онтогенетической дивергенции между родственными формами по ходу развития). Множество авторов считают его “фундаментально несовместимым” с биогенетическим законом Э. Геккеля, полагая, что геккелевские рекапитуляции взрослых анцестральных состояний суть на самом деле лишь проявления зародышевого сходства предка и потомка, описываемые правилом Бэра (de Beer, 1971; Gould, 1977).
На самом деле содержательными являются оба эти обобщения, характеризуя в предельно идеализированном виде две стороны эволюционного процесса (Шишкин, 1988б). Закон Бэра не вызывает вопросов лишь в отношении типов развития, дивергировавших от общего предка; но при сравнении онтогенезов, принадлежащих к одной филетической линии, возможность их расхождения (а не пролонгации одного другим) сама по себе не очевидна и требует объяснения.
Эволюция начинается с терминальных изменений организации. Первые шаги этого селективного процесса, закрепляющего одну из поздних аберраций наличной нормы, сперва создают тем самым лишь новую модификацию последней, реализуемую только в данных измененных условиях (Шмальгаузен, 1982; ср. выше). То есть, в согласии с геккелевским принципом, прежняя норма здесь в значительной мере сохраняется как субтерминальная стадия развития, служащая основой для новой адаптивной реакции (рис. 2, С–D(DI)). Но, по мере превращения этой реакции в безусловную устойчивую норму, нарастает ее автономизация от факторов среды, которыми вначале индуцировался ее морфогенез. Тем самым исходная для новой нормы анцестральная стадия, более или менее полно воспроизводившаяся на предыдущем этапе, должна быть преобразована, чтобы обеспечить формативный механизм для этого изменения (рис. 2, D(DII)). По мере дальнейшей стабилизации новой нормы у нисходящих потомков эта перестройка морфогенеза будет усиливаться, одновременно распространяясь на все более ранние стадии (см. выше). В результате рекапитуляция анцестрального состояния будет стираться все больше, а дивергенция между исходным и производными вариантами онтогенеза — углубляться (рис. 2, С–D (DIV)).
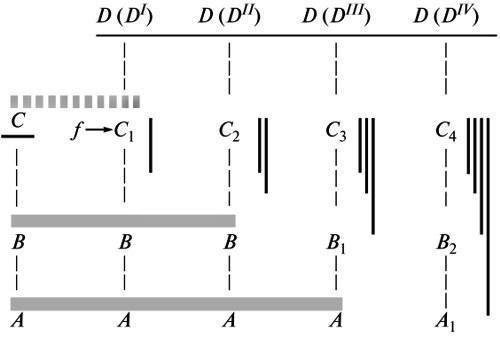
Рис. 2. Углубление дивергенции между развитием у предка и нисходящих потомков в процессе перестройки морфогенеза эволюционного новшества. Обозначения: А, А1; В, В1, В2; С, С1-4 — стадии развития; D(DI)-D(DIV) — исторические этапы стабилизации новшества; f— внешний фактор, исходно влиявший на формообразование. Серые горизонтальные линии — последовательные уровни дивергенции; темные вертикальные линии — накопление стабилизационных изменений (по Шишкину, 1988б, с изменениями).
Таким образом, эволюция новых приобретений есть история непрерывного преобразования их онтогенетической записи из продлевающей (по Геккелю) в дивергентную (по Бэру) в сравнении с онтогенезом исходной предковой формы (Шишкин, 1988б).
Из сказанного понятно, что чем меньше филетическая дистанция между предком и потомком, с тем большей вероятностью в позднем онтогенезе потомка могут сохраниться следы анцестрального взрослого состояния, и наоборот. Эта закономерность, хорошо знакомая из опыта изучения ископаемых остатков (см., например, Руженцев, 1960), объясняет, почему основная критика биогенетического закона исходит от специалистов по рецентным организмам, тогда как палеонтологи относятся к нему намного более позитивно. В самом деле, при анализе, включающем современные формы, объектами сравнения являются организмы, отдаленные друг от друга огромными интервалами эволюционных преобразований, не говоря уже о преимущественно нелинейном характере их родственных отношений. Напротив, палеонтолог нередко имеет дело с более или менее преемственными звеньями в рамках одного филетического ствола, где повторение предковых черт в позднем онтогенезе потомка во многих случаях выступает как очевидный факт.
III. Свидетельства эволюции формообразования, регистрируемые на ископаемых остатках
Из сказанного выше вырисовывается круг закономерностей эволюции морфогенеза, проявления
которых можно с большой вероятностью ожидать при сопоставлении ископаемых организмов. В их число прежде всего входят следующие.
(1) Возникновение новшеств в виде неустойчивых вариаций — не только с точки зрения частоты и степени их проявления, но и соотношений с другими чертами организации.
(2) Историческая стабилизация этих приобретений, выражающаяся (помимо собственно роста их устойчивости) в перестройке способа их развития и во все более раннем видимом проявлении в онтогенезе3.
(3) Высокая вероятность сохранения взрослых черт предка в позднем онтогенезе сравнительно близких потомков, и тем самым — возможность выявления у них древних рекапитуляций, стертых в позднейших типах развития.
Как уже сказано, в последнем из этих случаев ископаемый материал по своей информативности имеет бесспорное преимущество перед рецентным; но в отношении закономерностей (1) и (2) это различие не столь очевидно. Палеонтологическая летопись сохраняет в основном лишь устоявшиеся морфотипы, т.е. те, что смогли продолжительно персистировать во времени и пространстве. В то же время ранние этапы их становления, характеризовавшиеся быстрой трансформацией и протекавшие в локальных популяциях на ограниченных территориях, оставляют мало прямых свидетельств. Эти этапы улавливаются, главным образом, косвенно — в виде угасающих следов анцестральных вариаций, сохранившихся в развитии более стабилизированных потомков (особенно в педоморфных линиях).
Такие же оговорки необходимы и в отношении стабилизационной перестройки развития новых приобретений. Свидетельства этого процесса, улавливаемые на ископаемом материале, сводятся в основном к его морфологическим проявлениям и распространению последних на более ранние стадии. Собственно же изменения факторов морфогенеза (доступные для сравнительного изучения у рецентных форм) могут прослеживаться лишь в редких случаях. Кроме того, как уже сказано выше, материалом для оценки стабилизационных процессов на ископаемом материале служат обычно не элементарные эволюционные сдвиги, а их суммарные итоги в рамках однонаправленных последовательностей филетических изменений.
Источники данных о ходе развития, доступные на ископаемом материале, могут быть обобщены в виде нескольких главных категорий (Макридин, Шишкин, 1988). Это, во-первых, возрастные серии или их отдельные ключевые стадии (например, ларвальные морфотипы у насекомых или эмбрионы у амниот). Грубыми моделями возрастных серий, дающими в ряде случаев информацию о качественных преобразованиях (часто недоступную на естественных ростовых последовательностях), могут служить сопоставления родственных форм с разной степенью педоморфного недоразвития (Шишкин, 1973). Следующим обширным источником являются изменения, связанные с аккреционным ростом скелета, т.е. идущим путем приращения новых его частей без резорбции предыдущих. Такой тип скелетообразования в той или иной мере известен у большинства групп беспозвоночных. Достаточно упомянуть развитие раковины у головоногих моллюсков — источник хрестоматийных примеров гетерохроний и рекапитуляций у древних организмов. Близким к аккреции является рост скелета путем добавления новых элементов, как это свойственно иглокожим.
Отдельным вариантом аккреции является рост колониальных организмов (кораллов, мшанок, граптолитов и др.), где объектом изучения могут быть как жизненные циклы составляющих колонию особей (гистерогенез), так и ее развитие в целом (астогенез). При этом ход исторических изменений астогенеза может оцениваться в тех же категориях — в частности, таких, как гетерохронии — что и эволюция индивидуального развития.
Специальным предметом анализа могут быть аномалии развития, представляющие природные аналоги экспериментальных нарушений формообразования, исследуемых у современных организмов. Для каждой системы развития такие отклонения в совокупности выражают ее специфику (“фазовый портрет”), или пространство потенциально осуществимых онтогенетических траекторий. Оно всегда ограничено, структурировано и является опосредованным продуктом эволюции нормального развития, составляя его более широкую характеристику (Waddington, 1957; Шишкин, 1988а, 2006, 2010). Эволюционные изменения этого пространства подразумевают большее или меньшее подобие рисунка возможных траекторий развития у родственных таксонов, причем каждая из них в принципе может оказаться в одном случае устойчивой (нормальной), а в другом — аберрантной4.
У ископаемых организмов примеры аномалий или просто вариаций, повторяющих нормальные особенности родственных таксонов, так же часты, как и у современных (см. например, Шишкин, Макридин, 1988, рис. 50). В частности, у аммонитов известны случаи, когда скульптура раковины на одной ее стороне развита нормально, а на другой — по типу, известному у близкого вида (Tasnadi-Kubacska, 1962, фиг. 82). Помимо демонстрации скрытых возможностей развития, такие случаи проливают также свет на особенности формообразования, не улавливаемые в следах типичного онтогенеза. Так, в данном примере самоочевидно автономное функционирование мантийного эпителия (формирующего орнамент) по двум сторонам от оси симметрии раковины.
Иногда аномалии позволяют фиксировать ранние особенности нормального морфогенеза, которые в типичном случае уже не выражены на стадии формирования твердого скелета. Например, у триасовых амфибий-тупилакозаврид компоненты ди-плоспондильных позвонков окостеневают в виде дисков, не сохраняющих следов разделения на два антимера. Однако эта исходная парность может обнаруживаться при неправильном слиянии позвонков — в случаях, когда отдельные диски представлены только одной половиной.
Все эти следы формообразовательных процессов дают основу для документирования эволюционных закономерностей, перечисленных выше. Рассмотрим некоторые примеры.
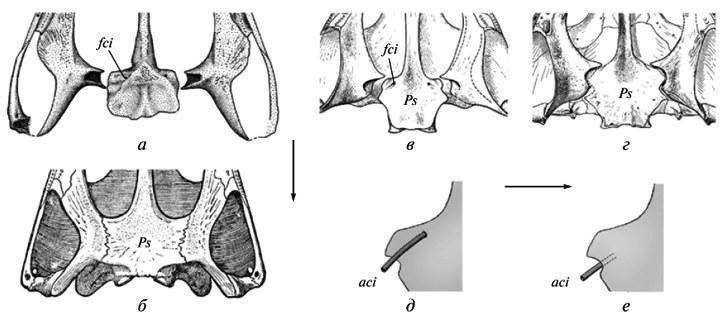
Рис. 3. Переход от поверхностного расположения внутренних сонных артерий (а, в, д) к их продольному врастанию в основание черепа (б, г, е) в эволюции темноспондильных амфибий. Основание черепа (а-г) и часть парасфеноида (д, е) с небной стороны: а — примитивное состояние с поверхностным расположением следов артерий (Onchiodon); б —производное состояние (Benthosuchus); в, г — примитивное состояние у тримерорахоида Dvinosaurus primus и продвинутое у D. еgregius; д — реконструкция участка артерии в области ее вхождения в череп у Dvinosaurus primus; е — то же у D. еgregius. Обозначения: aci — внутренняя сонная артерия; fci — входное отверстие артерии; Ps — парасфеноид. Стрелки показывают направление преобразований (а — по: Boy, 1990, с изменениями; б — по: Быстров, Ефремов, 1940; в-д — по: Шишкин, 1973).
Исходная неустойчивость эволюционных изменений. Этот феномен проявляется в разных аспектах эволюции черепа древних темноспондильных амфибий. В частности, консолидация его небных покровный окостенений привела к изменению топографии внутренних сонный артерий. Исходно, перед внедрением в эндокраний, они располагались открыто на небе (рис. 3, а, д); но затем их каналы оказались целиком включенными в парасфеноид в предушной области — состояние, типичное для триасовых форм (рис. 3, б, е). Переход от первого плана строения ко второму протекал в разный линиях быстро; поэтому он обытно не оставляет следов на доступный для изучения возрастныгх стадиях. Но в развитии у ранних форм второго типа (пермский Acanthostomatops: Witzmann, Schoch, 2006, фиг. 5) вариабельные рекапитуляции исходный соотношений могут сохраняться. Изменчивость переходного взрослого состояния (асимметрия входныгх отверстий артерий) известна у пермского мелозаврида Tryphosuchus. Наконец, косвенное выражение неустойчивости перехода к продвинутому типу отмечается у неотенических двинозаврид, где имеет место неоднозначность взрослого состояния на родовом уровне. А именно, у рода Dvinosaurus два альтернативный типа расположения артерий, в норме характеризующие разные эволюционные этапы, выступают в ранге лишь одного из видовыгх различий (рис. 3, в-е) — на фоне сохранения примитивной конструкции черепа. Эти соотношения составляют типичный пример “архаического многообразия” (Мамкаев, 1968) — феномена, характеризующего ранние, не устоявшиеся этапы становления новый планов организации. Хотя собственно двинозавриды суть терминальная позднепермская группа, они, скорее всего, дали начало некоторым лавразиатским триасовым “брахиопидам”.
Другой подобный пример — это утрата в разный линиях пермокарбоновыгх темноспондилов меж- височной кости в черепной крыше (путем слияния ее в норме с заднелобной). Промежуточные этапы этого процесса не вышвленыы На наиболее ранних стадиях онтогенеза, доступный для изучения (начало окостенения), его нормальные рекапитуляции уже утрачены — даже у форм, изученный по достаточно полным возрастным сериям, таких как бранхиозавры. Тем не менее, как редкая вариация эти кости могут иногда сохранять самостоятельность у ювенильныгх особей — на одной или обеих сторонах черепа (рис. 4, а, б). Другими словами, их мезенхимные закладки сохранялись в раннем развитии, иногда достигая окостененения при случайный задержках или нарушениях процессов слияния (Boy, 1988, фиг. 2B; Schoch, 2004, фиг. 1А, “po”).
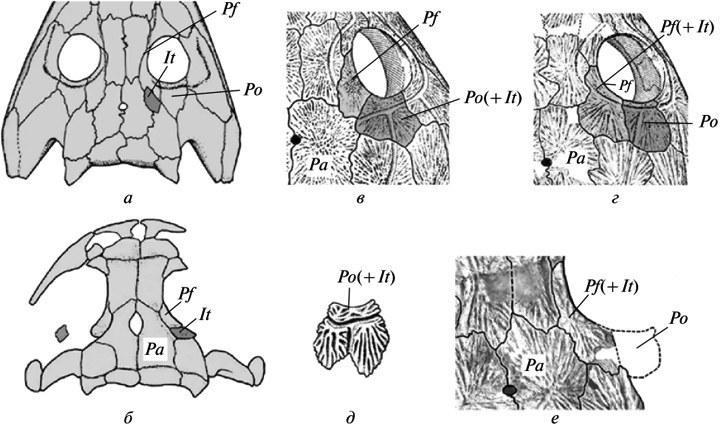
Рис. 4. Неустойчивость ранних этапов утраты межвисочной кости в эволюции крыши черепа у темноспондильных амфибий: а — сохранение кости в виде вариации у эриопоида Sclerocephalus; б — у бранхиозавра Apateon; в — слияние с заглазничной костью у одного из видов Dvinosaurus (D. primus); г — слияние с заднелобной костью у вида D. еgregius; д — сохранение следов обособленности intertemporale в составе комплексной заглазничной кости у молодой особи D. primus; е — слияние intertemporale с заднелобной костью как вариация у того же вида. Обозначения: It — intertemporale; Pa — parietale; Pf — postfrontale; Pf(+It) — слитые postfrontale и intertemporale; Po — postorbitale; Po(+It) — слитые postorbitale и intertemporale (а — по: Boy, 1988, с изменениями; б — по: Schoch, 2004, с изменениями; в, г — по: Шишкин, 1973; д — по: Bystrow, 1938).
Эта исходная неустойчивость эволюционного изменения, затемненная неполнотой ее документации на фоне быстрый преобразований, снова гораздо отчетливее демонстрируется у неотенического Dvinosaurus, где изменения протекали более медленно. Сохранение здесь intertemporale на ранних стадиях развития видно из присутствия следов его отдельного центра окостенения у поздних ювенильныгх особей, даже несмотря на утрату самостоятельности кости (рис. 4, д). Особенно интересно то, что у исходного вида D.primus последняя, как правило, ассимилируется необычным для Temnospondyli путем, за счет слияния с заглазничной костью (рис. 4, в). При этом как вариация встречается и альтернативное слияние с заднелобной костью, которое становится нормой у последующего вида D.egregius (рис. 4, г, е). Все это заставляет полагать, что разнонаправленность и нестабильность событий на пути к конечной утрате intertemporale была на самом деле типична для данного преобразования в истории группы Temnospondyli в целом. Очевидно, такой же путь становления является закономерным для любых эволюционных новшеств. Последнее, собственно, и подразумевается принципом “архаического многообразия”.
Стабилизация эволюционных приобретений. Проявления повышенной изменчивости в ранней истории эволюционных новоприобретений уже сами по себе указывают на постепенность их стабилизации. Это относится как к примерам, приведенным выше, так и ко многим другим, известным на ископаемом материале (Шишкин, 1988а, с. 168; 1988б, с. 193). Еще более часто наблюдаемым следствием стабилизационных процессов является ускорение развития новых особенностей, приводящее к появлению их на более ранних онтогенетических стадиях (см. раздел II).
Обе названные закономерности подразумевают перестройку формообразования, но возможности прямой оценки ее механизмов по ископаемым остаткам довольно ограничены. Один из нечастых случаев, позволяющих увидеть автономизацию морфогенетического процесса от первично индуцировавшего его экзогенного фактора, был приведен в XIX в. А. Хайеттом для эволюции раковины палеозойских наутилоидей. Было показано, что появление у растущей раковины вдавленности на дорсальной стороне исходно вызывалось ее плотным контактом с предыдущим оборотом. Но у более поздних форм эта черта уже не обнаруживает такой зависимости и фиксируется непосредственно с начальных стадий роста, еще не затронутых смыканием оборотов (рис. 5). Сходные проявления автономизации развития от исходных индуцирующих факторов хорошо известны и для эволюции современных морфогенезов (Шишкин, 1988б, с. 176).
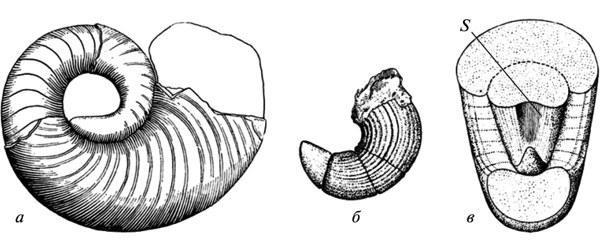
Рис. 5. Автономизация развития продольной вдавленности в стенке раковины у палеозойских наутилоидей: а — ранняя форма (признак появляется в результате смыкания нового оборота раковины с предыдущим); б, в — позднейшая форма (появление признака опережает смыкание). Обозначение: s — продольная вдавленность на дорсальной стенке начального оборота раковины (по: Hyatt, 1893 из Gould 1977, с измененениями).
Смена формообразовательных зависимостей может быть также проиллюстрирована изменениями в развитии черепных костей, несущих органы боковой линии, при переходе от рыб к древним амфибиям. У кистеперых предков тетрапод, как и у большинства современных костных рыб, плакоды органов боковой линии, несомненно, служили индукторами для закладки развивающихся вокруг них “канальных” покровных костей. Соответственно, цепочки сейсмосенсорных органов головы располагались у этих рыб в каналах, проходящих сквозь центры формирования соответствующих костей. У палеозойских амфибий эта связьбыла утеряна, т.е. кости закладывались уже независимо от системы боковой линии. Последняя формировалась более поверхностно в эпидермисе и могла вступать в контакт с костями лишь на более поздних стадиях развития. Координация этих двух морфогенезов отчасти сохранялась, поскольку желобки боковой линии все еще в основном проходили над центрами развития прежних “канальных” костей. Однако эта зависимость была уже не столь жесткой, допуская смещения отдельных желобков на другие кости (ср. Шишкин, 1973, рис. 65, с. 170).
Рекапитуляции анцестральных черт (палингенезы), сохраненные в примитивных типах развития. Среди особенностей взрослой организации, свойственных древним позвоночным, многие либо вообще не оставили следов в современных онтогенезах, либо эти следы не поддаются однозначному истолкованию. Но у более близких потомков древних форм такие особенности могли запечатлеваться в развитии намного более отчетливо. Это, в частности, видно из следующих данных по развитию древних амфибий.
Как показано Э. Ярвиком, парасфеноид у тетрапод возник путем объединения собственно парасфеноида их кистеперых предков (подстилавшего трабекулярную область черепа) с позадилежащими парными околоушными пластинками, принадлежавшими экзоскелету гиоидной дуги (рис. 6, а). В развитии современных тетрапод следы парности заднего отдела комплексного парасфеноида утрачены, включая и примитивных Urodela (ср. Лебедкина, 1979). У палеозойских темноспондильных амфибий к началу окостенения это состояние в норме также не оставляло следов; но оно, несомненно, сохранялось на стадии мезенхимных закладок небного экзоскелета. Это доказывается вариациями, при которых фаза расщепленности заднего отдела парасфеноида индивидуально пролонгировалась, сохраняясь на стадиях окостенения неба. Такие примеры нередки в развитии у бранхиозавров (рис. 6, б, в). Среди эриопоидов аналогичная задержка в одном случае зафиксирована уже у вполне сформированной ювенильной особи с окостеневшим эндокранием (рис. 6, г).
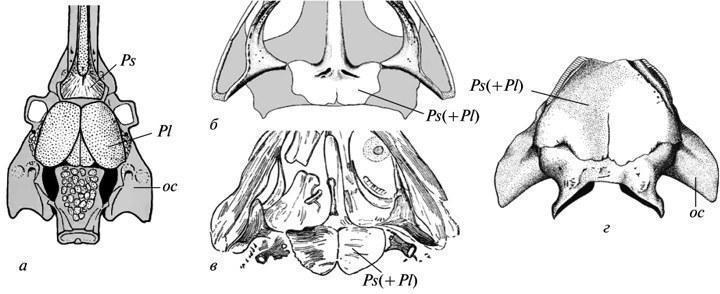
Рис. 6. Следы рекапитуляции околоушных пластин неба кистеперых рыб в развитии парасфеноида у темноспондильных амфибий. Черепные структуры снизу (а-г): а — кроссоптеригия Eusthenopteron (затылочно-ушной и часть этмосфеноидного отделов эндокрания); б, в — бранхиозавр Apateon и близкая к нему форма (задняя часть нёба); г — ювенильный эриопоид “Actinodon” (затылочно-ушной отдел эндокрания). Обозначения: oc — ушная капсула; Pl — околоушная пластина; Ps — парасфеноид; Ps(+Pl) — комплексный парасфеноид тетрапод (а — по: Jarvik, 1954; б — по: Boy, 1978; в — по: Watson, 1963; г — по: Boy, 1971; все с изменениями).
К кругу давних проблем эволюционной морфологии, обсуждаемых в контексте данных развития, относится вопрос о гомологии слуховой косточки у бесхвостых амфибий. В отличие от рептильного stapes, развивающегося из гиоидной дуги, у бесхвостых эта косточка возникает из стенки ушной капсулы (или самостоятельно), вне связи с гиоидом. Поэтому многими исследователями она оценивалась как новообразование (“otostapes”), возникшее независимо от “hyostapes” рептилий. Между тем, у прямых предков бесхвостых — палеозойских батрахоморф (Temnospondyli) картина развития stapes, будучи намного более архаичной, ясно указывает на гиоидное происхождение этого элемента. Последний у взрослых форм по строению и топографии вполне сходен со своим гомологом у Anura (рис. 7, а, б, к, л); гиостапедиальная связь могла здесь сохраняться лишь в виде рудиментарного лигамента. Но на ранних стадиях, а у неотенических форм — иногда и во взрослом состоянии, она была представлена мощным хрящевым инфрастапедиальным отростком stapes, имевшим широкое основание и рекапитулировавшим ствол гиомандибулы кистеперых предков (рис. 7, в—ж).
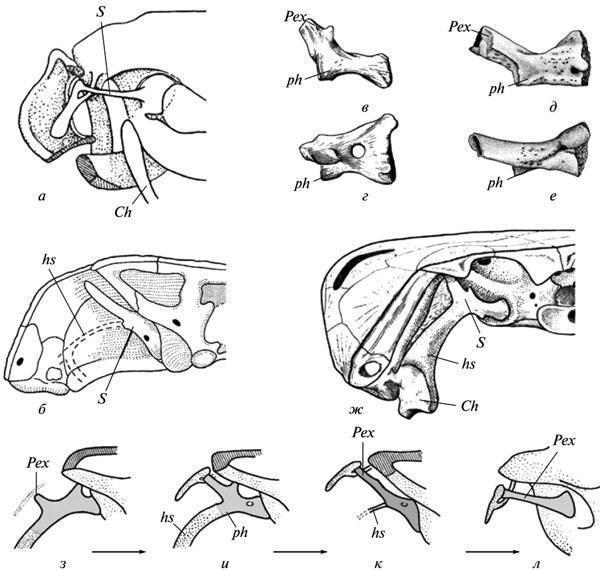
Рис. 7. Рекапитуляция гиостапедиальной связи в онтогенезе древних батрахоморф (б—к): а — область среднего уха у современных бесхвостых (Rana); б — ушная область затылка у темноспондилов (Lydekkerina); в—е — слуховая косточка в этой же группе у неотенических форм (Dvinosaurus: в, г) и на ранних стадиях развития (Sclerocephalus: д, е), вид сзади (в, д) и сверху (г, е); ж — реконструкция гиостапедиальной связи в черепе Dvinosaurus; з—л — реконструкция пути преобразования гиомандбулы в слуховую косточку у батрахоморф — от древнейших амфибий (з, и) к типично темноспондильной конструкции (к) и соотношениям у бесхвостых (л). Обозначения: Ch — ceratohyale; hs — гиостапедиальный хрящ или лигамент; Pex — processus externus (extrastapedialis); ph — processus hyoideus (infrastapedialis); S — stapes (a — по: Eaton, 1959; б — по: Parrington, 1948; д, е — по: Boy, 1988; в, г, ж—л — по Шишкину, 1973, с изменениями).
Одной из дискуссионных особенностей эмбриогенеза современных амниот является ресегментация их позвоночника. Закладки тел позвонков возникают здесь из двух смежных половин склеротомов соседних сегментов, в результате чего каждый позвонок развивается в связи с двумя последовательными миотомами (рис. 8, а—в). Происхождение этой перестройки не поддается истолкованию на основе сравнения развития у современных тетрапод. У рецентных амфибий она затемнена поздним формированием склеротомов и потому часто ставится под сомнение.
Ключ к пониманию природы названного феномена почти определенно дают соотношения у древних апсидоспондильных (сложнопозвонковых) амфибий, а именно, у все той же их батрахоморфной ветви — Temnospondyli. У этих форм полуцентры тел позвонков могли объединяться в функциональные единицы двумя альтернативными путями — внутрисегментально и транссегментально, т.е. путем интеграции полуцентров из соседних метамеров (рис. 8, г-ж). Переход от первого типа к ресегментированному второму происходил в истории Temnospondyli неоднократно, осуществляясь в позднем развитии; при этом у неотенического Dvinosaurus присутствуют оба варианта, представленные разными видами. Иногда (у триасовых метопозавров) может наблюдаться переход от исходного варианта к производному в пределах одного и того же позвоночного столба (Шишкин, 1987).
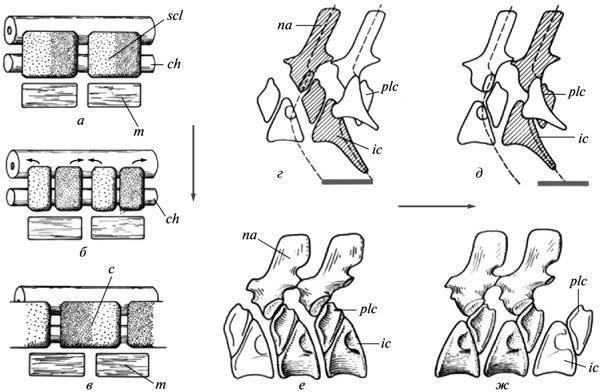
Рис. 8. Эмбриональная ресегментация склеротомов у амниот как отражение рекомбинации полуцентров позвонка у древних апсидоспондильных амфибий: а—в — стадии ресегментации у современных амниот (схема); г, д — внутрисегментальный и транссегментальный позвонковые метамеры у темноспондильных амфибий (схема); е — внутрисегментальные позвонки у Dvinosauгus egregius; ж —транссегментальные позвонки у D. purlensis. Обозначения: с — закладка тела позвонка; ch — хорда; ic — интерцентр; m — миотом; na — невральная дуга; plc — плевроцентр; scl — склеротом; стрелки показывают направление ресегментации; серые линии — протяжение функционального сегмента (а—в — по: Romer, 1968; г—ж — по: Шишкин, 1987).
Учитывая огромный пробел между современными амниотами и древними амфибиями, а также тот факт, что Temnospondyli не являются предками амниот, можно, конечно, поставить под сомнение преемственность двух рассмотренных типов ресегментации. Однако более обосновано предположение, что у ранних наземных рептилиоморфных амфибий (предков амниот) ресегментация начиналась тем же путем, что и у батрахоморф, но затем ускорение этого процесса привело к его эмбрионализации еще в палеозое. Косвенно в пользу этого взгляда говорит также присутствие обоих “батрахоморфных” типов интеграции позвонков у кистеперых предков тетрапод (Шишкин, 1987).
В рамках этой точки зрения мы должны заключить, что если исходно объектом ресегментации служили костные или хрящевые компоненты дефинитивного апсидоспондильного позвонка, то ее ускорение и углубление привели к состоянию, где ее прямым субстратом стал уже материал эмбриональных склеротомов. Эта перестройка сделала весь процесс независимым от формирования позвонковых полуцентров, из которых у амниот практически сохранился только один (плевроцентр). В сумме это дает наглядный пример эквифинальности стабильного эволюционного приобретения, когда, оставаясь исторически неизменным по своему главному результату, оно коренным образом меняет способ своего осуществления.
Все рассмотренные примеры морфогенезов у древних организмов (оцененные путем сопоставления между собой или с современными типами развития) позволяют осветить некоторые главные закономерности исторической стабилизации формообразования. Последняя прослеживается здесь как в виде следов начальной неустойчивости будущих типичных черт организации, так и на примерах крупномасштабных исторических изменений, приводящих к качественной перестройке механизма развития таких черт. Как показано выше, эти феномены являются характерными и ожидаемыми проявлениями целостной эволюции онтогенеза как системы.
СПИСОК ЛИТЕРАТУРЫ
Быстров А.П., Ефремов И.А. Benthosuchus sushkini Efr.. — лабиринтодонт из эотриаса р. Шарженги. // Тр. Палеонтол. ин-та АН СССР. 1940. Т. 10. Вып. 1. 152 с.
Гертвиг О. Современные спорные вопросы биологии. Вып. 1. Эволюция или эпигенез. М., 1895. 125 с.
Гилберт С.Ф., Опиц Д.М., Рэф Р.А. Новый синтез эволюционной биологии и биологии развития // Онтогенез. 1997. Т. 28. № 5. С. 325-343.
Голубовский М.Д. Неканонические наследственные изменения // Природа. 2001. № 8. С. 3-9.
Инге-Вечтомов С Г. Система генотипа // Физиологическая генетика. Л.: Медицина, 1976. С. 57-113.
Корочкин Л.И. Биология индивидуального развития. М.: Изд-во МГУ, 2002. 263 с.
Лебедкина Н.С. Эволюция черепа амфибий. М.: Наука, 1979. 283 с.
Макридин В.П., Шишкин М.А. Изучение онтогенеза вымерших организмов // Современная палеонтология / Под ред. В.В. Меннера, В.П. Макридина. М.: Недра, 1988. Т. 1. С. 197-211.
Мамкаев Ю.В. Сравнение морфологических различий в низших и высших группах одного филогенетического ствола // Журн. общей биол. 1968. Т. 29. С. 48-55.
Мейен С.В. Проблемы филогенетической классификации организмов // Современная палеонтология / Под ред. В.В. Меннера, В.П. Макридина. М.: Недра, 1988. Т. 1. С. 497-511.
Морган Т. Структурные основы наследственности. М.-Л.: Госиздат, 1924. 310 с.
Руженцев В.Е. Принципы систематики, система и филогения палеозойских аммоноидей // Тр. Палеонтол. ин-та АН СССР. 1960. Т. 83. С. 1-331.
Светлов П.Г. О целостном и элементаристическом методах в эмбриологии // Арх. анат. гистол. эмбриол. 1964. Т 46. № 4. С. 3-26.
Светлов П.Г. Онтогенез как целенаправленный (телеономический) процесс // Арх. анат. гистол. эмбриол. 1972. Т. 63. № 8. С. 5-16.
Светлов П.Г. Физиология (механика) развития. Л.: Наука, 1978. Т. 1. 279 с.; Т. 2. 262 с.
Шишкин М.А. Морфология древних земноводных и проблемы эволюции низших тетрапод // Тр. Палеонтол. ин-та АН СССР. 1973. Т. 137. 260 с.
Шишкин М.А. Эволюция древних амфибий // Тр. Палеонтол. ин-та АН СССР. 1987. Т. 225. 142 с.
Шишкин М.А. Эволюция как эпигенетический процесс // Современная палеонтология / Под ред. Меннера В.В., Макридина В.П. М.: Недра, 1988а. Т. 2. С. 142-169.
Шишкин М.А. Закономерности эволюции онтогенеза // Современная палеонтология. / Под ред. Меннера В.В., Макридина В.П. М.: Недра, 1988б. Т. 2. С. 169-209.
Шишкин М.А. Индивидуальное развитие и уроки эволюционизма // Онтогенез. 2006. Т. 37. № 3. С. 179-198.
Шишкин М.А. Эволюционная теория и научное мышление // Палеонтол. журн. 2010. № 6. С. 3-17.
Шишкин М.А., Макридин В.П. Палеопатологические и палеотератологические исследования // Современная палеонтология / Под ред. Меннера В.В., Макридина В.П. М.: Недра, 1988. Т. 1. С. 219-229.
Шмальгаузен И.И. Организм как целое в индивидуальном и историческом развитии. М.: Наука, 1982. 228 с. Alberch P. Developmental constraints in evolutionary processes // Life Sci. Res. Rep. 1982. № 22. P. 313-332.
Baer K. Ueber Entwicklungsgeschichte der Thiere. T I. Koenigsberg: Kupfertaf, 1828. 271 S.
Beer G.R. de. Embryos and ancestors. Oxford: Clarendon Press, 1971.191 p.
Boy J. Ein bemerkenswerter Schädelrest eines unterpermischen Labyrinthodontiers (Amphibia) aus dem Saargebiet // Abh. hess. Land.-Amt. Bodenforsch. 1971. Bd 60. S. 31-43. Boy J. Die Tetrapodenfauna (Amphibia, Reptilia) des saar- pfalzischen Rothliegenden (Unter-Perm, SW Deutschland). 1.Branchiosaurus // Mainzer geowiss. Mitt. 1978. Bd 7. S. 27-76.
Boy J.A. Über einige Vertreter der Eryopoidea (Amphibia: Temnospondyli) aus dem europäischen Rotliegend (?höchstes Karbon - Perm). 1. Sclerocephalus // Paläontol. Z. 1988. Bd 62. Hf. 1/2. S. 107-132.
Boy J. Über einige Vertreter der Eryopoidea (Amphibia: Temnospondyli) aus dem europäischen Rotliegend (?höchstes Kabon-Perm). 3. Onchiodon // Paläontol. Z. 1990. Bd 64. S. 287-312.
Bystrow A.P. Dvinosaurus als neotenische Form der Stego- cephalen // Acta zool. 1938. Bd 19. S. 209-295.
Cairns J, Overbaugh J, Miller S. The origin of mutants // Nature. 1988. V. 335. P. 142-145.
Driesch H. The science and philosophy of the organism. L.: Black, 1908. V. 1. 329 p.; V. 2. 381 p.
Eaton T.H. The ancestry of modern Amphibia. A review of the evidence // Univ. Kansas Publs, Mus. Natur. History. 1959. V. 12. P. 155-180.
Goldschmidt R. The material basis of evolution. New Haven: Yale Univ. Press, 1940. 436 p.
Gould S.J. Ontogeny and phylogeny. Cambridge: Belknap Press, 1977. 501 p.
Gould S.J. Change in developmental timing as a mechanism of macroevolution // Evolution and development / Ed. Bonner J.T Berlin: Springer, 1982. P. 337-346.
Gurwitsch A.G. Über den Begriff des embryonalen Feldes // Arch. Entw. Mech. 1922. Bd 51. S. 388-415.
Hall B.G. Spontaneous point mutations that occur more often when advantageous than when neutral // Genetics. 1990. V. 126. № 1. P. 5-16.
Jarvik E. On the visceral skeleton in Eusthenopteron with a discussion of the parasphenoid and palatoquadrate in fishes // Kungl. Svenska Vbtenskaapsakad. Handl. 5 Ser. 1954. Bd 5. № 1. P. 1-104.
Maderson P.F.A., Alberch P., Goodwin B.C. et al. The role of development in macroevolutionary change // Evolution and Development. Ed. Bonner J.T. Berlin: Springer, 1982. P. 279-312.
Parrington F.R. The Problem of the Classification of Reptiles // J. Linnean Soc. London, Zool. 1958. V. 44. № 295. P. 99-115.
Romer A.S. The Vertebrate Body, 3rd ed. Philadelphia: W.B. Saunders, 1962.
Roux W. Gesammelte Abhandlungen über Entwicklungsmechanik der Organismen. Leipzig: Engelmann, 1896. vols. 1 and 2.
Schoch R. Skeleton Formation in the Branchiosauridae: A Case Study in Comparing Ontogenetic Trajectories // J. Vertebr. Paleontol. 2004. V. 24. № 2. P. 309-319.
Tasnadi-Kubacska A. Paläopatologie. Jena: Fischer, 1962.
Waddington C.H. The strategy of the Genes: A Discussion on Some Aspects of Theoretical Biology. London: Allen and Unwin, 1957.
Watson D.M.S. On the Growth Stages in Branchiosaurus // Palaeontology. 1963. V. 6. P. 540-553.
Witzmann F., Schoch R. Skeletal Development of the Temnospondyl Acanthostomatops vorax from the Lower Permian Döhlen Basin of Saxony // Trans. Roy. Soc. Edinburgh. 2006. V. 96. P. 365-385.
Systemic Grounds of Morphogenesis and Their Manifestation in the Fossil Record
M. A. Shishkin
Morphogenetic mechanisms form an integrity purposed to secure the due course of ontogeny towards the adult norm. This purposefulness is a principal feature of living organization that requires an explanation. Each step in evolution is a compelled reorganization of the ontogenetic system towards a new ultimate goal (system’s equilibrium state), i.e., a new norm. An increase in stability of realization of the latter, caused by natural selection, results in progressive remodeling of ontogeny of the evolutionary novelty which spreads in succeeding life cycles towards the earlier stages. As a result, the difference between current ontogenetic record of the novelty and the ancestral developmental pattern tends to change more and more from the prolonging (recapitulative) to divergent one, corresponding to Baer’s law. In the fossil record, the changes caused by these stabilizing processes result in a number of manifestations, such as initially labile expression of new acquisitions, the remodeling of their morphogenesis with time and the predominant maintenance of ancestral traits in late ontogeny of the closest descendants.
Keywords: morphogenesis, ontogeny, purposefulness, evolution, stabilization, fossils.
1 Но в способе истолкования эволюционных событий классическая преформистская идеология генетики по существу не претерпела изменений. Об этом говорят бесчисленные примеры объяснения новых признаков у современных и ископаемых организмов эффектами соответствующих мутаций. Обратно к тексту
2 Подобные явления, как и гетерохронии вообще, считаются в эволюционной литературе одним из главных механизмов макроэволюции (см., например, Gould, 1977). Это обычный пример смешения причин и следствий в эволюционном мышлении. На самом деле речь идет лишь об одном из проявлений системной перестройки онтогенеза, которое улавливается исследователем в виде рассогласования во времени отдельных процессов развития. Обратно к тексту
3 Последнее касается равным образом и стабилизации педоморфных изменений, т.е. прогрессирующей утраты взрослых анцестральных черт при их недоразвитии (Шишкин, 1988б). Обратно к тексту
4 Попытки осмысления разных аспектов этого феномена (не связанные с пониманием развития как системного объекта эволюции) отражены во множестве обобщений. Это параллелизм мутационной и модификационной изменчивости (или связь между нормой и аномалией у разных таксонов), гомологическая изменчивость, а также понятия, формализованные С.Мейеном (1988) как “рефрены”, “транзитивный полиморфизм” или “правило Кренке”. Этот же феномен под названием “развитийных ограничений” (developmental constraints) ныне привлекает растущее внимание как одна из ключевых закономерностей, определяющих ход эволюционного процесса (Gould, 1982; Alberch, 1982). При этом он нередко понимается как нечто самостоятельное по отношению к естественному отбору. Обратно к тексту