Вивчення флуктуючої асиметрії річкового окуня (Perca fluviatilis L., 1758)
Виноградова К. П. 1, Сакун Ю. В. 1, Белоусова К. М. 1, Гончаров Г. Л. 2, Шабанов Д. А. 1
1 Харківський національний університет ім. В.Н. Каразіна;
2 Національний природний парк «Гомільшанскі ліси»
256 річкових окунів (Perca fluviatilis L., 1758) було відловлені на мілинах річки Сіверський Донець в Зміївському районі Харківської області. Ці окуні належали до трьох вікових груп (0+, 1+ і 2+). З обох боків кожної риби визначено кількість лусок вздовж бічної лінії, а також кількість променів в грудних і черевних плавцях. Показано, що ці ознаки можуть бути мірою флуктуючої асиметрії. Встановлено, що рівень флуктуючої асиметрії з віком знижується. Найнижча асиметрія характерна для особин з середнім для вікової групи розміром, тобто для особин з середньою швидкістю росту. Ці результати розглядаються як прояви дії стабілізуючого добору.
Ключові слова: Perca fluviatilis, флуктуюча асиметрія, швидкість росту, стабілізуючий добір.
Vinogradovа K. P, Sakun Yu. V., Belousova K. M., Goncharov G.L., Shabanov D. A. The study of fluctuating asymmetry of European perch (Perca fluviatilis L., 1758). 256 river perch (Perca fluviatilis L., 1758) was captured at the shallows on the Seversky Donets River in the Zmijivskij rajon (Kharkiv oblast). These perches belonged to the three age groups (0 +, 1 + and 2 +). Number of scales was count on both sides of each fish along the lateral line, number of rays in the pectoral and pelvic fins also was count. It is shown that these features are largely affected by fluctuating asymmetry. Found that the level of fluctuating asymmetry decreases with age. Lowest asymmetry is typical for individuals with an average size for the age group, ie individuals with an average growth rate. These results are considered as a manifestation of stabilizing selection.
Keywords: Perca fluviatilis, fluctuating asymmetry, growth rate, stabilizing selection.
Виноградова К. П., Сакун Ю. В., Белоусова К. М., Гончаров Г. Л., Шабанов Д. А. Изучение флуктуирующей асимметрии речного окуня (Perca fluviatilis L., 1758). 256 речных окуней (Perca fluviatilis L., 1758) были отловлены на отмелях реки Северский Донец в Змиевском районе Харьковской области. Эти окуни принадлежали к трем возрастным группам (0 +, 1 + и 2 +). С обеих сторон каждой рыбы определено количество чешуй вдоль боковой линии, а также количество лучей в грудных и брюшных плавниках. Показано, что эти признаки могут быть мерой флуктуирующей асимметрии. Установлено, что уровень флуктуирующей асимметрии с возрастом снижается. Низкая асимметрия характерна для особей со средним для возрастной группы размером, то есть для особей со средней скоростью роста. Эти результаты рассматриваются как проявления действия стабилизирующего отбора.
Ключевые слова: Perca fluviatilis, флуктуирующая асимметрия, скорость роста, стабилизирующий отбор.
Вступ. На склад та особливості вибірок з природних популяцій, які збирають під час польових біологічних досліджень, впливає цілий комплекс процесів, дія яких прихована від дослідників. Так, морфологічні особливості тварин є наслідком онтогенезу – самоорганізації, на яку впливає як внутрішня програма, так і зовнішні впливи. На склад природних популяцій впливає природний добір, що може залежати від різних чинників та бути спрямованим у різних напрямах.
Різні онтогенетичні процеси є різною мірою зарегульованими. Стійкість цих процесів досить важко спостерігати прямо, але можна оцінювати побічно. У кожному випадку ми можемо спостерігати певний результат онтогенезу, але не можемо визначити, з якою вірогідністю могли реалізуватися якісь інші результати. У більшості випадків ми не можемо спостерігати за розвитком сукупності клонів організмів у однакових умовах середовища. Але у багатьох випадках визначити стійкість онтогенезу можна, вивчаючи не міжіндивідуальну, а внутрішньоіндивідуальну мінливість.
Флуктуюча асиметрія – випадкові, неспрямовані відхилення від білатеральної симетрії організмів, які можна розглядати як наслідки «онтогенетичного шуму» – неповної здатності організмів розвиватися строго визначеними шляхами [3; 5]. Оскільки флуктуюча асиметрія має стохастичну природу, вивчати її можна не на рівні окремих особин, а лише розглядаючи їх групи під час дослідження вибірок.
У багатьох роботах показано, що рівень флуктуючої асиметрії зростає під впливом несприятливих умов розвитку, у гібридних організмів з незбалансованим генотипом тощо [2]. Згідно з класифікацією видів асиметрії, що дав Л. Ван Вален [9], слід розрізняти направлену та флуктуючу асиметрію, а також антисиметрію. Стійкість розвитку можна оцінювати лише за допомогою флуктуючої асиметрії.
Особливий інтерес представляє вивчення флуктуючої асиметрії у природних вибірках, що складаються з особин різного віку.
Матеріал і методи. Дане дослідження виконане як учбово-дослідна робота студентів під час навчально-польової практики студентів другого курсу біологічного факультету Харківського національного університету імені В. Н. Каразіна. У роботі вивчено вибірку з 256 особин річкового окуня (Perca fluviatilis Linnaeus, 1758), які були виловлені 12, 13 і 23 червня 2012 року мальковою волокушею з кроком вічка 6 мм на трьох піщаних мілинах річки Сіверський Донець у околицях біологічної станції Харківського національного університету імені В. Н. Каразіна (село Гайдари, Зміївський район Харківської області) у рамках лімітів на вилов, отриманих Національним природним парком «Гомільшанські ліси».
У кожної особини за допомогою штангенциркулю визначали стандартну довжину тіла L (в мм.), а також під бінокулярним мікроскопом підраховували кількість лусок (S) вздовж бічної лінії с правого (Sr) та лівого (Sl) боків тіла, кількість променів число в грудних (P) і черевних (V) плавцях з правого та лівого боків (Pr і Pl; Vr і Vl відповідно).
Досліджених риб розділили на розмірні групи за довжиною тіла. Зразки луски представників кожної вікової групи досліджували під бінокулярним мікроскопом за стандартними методиками [4], підраховуючи кількість зон уповільненого росту (кілець зближених склеритів), що відповідають зимівлям.
Статистичну обробку результатів вимірювання асиметрії досліджуваних ознак проводили згідно з програмою, викладеною у роботі Д. В. Гелашвілі з співавторами [1] з використанням програми Statistica 7.0 (StatSoft). Для кожної з особин обчислювали значення асиметрії за досліджуваними ознаками: DS=Sr-Sl, DP=Pr-Pl, DV=Vr-Vl та абсолютні значення асиметрії за цими ж ознаками: AS=│Sr-Sl│, AP=│Pr-Pl│, AV=│Vr-Vl│. Крім того, для кожної особини обчислювали загальний показник асиметрії: Σ(A)= AS+AP+AV.
Результати та обговорення.
1. Відповідність між розмірними та віковими групами. Кількісний, розмірний та віковий склад досліджуваної вибірки особин показано у табл. 1 та на рис. 1. Як показав аналіз луски, розмірні групи відповідають віковим: особини розмірної групи І не мають зон зближених склеритів, що відповідають зимівлям, розмірної групи ІІ мають одну таку зону, а розмірної групи ІІІ – дві зони. Як можна побачити (табл. 1; рис. 1) розмірні групи розділені розривом та відокремлені одна від одної досить добре.
Таблиця 1. Розмірні (вікові) групи досліджуваних особин Perca fluviatilis
|
Розмірні групи |
Вік |
Довжина тіла, мм: середня (мінімальна – максимальна) |
Кількість особин |
|
I |
0+ |
39,5 (30 – 57) |
196 |
|
II |
1+ |
89,6 (74 – 116) |
51 |
|
ІІІ |
2+ |
137,2 (123 – 160) |
10 |
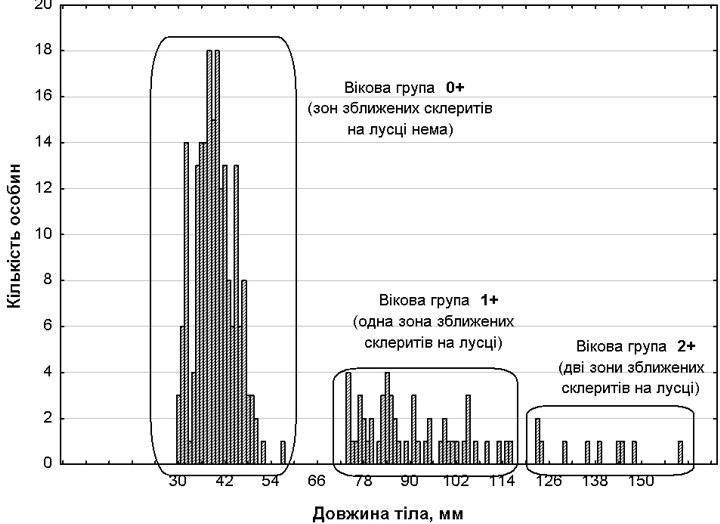
Рис. 1. Розмірний та віковий розподіл досліджуваних особин Perca fluviatilis
2. Аналіз розподілів мір асиметрії. Для того, щоб застосувати асиметричність за певними ознаками як міру нестійкості розвитку, слід впевнитися, що мінливість цих ознак відбиває саме флуктуючу асиметрію [1]. Значення флуктуючої асиметрії мають розподіл, близький до нормального, який розташований симетрично відносно нульового значення. Саме такий розподіл свідчить, що на мінливість ознаки впливає «онтогенетичний шум». Розподіли досліджуваних ознак показані на рис. 2. Легко побачити, що розподіл усіх ознак є симетричним, тобто асиметрія за усіма дослідженими ознаками є неспрямованою.
Д. В. Гелашвілі з співавторами [1] рекомендують проводити статистичний тест на відповідність розподілу ознаки, що вивчається, нормальному. З цим не можна погодитися повністю. Річ у тім, що для однієї і теж ознаки, чим краще охарактеризований її розподіл (чим більшою є досліджувана вибірка), тим вірогідніше, що відхилення цього розподілу від нормального, обчислюване за загальноприйнятними статистичними критеріями, буде значущим. Для використання ознаки, що відбиває флуктуючу асиметрію, важливою є певна відповідність її розподілу до нормального (симетричність відносно нульового значення; найчисельнішість симетричних особин), а не незначущість відхилень цього розподілу від нормального, що оцінена за статистичними критеріями.
Інтерес викликає форма розподілу асиметрії за кількістю лусок вздовж бічної лінії (рис. 2, А). Кількість особин, у яких різниця між лівим та правим боками становить 2 луски, більша за кількість тих, у яких ця різниця становить 1 луску, що надає розподілу характерну тривершинну форму. Цей ефект не є випадковим: порівняння часток за Р. Фішером з використанням однобічного критерію (72 особини з 256, у яких SA=1, та 91 особина з 256, у яких SA=2) доводить, що цей ефект є значущим (p=0,04).
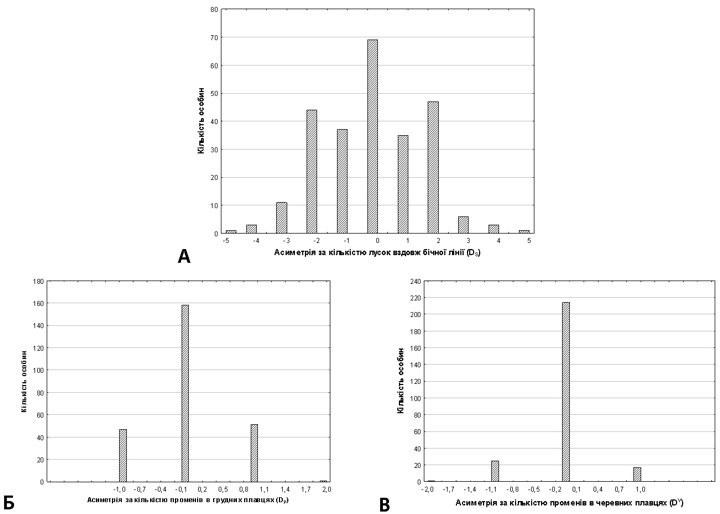
Рис. 2. Розподіл досліджуваних асиметричних ознак. А: DS; Б: DP; В: DV
Вірогідно, пояснити особливості розподілу асиметрії за кількістю лусок можна, згадавши, що нормальний розвиток є каналізованим внаслідок дії стабілізуючого добору. Застосовуючи метафору епігенетичного ландшафту (рис. 3), що запропонована К. Х. Уоддингтоном [10], можна припустити, що шлях розвитку, який веде до утворення симетричної особини, є заглибленим (стабілізованим). Невеликі відхилення від симетричності розвитку, згідно з цим припущенням, з певною вірогідністю коректуються.
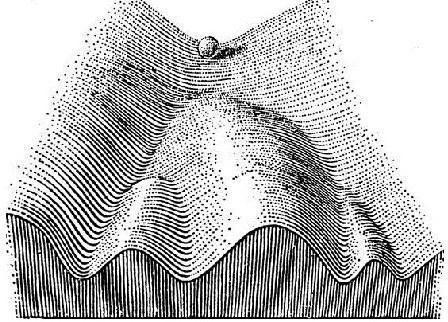
Рис. 3. Епігенетичний ландшафт за К. Х. Уоддингтоном [10]. Онтогенез порівнюється зі скатуванням кулі поверхнею складної форми. Вірогіднішому стану системи, що розвивається, відповідають заглиблені ділянки поверхні
Якщо прийняти це припущення, можна дійти до висновку, що усі досліджені ознаки можна використовувати як показники флуктуючої асиметрії, оскільки вони відбивають саме онтогенетичний шум – стохастичні впливи на процес онтогенезу.
3. Перевірка кореляції між мірами та їх розмір-залежності. Наступними кроками перевірки того, чи пригідні вимірювані ознаки для характеристики флуктуючої асиметрії, є обчислення зв’язку абсолютної асиметрії з розміром та розрахунок коефіцієнтів кореляції між різними показниками асиметрії. Ці результати наведені у табл. 2.
Таблиця 2. Значення коефіцієнту кореляції Спірмена (rS) та їхня значущість (p) для розмірних показників та абсолютних показників асиметрії. Комірки, що дублюються, заповнені в одному місці; значущі зв’язки виділено напівжирним
|
|
L (довжина тіла) |
AS |
AP |
AV |
|
Sr+Sl (сума кількості лусок на обох боках) |
rS=0,23 p=0,0003 |
rS=–0,08 p=0,16 |
|
|
|
Pr+Pl (сума кількості променів в грудних плавцях) |
rS=0,39 p=0,0000 |
|
rS=–0,06 p=0,27 |
|
|
Vr+Vl (сума кількості променів в черевних плавцях) |
rS=0,06 p=0,33 |
|
|
rS=–0,52 p=0,0000 |
|
AS (абсолютна асиметрія за кількістю лусок) |
rS=–0,36 p=0,0000 |
— |
rS=–0,07 p=0,21 |
rS=0,14 p=0,027 |
|
AP (абсолютна асиметрія за променів у грудних плавцях) |
rS=0,04 p=0,52 |
|
— |
rS=0,20 p=0,001 |
|
AV (абсолютна асиметрія за променів у черевних плавцях) |
rS=–0,09 p=0,14 |
|
|
— |
На підставі даних, наведених у табл. 2, можна зробити наступні висновки. Усі використані міри асиметрії демонструють від’ємну кореляцію з кількістю структур, що підраховувалися; з цих зв’язків значущим є тільки той, що стосується кількості променів в черевних плавцях. З ростом лінійних розмірів реєструється значуще зниження асиметрії за кількістю лусок; зв'язок з двома іншими параметрами є несуттєвим. З ростом лінійних розмірів збільшується кількість структур, що рахувалися (для кількості променів в черевних плавцях цей зв'язок є незначущим). Асиметрія за кількістю променів в черевних плавцях значуще пов’язана з асиметрією за кількістю лусок та кількістю променів в грудних плавцях; зв'язок двох останніх величин є незначущим.
Розмір-залежність мір асиметрії (випадок, якщо асиметрія пов’язана з кількістю досліджуваних структур) може призвести до того, що, ті особини, у котрих таких структур буде більше, будуть помилково розглядатися як особини з менш стійким онтогенезом. У тих випадках, коли досліджувані величини демонструють розмір-залежність, Д. В. Гелашвілі з співавторами [1] рекомендують проводити нормування (відносити міру асиметрії до кількості структур, що підраховуються).
Складніше інтерпретувати від’ємну кореляцію асиметрії з кількістю меристичних структур. Зменшення рівню асиметрії зі зростанням кількості перелічуваних структур не є суто арифметичним феноменом; пояснити цей ефект можна тільки дією певного фактора, що зменшує асиметрію у разі збільшення меристичніх структур. Крім того, ознака, для якої зареєстрована від’ємна розмір-залежність, значуще пов’язана з двома іншими мірами асиметрії. З урахуванням цих обставин, ми для подальшого аналізу ми використовували абсолютні міри асиметрії та їх суму: Σ(A)= AS+AP+AV.
4. Зв'язок асиметрії з віком та швидкістю росту особин. У нашій роботи досліджуються три вікові групи P. fluviatilis. Але ці групи не є однорідними (рис. 1): в кожній з них можна виділити особин, що мають відносно малі, середні, або великі розміри. Нерест P. fluviatilis одноразовий і відбувається в умовах певної водойми переважно протягом 9-13 діб, іноді до місяця [8]. У останні роки за відсутності значних повеней та при досить швидкому зростанні температури води у весняний період масовий нерест окуня на русловій ділянці Сіверського Дінця на території НПП «Гомільшанські ліси» відбувається переважно протягом 7 – 10 днів. Якщо вік цих особин є майже однаковим, ця різниця у розмірах пов’язана зі швидкістю росту. За цією ознакою ми розділили кожну вікову групу на три близькі за чисельністю підгрупи (табл. 3). Відхилення від рівної чисельності підгруп пов’язані з тим, що усіх особин, однакових за довжиною тіла, ми відносили до однієї підгрупи.
Таблиця 3. Підгрупи, що виділені за швидкістю росту, у кожній з розмірно-вікових груп P. fluviatilis
|
Розмірно-вікові групи та їх чисельність (n) |
Підгрупи за швидкістю росту: n, особин; (min-max), мм |
||
|
Повільноростучі |
Середні за швидкістю росту |
Швидкоростучі |
|
|
І (0+); n=195 |
n=68; (30–37) |
n=63; (38–41) |
n=64; (42–57) |
|
ІІ (1+); n=51 |
n=15; (74–82) |
n=19; (83–94) |
n=17; (95–116) |
|
ІІІ (2+); n=10 |
n=4; (123–130) |
n=2; (136–139) |
n=4; (144–160) |
Ми провели двохфакторний дисперсійний аналіз, в якому як фактори розглядали вік P. fluviatilis та швидкість їх росту, а у якості величини, мінливість якої залежить від цих факторів, розглядали загальний показник асиметрії Σ(A). Результати цього аналізу показані в табл. 4 та на рис. 4.
Як вплив віку, так і вплив швидкості росту виявилися значущими, але вплив віку є більш важливим. Як можна пояснити те, що рівень асиметричності у вибірках P. fluviatilis з віком зменшується?
Можна висунути дві гіпотези. Перша (гіпотеза виправлення) полягає в тому, що з часом певні особини можуть ставати біль симетричними. Згідно з другою (гіпотезою стабілізуючого добору), асиметричні особини мають вищі шанси загинути, і тому їхня частка у генерації з часом зменшується. Гіпотеза виправлення здається нам маловірогідною, хоча б тому, що процеси, які вона припускає, раніше, наскільки нам відомо, не були зареєстровані. На користь гіпотези стабілізуючого добору свідчить і те, що найменший рівень асиметрії є характерним для особин з середньою швидкістю росту, а найвищий – у найменших в своїх розмірних групах особин. Слід врахувати, що саме стабілізуючий добір є причиною стійкості нормального розвитку [7].
Таблиця 4. Результати дисперсійного аналізу
|
Ефект |
SS Сума квадратів |
df Ступенів свободи |
MS Середній квадрат |
F Крітерій Фішера |
p Значущість |
|
Вік |
31,01 |
2 |
15,51 |
10,4 |
0,00005 |
|
Швидкість росту |
10,05 |
2 |
5,03 |
3,38 |
0,036 |
|
Взаємодія віку та швидкості росту |
2,10 |
4 |
0,53 |
0,35 |
0,84 |
|
Похибка |
367,78 |
247 |
1,49 |
|
|

Рис. 4. Залежність загального показника асиметрії P. fluviatilis від віку та швидкості росту. Вертикальні лінії показують 95% довірчий інтервал значень показника асиметрії для кожної групи (за віком) та підгрупи (за швидкістю росту)
Комплекс викладених обставин дозволяє нам вважати, що зареєстроване зниження флуктуючої асиметрії у старших вікових групах є наслідком дії стабілізуючого добору, що усуває більш асиметричних особин.
Висновки. У роботі зареєстровані прояви флуктуючої асиметрії за трьома меристичними ознаками P. fluviatilis. Рівень асиметрії пов'язаний зі швидкостю росту і є найменшим для особин з середнім розміром для своєї вікової групи. У вибірках старшого віку рівень флуктуючої асиметрії нижчий, ніж у вибірках молодшого віку. Вірогідно, це є наслідком дії стабілізуючого добору, що усуває більш асиметричних особин.
Література.
- Гелашвили Д. Б., Якимов В. Н., Логинов В. В., Епланова Г. В. Cтатистический анализ флуктуирующей асимметрии билатеральных признаков разноцветной ящурки Eremias arguta// Актуальные проблемы герпетологии и токсинологии: Сборник научных трудов. Вып. 7. — Тольятти, 2004. — С. 45–59.
- Гавриков Д. Е. Асимметрия билатеральных признаков позвоночных животных // Бюллетень ВСНЦ СО РАМН, 2007. — №2. — С. 26–28.
- Захаров В. М. Асимметрия животных. М.: Наука, 1987. — 216 с.
- Правдин И.Ф. Руководство по изучению рыб. – М.: Пищевая промышленность, 1966. — 374 с.
- Романов Н. С. Флуктуирующая асимметрия лососей заводского и естественного воспроизводства // Чтения памяти В. Я. Леванидова. Владивосток: 2001. — С. 328–330.
- Халафян А. А. Statistica 6. Статистический анализ данных. — М.: ООО "Бином-ПРесс", 2007. — 512 с.
- Шишкин М. А. Индивидуальное развитие и эволюционная теория // Эволюция и биоценотические кризисы. М.: Наука, 1987. — С. 76–124.
- Щербуха А.Я. Фауна України. Том 8. Риби. Вип. 4. Окунеподібні (окуневидні, губаньовидні, драконовидні, собачковидні, піщанковидні, ліровидні, скумбрієвидні). — Київ: Наукова думка, 1982. — 384 с.
- Van Valen L. Study of fluctuating asymmetry / L. Van Valen // Evolution. — 1962. — Vol. 16, № 2. —P. 125–146.
- Waddington С. Н. The Strategy of the Genes: a Discussion on Some Aspects of Theoretical Biology. L.: Alien and Unwin, 1957. — 262 p.