Шабанов Дмитро Андрійович1,
Бірюк Ольга Вікторівна1,
Коршунов Олексій Владиславович1,
Кравченко Марина Олександрівна1,2
1Харківський національний університет імені В. Н. Каразіна,
61022, Україна, Харків, майдан Свободи, 4;
shabanov@karazin.ua;
2Харківський національний педагогічний університет імені Г. С. Сковороди
ПОШИРЕННЯ РІЗНИХ ТИПІВ ГЕМІКЛОНАЛЬНИХ ПОПУЛЯЦІЙНИХ СИСТЕМ ГІБРИДОГЕННОГО КОМПЛЕКСУ ЗЕЛЕНИХ ЖАБ (PELOPHYLAX ESCULENTUS COMPLEX) У БАСЕЙНІ СІВЕРСЬКОГО ДОНЦЯ
Зелені жаби — група тварин, що заслуговує виняткової уваги дослідників і природоохоронців. Це пов’язано не лише з їх з їх відносно високою біомасоюта потужним впливом на біогеоценотичні процеси. Саме на прикладі зелених жаб можна спостерігати недостатньо вивчені механізми подолання стерильності міжвидових гібридів, міжвидової рекомбінації, неменделевського спадкування, існування незвичних типів біосистем тощо.
Середньоєвропейський гібридогенний комплекс зелених жаб, Pelophylax esculentus complex, утворюють два батьківські види, ставкова жаба, Pelophylax lessonae (Camerano, 1882) і озерна жаба, Pelophylax ridibundus (Pallas, 1771), а також їх різноманітні гібриди, до яких вживається подібна до видової назва: їстивна жаба, Pelophylax esculentus (Linnaeus, 1758).
Представники батьківських видів легко схрещуються, утворюючи життєздатних гібридів. Втім, геноми P. lessonae і P. ridibundus, хоча й складаються з однакової кількості хромосом (n=13), набули таких відмінностей, що їхня нормальна кон’югація під час мейозу стала неможливою. Саме це явище є причиною гібридної стерильності багатьох міжвидових гібридів. У різних рослин та тварин відомі два основних шляхи подолання такої стерильності: тетраплоїдизація (внаслідок якої у кожної хромосоми з’являється гомологічна їй пара) або ж перехід до клонального, тобто нерекомбінантного відтворення (різні форми нестатевого розмноження, партеногенез, гіногенез тощо). Третій шлях є найрідкіснішим: це геміклональне відтворення гібридів. У типовому випадку в зародковій лінії клітин у P. esculentus відбувається елімінація одного з батьківських геномів и ендоредуплікація (подвоєння без подальшого поділу) іншого батьківського геному, клонального (Plötner, 2005; Dedukh et al., 2015). У типовому випадку відтворення P. esculentus пов’язане зі схрещуванням гібридів з представниками того з батьківських видів, геном якого вони елімінують.
Пояснимо викладене в попередньому абзаці, застосуючи літеру L для позначення геному P. lessonae, а R — геному P. ridibundus; клональність позначимо заключенням символу геному у дужки. Гібриди виникають внаслідок гібридизації представників батьківських видів (гамети показано у прямокутних комірках: 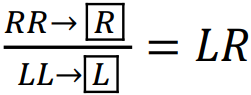 . У гібридів утворюються гамети, що несуть клональний геном:
. У гібридів утворюються гамети, що несуть клональний геном: 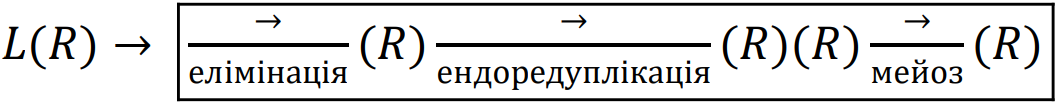 . За схрещування гібридів з представником того батьківського виду, геном якого елімінується, усе потомство складається з гібридів:
. За схрещування гібридів з представником того батьківського виду, геном якого елімінується, усе потомство складається з гібридів: 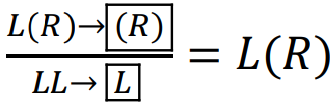 . Відтворення гібридів може таким чином відбуватися протягом багатьох поколінь. На більшій частині ареалу P. esculentus найчастіше клонально передає геноми R (Plötner, 2005).
. Відтворення гібридів може таким чином відбуватися протягом багатьох поколінь. На більшій частині ареалу P. esculentus найчастіше клонально передає геноми R (Plötner, 2005).
На частині ареалу P. esculentus зустрічаються як диплоїдні, так і триплоїдні гібриди, які можуть існувати у двох геномних композиціях: LLR і LRR. Типовим можна вважати такий варіант гаметогенезу триплоїдів, за якго вони передають у гамети той геном, що представлений у них двома копіями:  (Vinogradov et al., 1990).
(Vinogradov et al., 1990).
З викладеного зрозуміло, що відтворення P. esculentus відбувається не в популяціях будь-якого виду, а в особливих системах, де у розмноженні спільно беруть участь представники батьківських видів і різноманітні гібриди, і де з покоління до покоління передаються, крім рекомбінантних, клональні геноми. Ці біосистеми названі геміклональними популяційними системами, ГПС (Шабанов та ін., 2009). Ми застосовуємо позначення типів ГПС, де літерами позначаємо форми жаб у їх складі: L і R — батьківські види, а E — P. esculentus. Позначка p відповідає наявності в ГПС поліплоїдів (триплоїдів), а позначки f або m — випадку, коли певна форма представлена лише самицями або самцями відповідно. Якщо певна форма представлена лише однією геномною композицією, це можна позначити відповідним індексом. Наведемо приклади застосування описаних позначень: наприклад, для НПП «Гомільшанські ліси» є характерними R‑E‑Ep‑ГПС, а в НПП «Святі гори» зустрічаються R‑LLREpf‑ГПС.
Наведені приклади стосуються басейну Сіверського Донця. Вивчення зелених жаб цього регіону має довгу історію, але, на жаль, у багатьох випадках неможливо визначити, про які форми жаб йдеться у роботах, що виконувалися до розробки надійних методів ідентифікації особин. Застосування проточної ДНК-цитометрії, що дозволяє вирізняти форми жаб за розміром їх геному, привело до відкриття у басейні Сіверського Донця регіону поширення триплоїдів (Borkin et al., 2004). Невдовзі після цього було показано, що різноманіття складу ГПС цього регіону пов’язане з різноманіттям гаметогенезу поширених в ньому P. esculentus. Серед диплоїдних гібридів є такі, що передають геноми одного чи другого батьківських видів, тобто L(R) і (L)R, а також ті, що продукують суміш статевих клітин з геномами обох батьківських видів: (L)(R) (Боркин и др., 2005). Складність ГПС, описаних у Сіверському Донці, є безпрецедентною. Наприклад, на території НПП «Гомільшанські ліси» поширені:
— представники одного з батьківських видів: ♀♀ и ♂♂ RR;
— диплоїдні гібриди: ♀♀ и ♂♂ L(R), (L)R і (L)(R);
— триплоїдні гібриди: ♀♀ и ♂♂ LLR і LRR;
— поодинокі незрілі тетраплоїди LLRR і представники іншого батьківського виду, LL.
Як це не парадоксально, статевозрілі P. lessonae у басейні Сіверського Донця не знайдені (Коршунов, 2010); усі геноми L передаються тут через гібридів. З часом це мало б призвести до деградації L‑геномів внаслідок дії «храповика Мюлера», що є наслідком припинення генетичної рекомбінації. Вірогідно, це не відбувається через рекомбінацію L‑геномів під час гаметогенезу гібридів LLR (у відповідності до наведеної вище схеми).
Викладені обставини привели до опису Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex (Шабанов та ін., 2009). Цей центр складається з частин, що відрізняються між собою. Зроблено два описи географічної структури цього центру.
Перший з цих описів спирається на аналіз складу ГПС зелених жаб Лівоберіжного Лісостепу України (Шабанов, 2015). У цьому регіоні виділено 5 субрегіонів.
— I субрегіон населяють звичайні популяції P. ridibundus. До нього належать частини басейну Сіверського Донця, де немає місцеперебувань, що відповідають біогеоценотичним вимогам P. lessonae і P. esculentus, а також басейн Осколу, лівого притоку Сіверського Донця.
— II субрегіон розташований у басейні Дніпра, у тій його частині, де є придатні для P. esculentus і P. lessonae водойми, розташовані поруч з ділянками лісів. Інша частина Лівобережного Лісостепу у басейні Дніпра належить до I субрегіону. Найскладнішими в II субрегіоні є R‑E‑L‑ГПС, також зустрічаються R‑E‑ГПС, L‑E‑ГПС і популяції P. ridibundus. До його складу входить ізольований (вірогідно, реліктовий) анклав з R‑E‑L‑ГПС у заказнику «Руський Орчик» на межі Харківської, Полтавської та Дніпропетровської областей.
— III субрегіон знаходиться у басейнах річок Мжа та Уди, правих притоків Сіверського Донця, а також річки Лопань (притоку Уд) і річки Харків (притоку Лопані). Тут поширені R‑E‑ГПС і популяції P. ridibundus.
— IV субрегіон є найрізноманітнішим за кількістю зареєстрований в ньому форм жаб (які перелічені вище для НПП «Гомільшанські ліси»). Він тягнеться вздовж Сіверського Дінця до Донецької області. Найскладнішими системами в ньому є R‑E‑Ep‑ГПС. У певних ГПС втрачаються деякі форми жаб; наприклад, у ставку с. Замулівка (колишне Жовтневе) Вовчанського району мешкає R‑E‑Epf‑ГПС, де триплоїдні гібриди представлені лише самицями.
— V субрегіон розташовано нижче IV субрегіону за течією Сіверського Донця, на території Донецької та Луганської областей та, вірогідно, Ростовської області Російської Федерації. Для нього є характерними R‑LLREpf‑ГПС. Об’єктами вивчення авторів даної роботи стали ГПС околиць с. Брусівка Донецької області та м. Кремінна Луганської області, але в цьому регіоні існують й інші подібні системи (Шабанов, 2015). На території V субрегіону лишилися ділянки, що відповідають за складом ГПС зелених жаб IV субрегіону; так, у околицях с. Ясногорка поблизу м. Краматорська зареєстрована R‑E‑Ep‑ГПС (вірогідно, реліктова).
Другий опис географічної структури Сіверсько-Донецького центру різноманіття зелених жаб пов’язаний з вивченням гаметогенезу та системи схрещувань у ГПС (Biriuk et al., 2016; Бірюк, 2017). За цими ознаками виділено три групи ГПС.
— ГПС групи І складаються з P. ridibundus і диплоїдних P. esculentus. Вони відповідають III субрегіону за попередньою класифікацією (Шабанов, 2015).
— ГПС групи ІІ складаються з P. ridibundus і триплоїдних P. esculentus, та відповідають V субрегіону.
— ГПС групи ІІІ географічно розташовані між двома попередніми, складаються з P. ridibundus, ди- і триплоїдних P. esculentus та відповідають IV субрегіону за попередньою класифікацією.
Найцікавішою є система схрещування, що описана для ГПС ІІ групи, тобто R‑LLREpf‑ГПС. Самиці з генотипом LLR зазвичай продукують гаплоїдні яйцеклітини L (Dedukh et al., 2015). Вони схрещуються з самцями P. ridibundus, що продукують сперматозоїди з генотипом R. Зроблено припущення (Biriuk et al., 2015), що триплоїдні зиготи виникають внаслідок подвоєння L-геному, яке відбувається після запліднення. Можливі й інші пояснення зареєстрованого феномену. Серед них (крім, звісно, наявності в цих ГПС незареєстрованих P. esculentus, що продукують диплоїдні гамети), блокування останнього поділу мейозу і відділення яйцеклітинами полярного тільця, або злиття яйцеклітини з полярним тільцем після запліднення.
Під час опису субрегіонів Лівобережного Лісостепу України за складом ГПС зелених жаб вказано, що до складу Сіверсько-Донецього центру різноманіття Pelophylax esculentus complex входять III і IV субрегіони (Шабанов, 2015). Центром різноманіття є територія, на якій реєструється підвищена, порівняно з іншими територіями, кількість форм групи, що розглядається (Шабанов та ін., 2009). Зараз, на наш погляд, можна вважати, що незвичайний характер відтворення триплоїдів в ГПС V субрегіону дає підставу також включати цей субрегіон до складу Сіверсько-Донецього центру різноманіття, адже тут відтворення зелених жаб відбувається більшою кількістю способів, ніж в інших місцях. Включення III субрегіону до складу цього центру виправдовується, на наш погляд, його наближеністю (і географічною, і за складом диплоїдних P. esculentus) до IV субрегіону (Шабанов, 2015) і різкою відмінністю від ГПС зелених жаб, що населяють басейн Дніпра (Mezhzherin et al., 2010).
Ми пропонуємо поєднати два перелічених описи і виділити в складі Сіверсько-Донецього центру різноманіття Pelophylax esculentus complex три субрегіони (рис. 1).
Походження усього центру різноманіття пов’язано, вірогідно, з R‑E‑Ep‑субрегіоном. Позначення цього (й інших) субрегіонів показує не те, якими ГПС населена уся їх територія, а те, які ГПС населяють сприятливіші місцеперебування. Поширені тут форми жаб могли потрапити на територію України з Білгородської області РФ, спускаючись за течією Сіверського Донця. Підставою для такого припущення є те, що в Білгородській області було зареєстровано E‑ГПС (Лада, 1998), які були також описані в R‑E‑Ep‑субрегіоні. На жаль, E‑ГПС в R‑E‑Ep‑субрегіоні (ГПС Іськова ставку і Коряков яру) перейшли в інший стан; вивчення їх трансформацій є цікавою задачею (Макарян та ін., 2016). Важливою проблемою вивчення цього субрегіону є встановлення механізмів збереження значної кількості L-геномів і підтримання відносно постійної частки триплоїдних гібридів в R‑E‑Ep‑ГПС.
R‑E‑субрегіон розташований у басейнах Мжи і Уд. Склад його популяційних систем вивчався, крім іншого, за допомогою імітаційного моделювання (Кравченко, Шабанов, 2010; Шабанов, 2015). Показано, що характерні для нього ГПС можуть підтримувати стійкість завдяки відмінностям внутришньопопуляційних онтогенетичних стратегій жаб, що їх утворюють (Шабанов и др., 2014; Усова и др., 2015; Shabanov et al., 2015).
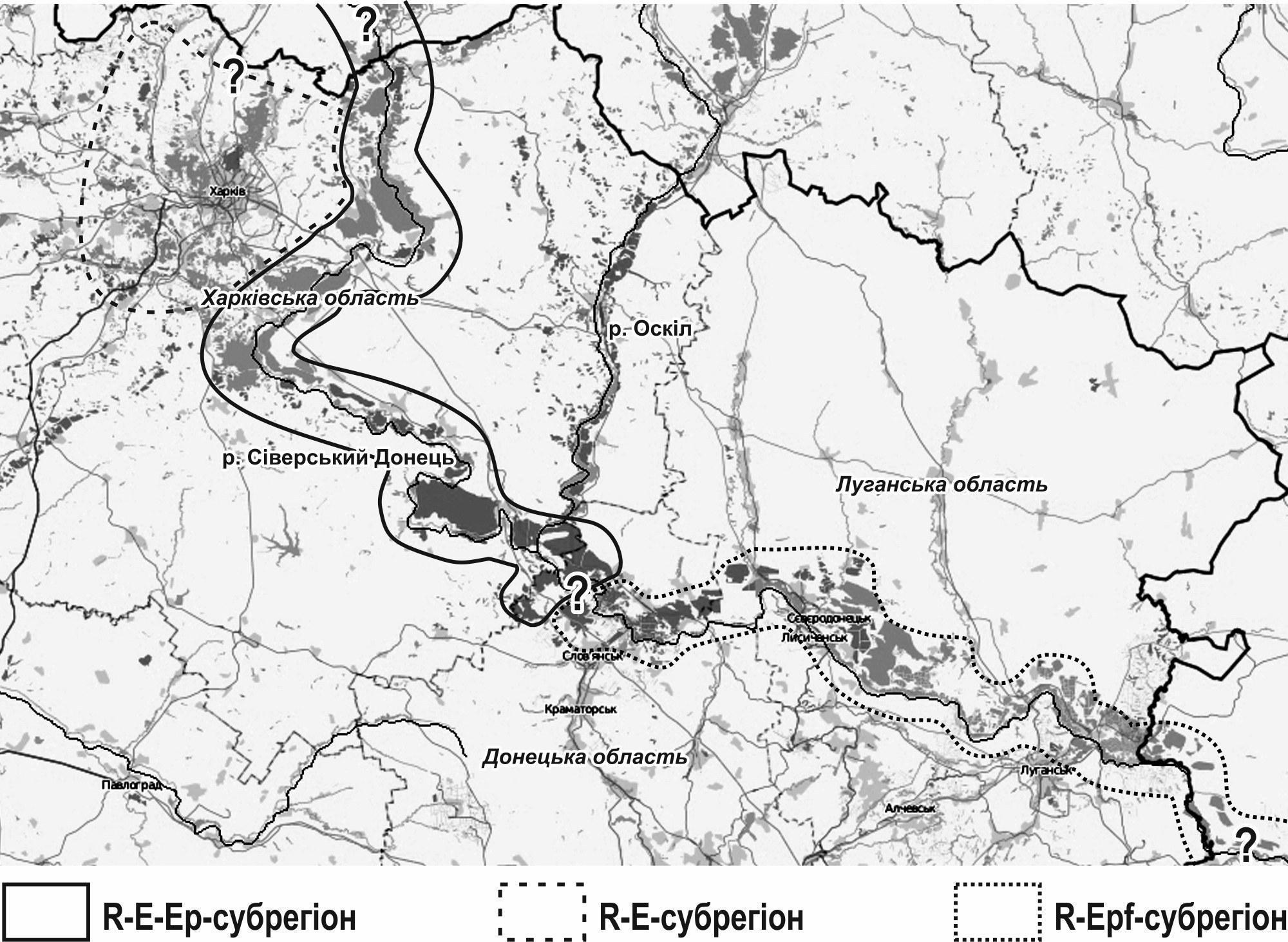
Рис. 1. Субрегіони Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex, гібридогенного комплексу зелених жаб
R‑Epf‑субрегіон розташований на території Донецької та Луганської областей. В ньому зустрічаються не лише R‑LLREpf‑ГПС, а й R‑E‑Epf‑ГПС і, природньо, популяції P. ridibundus. Можна припустити, що R‑Epf‑субрегіон виходить за межі України на територію РФ, оскільки триплоїдні самиці LLR були знайдені і в Ростовській області (Borkin et al., 2006). Зрозуміло, що R‑LLREpf‑ГПС поширені не на усій показаній на рис. 1 території, а лише у сприятливих місцеперебуваннях
Слід підкреслити, що визначення триплоїдних самок P. esculentus за зовнішніми ознаками є ненадійним (Коршунов, 2010); крім високотехнологічних методів для підтвердження діагностики триплоїдів можна застосовувати визначення розміру еритроцитів (Бондарева и др., 2012). Беззаперечно, що встановлення механізмів відтворення триплоїдних гібридів в цьому регіоні є актуальною науковою проблемою. Самі R‑Epf‑ГПС є унікальними і недостатньо вивченими природними біосистемами і потребують ретельної охорони. За нашими даними (Коршунов, 2010), важливішою умовою збереження незвичайних ГПС зелених жаб є підтримання характерних для них біогеоценозів, які включають водойми, де відтворюються жаби, та розташовані поруч з ними лісові ділянки.
Бірюк О. В. Цитогенетичні особливості сперматогенезу у диплоїдних і триплоїдних геміклональних міжвидових гібридів Pelophylax esculentus complex: автореф. дис. … канд. біол. наук за спец. 03.00.15 – генетика. – Київ, 2017. – 22 с.
Бондарева А. А., Бибик Ю. С., Самило С. М., Шабанов Д. А. Цитогенетические особенности эритроцитов зеленых лягушек из Северско-Донецкого центра разнообразия Pelophylax esculentus complex // Вісн. ХНУ ім. В. Н. Каразіна. Сер.: біол. – 2012. –15 (1008) – С. 116–123.
Боркин Л. Я., Зиненко А. И., Коршунов А. В. и др. Массовая полиплоидия в гибридогенном комплексе Rana esculenta (Ranidae, Anura, Amphibia) на Востоке Украины: Мат. Першої конф. Українського Герпетологічного Товариства – К.: Зоомузей ННПМ НАНУ, 2005. – С. 23–26.
Коршунов О. В. Екологічні закономірності розподілу Pelophylax esculentus complex у біотопах басейну верхньої течії ріки Сіверський Донець: автореф. дис.... канд. біол. наук за спец. 03.00.16 – екологія. – Дніпропетровськ, 2010. – 20 с.
Кравченко М. А., Шабанов Д. А. Моделирование трансформаций гемиклональных популяционных систем зеленых лягушек (Pelophylax esculentus complex; Amphibia, Ranidae) с помощью рекуррентных разностных уравнений // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2010. – Вип.12 (№920). – С. 70–82.
Лада, Г. А. О необходимости сохранения уникальных «чистых» популяций диплоидной съедобной лягушки (Rana esculenta Linnaeus, 1758) в Белгородской и Харьковской областях: Пробл. охраны и рац. исп. природн. экосистем и биол. ресурсов. – Пенза, 1998 . – С. 333–335.
Макарян Р. М., Бірюк О. В., Коршунов О. В. та ін. Склад пуголовків зелених жаб (Pelophylax esculentus complex) в Іськовому ставі (НПП «Гомільшанські ліси»): Стан і біорізноманіття екосистем Шацького НПП та інших природоохор. територій. – Львів: СПОЛОМ, 2016. – С. 61-65.
Усова Е. Е., Кравченко М. А., Шабанов Д. А. Внутрипопуляционные онтогенетические стратегии у зеленых лягушек (Pelophylax esculentus complex) // Вісник Харківського нац. ун-ту ім. В. Н. Каразіна. Серія: біологія. – 2015. – Вип. 25. – С. 223-238.
Шабанов Д. А. Еволюційна екологія популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) Лівобережного лісостепу України: автореф. дис. ... д-ра біол. наук за спец. 03.00.16 – екологія. – Дніпропетровськ, 2015. – 36 с.
Шабанов Д. А., Коршунов А. В., Кравченко М. А. и др. Внутрипопуляционные онтогенетические стратегии скороспелости и тугорослости: определение на примере бесхвостых амфибий // Вестник Харьковского национального университета имени В. Н. Каразина, серия "Биология". – 2014. – Вып.22, №1126. – С. 115-124.
Шабанов Д. А., Коршунов О. В., Кравченко М. О. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми // Біологія та валеологія. – Вип. 11. – Харків: ХДПУ, 2009. – С. 116–125.
Biriuk O., Shabanov D., Korshunov O. et al. Gamete production patterns and mating systems in water frogs (hybridogenetic Pelophylax esculentus complex) in North-Western Ukraine // J. of Zoology Systematics and Evolutionary Res. – 2016. – 54 (3). – P. 215–225. DOI: 10.1111/jzs.12132.
Borkin L. J., Korshunov A. V., Lada G. A. et al. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian J. of Herpetology. – 2004. – 11 (3). – P. 194–213.
Borkin L. J., Lada G. A., Litvinchuk S. N. et al. The first record of mass triploidy in hybridogenic green frog Rana esculenta in Russia (Rostov oblast’) // Russian Journal of Herpetology. – 2006. –13 (1). – P. 77–82.
Dedukh D., Litvinchuk S., Rosanov J. et al.Optional endoreplication and selective elimination of parental genomes during oogenesis in diploid and triploid hybrid european water frogs // PLoS ONE. – 2015. – 10(4): e0123304. doi:10.1371/journal.pone.0123304.
Mezhzherin S. V., Morozov-Leonov S. Yu., Rostovskaya O. V. et al. The ploidy and genetic structure of hybrid population of water frogs Pelophylax esculentus complex (Amphibia, Ranidae) of Ukraine fauna // Cytology and Genetics. – 2010. – Vol. 44, No 4. – P. 212–216.
Plötner J. Die westpaläarktichen Wasserfrösche. – Bielefeld: Laurenti-Verlag, 2005. – 161 S.
Shabanov D., Usova O., Kravchenko M. et al. Sustainable coexistence of the parental species and hemiclonal interspecific hybrids is provided by the variety of ontogenetic strategies // Herpetological Facts Journal – 2015. – 2. – P. 35–43.
Vinogradov A. E., Borkin L. J., Günther R., Rosanov J. M. Genome elimination in diploid and triploid Rana esculenta males: cytological evidence from DNA flow cytometry // Genome. – 1990. – Vol. 33, № 5. – P. 619–627.
Шабанов Д. А., Бірюк О. В., Коршунов О. В., Кравченко М. О. Поширення різних типів геміклональних популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) у басейні Сіверського Донця // Сучасний стан та охорона природних комплексів в басейні Сіверського Дінця. Мат. наук.-практ. конф. з нагоди 20-річчя НПП “Святі гори”. – Святогірськ, ТОВ “Видавництво “Друкарській двір”, 2017. – С. 139-144.