Особливості геміклональної популяційної системи зелених жаб (Pelophylax esculentus complex) Корякового яру (НПП «Гомільшанські ліси»)
Стах В. О.1, Боброва А. А.2, Єрмаков Д. В.2, Мелешко О. В.2, Тарасенко К. С.2, Коршунов О. В.2, Кравченко М.О.2
1Львівський національний університет імені Івана Франка
2Харківський національний університет імені В.Н. Каразіна
marinakravchenko2106@gmail.com
Місцевість під назвою «Коряків яр», що розташована поблизу с. Гайдари Зміївського району Харківської області, примітна тим, що саме тут у 1914 р. була створена Сіверсько-Донецька біологічна станція Товариства дослідників природи при Харківському університеті. Це місце було обрано тому, що попередні дослідження членів Товариства довели, що нагірна діброва в околицях колишнього хутору Зайцево є «надзвичайно цікавим в природничо-історичному сенсі місцем» [1]. За роки існування біологічної станції університету (тепер — Харківського національного університету імені В.Н. Каразіна) її перемістили приблизно на 1,5 км у північно-східному напрямку, у колишній маєток фотографа О.М. Іваницького.
Наша робота присвячена зеленим жабам, які населяють невеликий ставок, розташований у Коряковому ярі (49º36´57´´ пн. ш.; 36º18´44´´ сх. д.). Цей ставок лежить у нагірній діброві над заплавою Сіверського Донця, на території Національного природного парку «Гомільшанські ліси». Як відомо, зелені жаби (Pelophylax esculentus complex) – гібридогенний комплекс, що складається із двох батьківських видів: ставкової жаби Pelophylax lessonae (Camerano, 1882) та озерної жаби Pelophylax ridibundus (Pallas, 1771), а також різних за плоїдністю та складом геномів форм їхніх міжвидових гібридів. Для останніх застосовується назва, аналогічна до видової: їстівна жаба Pelophylax esculentus (Linnaeus, 1758). Особливістю P . esculentus є їхня здатність до геміклонального спадкування – передачі у гаметах геному одного з батьківських видів без рекомбінації, клонально. Відтворення P . esculentus у більшості випадків відбувається внаслідок спільного розмноження з представниками батьківських видів. Такі сукупності різних форм зелених жаб, що спільно відтворюються і передають як рекомбінантні, так і клональні геноми, названі геміклональними популяційними системами, ГПС або HPS [9].
Наявність у ГПС P. lessonae, P. ridibundus та P. esculentus позначається першими літерами їхніх видових назв [2, 8, 14 зі змінами]. Так, для Східної України є характерними R-E-HPS, до складу яких входять P. ridibundus і P. esculentus. Наявність поліплоїдних P. esculentus позначається Ep; у разі якщо P. esculentus представлені лише самицями або лише самцями, це позначається буквами f або m. Наприклад, у Сіверському Донці в околицях НПП «Гомільшанські ліси» трапляється R-E-Ep-HPS, до складу якої входять P. ridibundus, ди- та триплоїди P. esculentus обох статей. На відміну від заплави Сіверського Донця, ставок у с. Замулівка (колишнє Жовтневе) Вовчанського району Харківської області населяє R-E-Epf-HPS, в якій триплоїдні гібриди представлені лише самицями [8].
Вивчення ГПС Корякового яру почав у 1995–1996 рр. Г. А. Лада – тамбовський батрахолог, який співпрацював із Зоологічним Інститутом РАН (м. Санкт-Петербург). Він виявив у Коряковому яру, а також у Іськівому ставку (якому присвячена інша публікація у цьому збірнику) популяційні системи, що складалися лише з диплоїдних гібридів [8].
На початку ХХІ ст. в Коряковому яру траплялися нечисленні зелені жаби. Цей ставок є об’єктом гідробіологічних досліджень під час практики студентів ХНУ імені В.Н. Каразіна; в якому знаходили поодиноких пуголовків зелених жаб. У 2002 р. співробітники університету спільно з представниками Зоологічного інституту РАН розпочали систематичні дослідження складу зелених жаб у Харківській області. Ці дослідження привели до виявлення регіону масового поширення триплоїдних гібридів [4, 12], який пізніше було названо Сіверсько-Донецьким центром різноманіття Pelophylax esculentus complex [9]. За період 2002-2013 рр. за допомогою проточної ДНК-цитометрії було встановлено генотипи 45 жаб з Корякового яру.
Автори висловлюють щиру подяку С.М. Литвинчуку та Ю.М. Розанову за виконання цитометричного аналізу та надання його результатів. Згідно з цими результатами, у Коряковому яру знайдено 20 незрілих (включно з метаморфами) P. ridibundus, 8 незрілих і 11♂♂ P. esculentus (2n), 1 незріла і 1♀ P. esculentus (3n) з геномною композицією LLR, а також 1 незріла особина і 2♂♂ P. esculentus (3n) з геномною композицією LRR (літерою L позначено геном P . lessonae, а R – P . ridibundus). Ймовірно, відсутність диплоїдних самиць P. esculentus є випадковою і пов’язана з тим, що за цей час у ставку було знайдено лише одну самку.
У 2015 р. ситуація докорінно змінилася. У травні 2015 р. у ставку спостерігався нерестовий котел, в якому, за підрахунками О.В. Коршунова, було не менше як 67 самців, усі без винятку, судячи з особливостей вокалізації, – P. esculentus [10]. Як не дивно, після активного нересту в цьому оселищі не спостерігалося значної кількості пуголовків. Завдяки багаторазовим пошукам, вдалося виловити лише двох пуголовків; їхню плоїдність було встановлено із застосуванням каріоаналізу; обидва виявилися диплоїдами [7].
У 2015 р. було також розпочато оцінювання чисельності та складу ГПС Корякового яру із застосуванням мічення та повторного вилову. Вибірка, зібрана у червні та липні 2015 р., складалася (за даними Ю. Артемової, Т. Бешенцевої та О. Мелешко) з 5♂♂ P . ridibundus, 1♀ і 72♂♂ P. esculentus (2n), 4♀♀ і 1♂ P. esculentus (3n). Зібрана у липні 2016 р. вибірка складалася з 4♀♀ і 3♂♂ P. ridibundus, 1♀ і 27♂♂ P. esculentus (2n), 1♀ і 1♂ P. esculentus (3n). Різниця між складом вибірок 2015 і 2016 рр. статистично незначуща (p=0,53). Таким чином, за даними багаторічних спостережень, у Коряковому яру існує R-E-Ep-HPS зі значним переважанням самців диплоїдних гібридів. Розрахована за методом Петерсона [6] чисельність ГПС у 2015 р. становила 413±177 особин. На жаль, у вибірці 2016 р. мітки 2015 року виявлені не були.
За даними проточної цитометрії частка P. ridibundus вища серед незрілих особин порівняно зі статевозрілими. Ми розглядаємо це як підтвердження гіпотези [2], згідно з якою доля диплоїдів серед статевонезрілих жаб вища внаслідок того, що до їхнього числа належать представники батьківських видів, які утворилися внаслідок схрещування гібридів (т. зв. гібридолізу). Відомо, що такі схрещування призводять до появи нежиттєздатного потомства [13].
Кількість пуголовків у ставку в 2016 р. підвищилася у багато разів: за одну екскурсію у липні 2016 було зібрано 15 особин. Щоб встановити ймовірні причини відсутності пуголовків у 2015 р., наступного – 2016 р., ми перевірили фертильність самців. Кожна особина отримувала ін’єкцію з 1 мкг сурфагону – синтетичного гонадотропного рилізинг-гормону, на 1 г маси [3]. Через 2 год у самця шляхом масажу отримували уринальну сперму та підраховували в ній кількість активних сперматозоїдів на 1 мкл. З трьох досліджених самців P. ridibundus у одного сперматозоїди в уринальній спермі були відсутні, у іншого їхня кількість була зниженою (близько 5000 на мкл.), а у останнього була нормальною (більше 15000 на мкл.). Серед 28 самців P. esculentus сперматозоїди були відсутні у 23, у трьох особин (у тому числі — в єдиного триплоїда у вибірці) кількість сперматозоїдів була зниженою (близько 3000 у триплоїда, 3000 і 7000 у двох диплоїдів), і лише у двох самців була зареєстрована нормальна кількість сперматозоїдів (більше 15 000 на мкл). Слід зазначити, що кілька самців із вибірки 2016 р. мали невиражені вторинні статеві ознаки. У них були недорозвинені шлюбні мозолі та резонатори. Одного з таких самців для перевірки визначення статі було піддано анатомічному розтину. Він мав недорозвинені сім’яники. Усі такі фемінізовані самці не виробляли активних сперматозоїдів під впливом гормональної стимуляції.
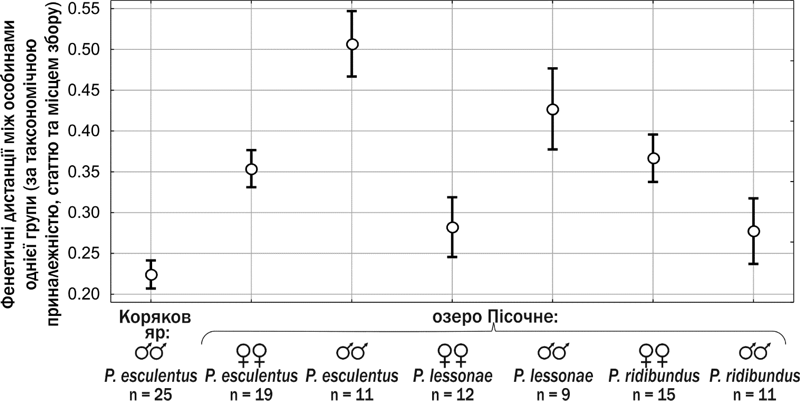
Порівняння внутрішньогрупової мінливості зелених жаб із двох місцеперебувань. Вертикальні лінії позначають 0,95% довірчий інтервал значення середнього для фенетичних дистанцій між різними особинами в кожній групі
Ці результати дають підстави припустити, що спалах чисельності зелених жаб, який спостерігався у 2015 р., є наслідком одного «вдалого» виводка або кількох виводків, у яких передавався один клональний геном (вірогідніше — чоловічий геном P . lessonae, отриманий від самця), що призвів до порушень фертильності самців P. esculentus. Щоб перевірити це припущення, ми провели аналіз фенетичного різноманіття самців P. esculentus та представників усіх трьох форм Pelophylax esculentus complex із озера Пісочне (51º56´94´´ пн. ш.; 23º90´23´´ сх. д.). Кожна особина була описана за 8 ознаками: характером дорзомедіальної смуги, поверхні шкіри, плям на спині, забарвлення черева, горла, морди та ніг, а також відносним розташуванням задньогомілкових з’єднань (див. рисунок). Для кожної пари особин у кожній групі (за таксономічною приналежністю, статтю і походженням) розраховували фенетичну дистанцію: частку відмінностей між особинами. Так, дистанція 1 відповідає особинам, які мають різні стани усіх 8 досліджених ознак, а дистанція 0,125 — особинам, що відрізняються лише за однією ознакою.
Судячи з рисунка, можна впевнитися, що самці P. esculentus з Корякового яру є значно однорідніші, ніж представники інших груп (відмінності мають високу статистичну значущість). Це свідчить про їхню високу генетичну подібність, що може бути наслідком поширення в ярі одного клонального геному. Певним парадоксом є те, що для того, аби поширитися у ставку, цей клональний геном мав забезпечити успішне відтворення його носія, а на даному етапі ми спостерігаємо, що він призводить до порушень статевого розвитку. У будь-якому разі, подальші дослідження ГПС Pelophylax esculentus complex з Корякового яру становлять значний пізнавальний інтерес.
1. Арнольди В.М. Северо-Донецкая Биологическая станция Об-ва испытателей природы при Харьковском университете // Харьков, 1918. 16 с.
2. Бирюк О.В., Усова Е.Е., Мелешко Е.В., Шабанов Д.А. Устойчивость сперматогенеза и проявления отбора на клеточном и индивидуальном уровне у незрелых представителей Pelophylax esculentus complex // Вісник Дніпропетр. ун-ту. Біологія, екологія. 2016. 24(1). С. 193–202.
3. Боброва А.А., Макарян Р.М., Шейко В.П., Шабанов Д.А. Порушення фертильності у міжвидових гібридів зелених жаб із Сіверсько-Донецького центру різноманіття Pelophylax esculentus complex // Біологія та валеологія. 2014. Вип. 16. С. 7–15.
4. Боркин Л.Я., Зиненко А.И., Коршунов А.В. и др. Массовая полиплоидия в гибридогенном комплексе Rana esculenta (Ranidae, Anura, Amphibia) на Востоке Украины // Мат. І конф. Укр. герпетол. т-ва. К.: Зоомузей ННПМ НАНУ, 2005. С. 23–26.
5. Лада Г.А. О необходимости сохранения уникальных «чистых» популяций диплоидной съедобной лягушки (Rana esculenta Linnaeus, 1758) в Белгородской и Харьковской областях // Проблемы охраны и рационального использования природных экосистем и биологических ресурсов. Пенза, 1998. С. 333–335.
6. Коли Г. Анализ популяций позвоночных. М.: Мир, 1979. 362 с.
7. Пустовалова Э.С., Батуева Е.Д., Харченко Т.С. Плоидность головастиков зеленых лягушек из Иськова пруда и Корякова яра (Змиевской район Харьковской области) // Біологія: від молекули до біосфери: матеріали X Міжнар. конф. молодих учених (2–4 грудня 2015 р., м. Харків, Україна). Х.: ФОП Шаповалова Т. М., 2015. С. 183-184.
8. Шабанов Д.А. Еволюційна екологія популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) Лівобережного лісостепу України: автореф. дис. ... д-ра біол. наук за спец. 03.00.16 — екологія. Дніпропетровськ, 2015. 36 с.
9. Шабанов Д.А., Коршунов О.В., Кравченко М.О. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми // Біологія та валеологія. 2009. Вип. 11. С. 116–125.
10. Шабанов Д.А. Пруды в «Больших Гайдарах»: Иськов пруд, Коряков яр, Нижний Добрицкий пруд // http://batrachos.com/Иськов-Коряков-Добрицкий [Электронный ресурс]
11. Biriuk O., Shabanov D., Korshunov O. et al. Gamete production patterns and mating systems in water frogs (hybridogenetic Pelophylax esculentus complex) in North-Western Ukraine // Journal of Zoological Systematics and Evolutionary Research. 2015. Vol. 54. I. 3. P. 215–225.
12. Borkin L.J., Korshunov A.V., Lada G.A et al. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian J. of Herpetology. 2004. Vol.11 (3). P. 194–213.
13. Günther R., Plötner J. Zur Problematik der klonalen Vererbung bei Rana kl. esculenta (Anura) // Beiträge zur Biologie und Bibliographie (1960-1987) der europäischen Wasserfrösche / R. Günther, R. Klewen. – Duisburg: Verlag für Ökologie und Faunistik, 1988. P. 23–46.
14. Uzzell T.M., Berger L. Electrophoretic phenotypes of Rana ridibunda, Rana lessonae and their hybridogenic associate Rana esculenta // Proc. Acad. nat. Sci. Phila. 1975. Vol. 127. P. 13–24.