Сетевые и бумажные СМИ недавно обошло сообщение про очередную палеонтологическую находку, которая подается чуть ли не как доказательство теории эволюции. Речь идет о верхнедевонской рыбе, получившей имя тиктаалик (Tiktaalik, рис. 1). Чтобы понять, в чем ценность этой находки и в чем ошибаются ее многочисленные комментаторы, надо разобраться в проблеме происхождения четвероногих.

Реконструкция тиктаалика1
От рыб к четвероногим
Мы, без ложной скромности, относимся к одной из самых «удачных» групп животных — позвоночным. Самые серьезные конкуренты позвоночных — членистоногие и головоногие. Благодаря «модульному» принципу сборки и жестким покровам членистоногие добились победы в малом размерном классе и в море (ракообразные), и на суше (насекомые). Но размер членистоногих жестко ограничен наружным скелетом и (для наземных представителей) трахейной дыхательной системой, которая должна подводить воздух к каждой клетке. Ограничения предельных размеров позвоночных менее жесткие, и лишь головоногие смогли приблизиться к ним по размерам тела. Увы (или к счастью), головоногие так и не сумели выйти из моря. В нашем мире часто выигрывает тот, кто крупнее, потому наша группа стала главным игроком в крупном размерном классе. План строения рыб позволил им оказаться доминирующей группой крупных животных в воде, а план строения четвероногих дал возможность завоевать сушу.
И рыбы, и четвероногие — удачные конструктивные решения. Но как перейти от одной конструкции к другой? Нужно ли пройти через стадию полурыбы-получетвероногого, или этот переход можно преодолеть в один скачок?
Эволюционное сальто
Классический дарвинизм (продукт второй половины XIX века) и синтетическая теория эволюции (середина XX века) предполагают, что эволюция идет маленькими шажками в течение длительного времени. Но сальтационная теория (от сальто — прыжок), возникшая в начале XX века, уверена, что эволюционные изменения могут проходить в одно поколение. Рассмотрим пример, подтверждающий этот взгляд.
Интересным этапом эволюции предков четвероногих было изменение устройства их ноздрей. Эволюционно орган обоняния развился из обонятельных ямок на передней части головы. Неудивительно, что ноздри первых рыб (как и большинства современных видов) не были связаны с дыхательной системой. Передняя ноздря открывается в обонятельный канал, из которого вода выходит в заднюю ноздрю (рис. 2). Движение рыбы вперед обеспечивает ток воды мимо обонятельного эпителия. У рыб, которые были нашими предками (их называют лопастепёрыми), передняя ноздря открывалась сверху на наружной поверхности головы, а «задняя» (называющаяся хоаной) — в ротовой полости, на верхнем нёбе. Ее наличие облегчает жизнь хищника-засадника, застывшего в стоячей воде. Через ротовую полость течет вода, которую прокачивают жабры. Благодаря этому обеспечивается ток воды через обонятельный канал, и рыба может почувствовать приближение жертвы.
Но как выход обонятельного канала переместился с верхней поверхности головы внутрь ротовой полости? Иван Иванович Шмальгаузен, советский эволюционист-классик, предположил, что отверстие носового канала смещалось к краю челюсти, где наблюдался хоть какой-то ток воды. И так ноздря в ряду поколений постепенно «переползла» внутрь рта. Увы, не ясно, как носовое отверстие «преодолело» усаженную зубами верхнечелюстную кость, стоявшую на ее пути. Вероятно, оно могло оказаться внутри ротовой полости лишь скачком. Рассмотрим аналогию.
У человека иногда встречается аномалия, называемая «волчья пасть». Дело в том, что во время нашей эволюции отверстие хоан оказалось отгорожено от ротовой полости вторичным костным небом. В норме у человека хоаны открываются не в ротовую полость, а в носоглотку (это позволяет дышать во время пережевывания пищи). «Волчья пасть» вызвана нарушением формирования вторичного костного неба. Заглянув в рот такому больному, можно увидеть зияющее отверстие, ведущее в носоглотку.
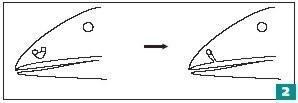
Хоаны (внутренние ноздри) — результат преобразования ноздрей, находившихся на поверхности головы
Видимо, в эволюции девонских рыб появилась аномалия, напоминающая «волчью пасть». Какой-то дефект развития привел к неполному образованию перегородки между обонятельным каналом и ротовой полостью. Это нарушение существенно улучшало работу органа обоняния и оказалось поддержано отбором. Развитие хоан стабилизировалось, а вторая наружная ноздря исчезла за ненадобностью. В результате этого события мы можем дышать с закрытым ртом!
Девонский мир
Однако переход от рыб к четвероногим в целом не мог пройти скачком, ведь для него необходимо согласованное изменение многих органов и систем. Видимо, в девоне между двумя адаптивными зонами (совокупностями образов жизни), соответствующими рыбам и четвероногим, существовал какой-то переход.
Обратимся к условиям жизни в девонском периоде. Это был во многом иной мир, в котором даже год состоял не из 365 с четвертью суток, а примерно из 4002. Растения лишь начали осваивать сушу, и континенты не были покрыты ни растительностью, ни почвой. Дожди размывали рыхлые породы, и водные потоки несли взвешенные частицы к морю. При замедлении тока воды эти частицы оседали, формируя донные отложения. И сегодня в местах впадения Дуная в Черное море или Волги — в Каспийское формируются обширные дельты. Донные осадки вдаются в море, а река разбивается на множество рукавов. Сегодняшние дельты зарастают растениями, укрепляющими границы суши и воды (если им не мешают приливы и отливы). В девоне осадков было намного больше, а растительности — куда меньше. На огромных мелководьях без четких границ суши и воды полуводную-полуназемную среду осваивали полурыбы-получетвероногие.
Важнейшие для наземной жизни особенности этих животных возникали как приспособления к обитанию в воде. Первое из таких приспособлений — легкие. Поскольку большинство наземных четвероногих дышат легкими, может показаться, что этот орган является приспособлением к наземной жизни. Это не так, что и доказывают современные рыбы. Те из них, которые имеют легкие3, не выходят на сушу, а те, кто выходят на сушу4, не используют легкие и дышат через кожу!
Судьба легких
На девонском мелководье дышать было нелегко. Разлагающиеся остатки растений поглощали растворенный в воде кислород. Однако воздух был рядом, и его нужно было только заглотить. Рыбы поднимались на поверхность и хватали пузырь воздуха ртом. Когда они опускались на дно, пузырь доходил до определенного места в пищеводе, где обилие кровеносных сосудов обеспечивало газообмен. Чтобы не мешать питанию, этот участок преобразовался в выпячивания — легкие. Увы, для дыхания на берегу легкие не годились. Тысячекратно возросший из-за отсутствия архимедовой силы вес рыбы выжимал воздух из ее легких. На суше приходилось дышать через кожу…
Легкие не только помогали рыбам дышать, но и удерживали их в толще воды, делая тело фактически невесомым. Когда такие рыбы заселили моря, дыхательная функция легких оказалась излишней, а гидростатическая пришлась весьма кстати. Легкое превратилось в плавательный пузырь. Некоторые рыбы переходили к жизни у дна и теряли этот орган. Некоторые из потерявших плавательный пузырь рыб осваивали пресные воды, где недостаток кислорода заставил их начать заглатывать пузыри воздуха с поверхности. Такое поведение можно увидеть у многих аквариумных сомиков и вьюнов. Круг замкнулся… Но почему он не пошел на второй оборот, почему у этих рыб не возникли новые легкие?
Ответ может дать эпигенетическая теория эволюции. С ее точки зрения, роль отбора состоит в стабилизации удачных вариантов индивидуального развития. В девоне рыбы были молодой группой, и их развитие было слабо зарегулировано. Те изменения плана строения, которые были возможны тогда, стали невозможными сейчас, после десятков и сотен миллионов лет отбора на стабилизацию «рыбьей» конструкции. А поведение, как-никак, перестраивается намного легче. В результате современные рыбы ведут себя как и их далекие предки, но не претерпевают аналогичных изменений строения5.
«Рыбы с ножками»
Второе важное для наземной жизни приобретение, возникшее в воде, — конечности. Как ни странно, они появились не для ползания по суше.
Осваивавшие переползание по мелководью и суше рыбы двигались способом, который называется «концертино». Тело рыбы изгибается в сторону. Грудной плавник на выгибающейся стороне тела проволакивается по опоре, а на сгибающейся — упирается в грунт и толкает тело вперед. Тело изгибается в противоположную сторону, и плавники меняются ролями. Такое перемещение не требует существенных изменений рыбьей конструкции. При нем работает та же мускулатура, что и при плавании, а плавник попросту преобразуется в жесткую опорную ось. Однако эта ось — еще не нога. А как же тогда возникли ноги?
Вам приходилось когда-либо плавать на мелководье? Представьте себе: вы гребете руками в воде и вдруг хотя бы кончиками пальцев опираетесь на дно и отталкиваетесь от него — при этом тело получает заметный толчок. Вот и рыбы-засадники, кидавшиеся на добычу на мелководье, тоже отталкивались от дна.
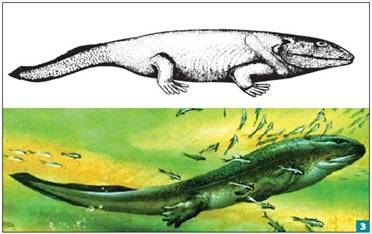
Классическая6 и современная реконструкция ихтиостеги — самого известного вида ранних четвероногих
А как вы будете отталкиваться от мягкого, илистого дна? Растопыренными пальцами. Они погрузятся в ил, но какой-то из них может нащупать что-то твердое и дать подходящую опору. Рыбий плавник из многих относительно мягких лучей не годился для этой роли, как не годилась и одна жесткая ось. Оптимумом была конечность с несколькими выростами — пальцами. Разные группы использовали варианты с числом пальцев от восьми до четырех, но самым ходовым оказался вариант пятипалой конечности.
Так возникли разнообразные животные, которых можно называть «рыбами с ногами». Полвека назад из них была известна лишь ихтиостега (Ichthyostega), найденная в верхнедевонских отложениях Гренландии (рис. 3). Сегодня описано не меньше десятка родов девонских четвероногих, «рыб с ножками». Древнейшая из них — акантостега (Acanthostega, рис. 4). Последние исследования изменили прежние представления об образе жизни этих животных. Вероятно, они были еще первичноводными и практически не выходили на сушу. Настоящее освоение суши началось позже, в каменноугольном периоде…
Откуда должны расти руки
Сравнивая первых четвероногих друг с другом, можно использовать много разных признаков. Рассмотрим один из них — связь плечевого пояса с черепом.

Рисунок и подпись к нему из школьного учебника7: Еще рыба (пандерихт, 378 млн. лет тому назад, наверху) и уже четвероногое (акантостега, 360 млн. лет тому назад). Нашими предками могли быть похожие на них животные
Наши руки — преобразованные грудные плавники рыб, которые возникли как рули и обеспечивали повороты при плавании. Грудные плавники опирались на расположенную в толще тела пластину — плечевой пояс; опорой брюшных плавников (наших ног) стал тазовый пояс. Чтобы усилие, развиваемое грудными плавниками, не гасло в толще мускулатуры, плечевой пояс следовало связать с осевым скелетом. Такая связь соединила плечевой пояс с черепом — крупной и жесткой структурой в передней части тела (рис. 5). Однако на суше связь плечевого пояса и затылочного отдела черепа будет только мешать. Представьте себе, что находящаяся на земле рыба приподнимает тело, выпрямляя ноги. Из-за гибкости ее позвоночника она просто задерет голову и хвост кверху. Чтобы избежать этого, плечевой пояс отсоединяется от головы и смещается назад. При этом вес грудного отдела компенсируется весом тяжелой головы. А для компенсации веса брюшного отдела сзади развивается массивный хвост. Именно так появился мускулистый хвост, являющийся характерной чертой внешнего облика и типичных земноводных (хвостатых), и даже рептилий (ящериц и крокодилов)8.
Первые по-настоящему наземные животные имели плечевой пояс, расположенный в толще мышц. Лишь позже, в эволюционной линии, ведущей к нам, плечевой пояс оказался соединен с грудной клеткой, а через нее — и с позвоночником.
А эволюционную роль, которую сыграла возможность приподнять тело на конечностях, трудно переоценить. Как мы уже отметили, на суше рыбы-четвероногие осуществляли газообмен через кожу. Но если они приподнимали тело, то разблокировали легкие и могли включить намного более эффективное легочное дыхание! Легкие и ноги, независимо приобретенные как приспособления к жизни в воде, в сочетании друг с другом оказались чрезвычайно полезными для наземного существования.
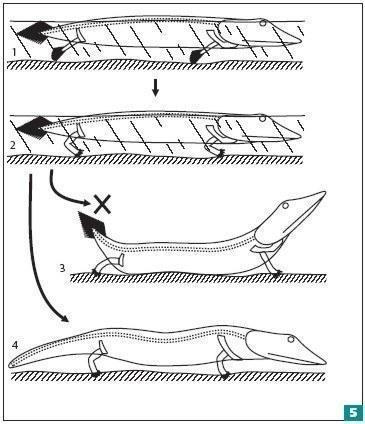
Связь плечевого пояса и черепа для животного с гибким позвоночником окажется на суше крайне неудобной. Самые первые посетители суши защищались от ситуации, показанной на третьей картинке, благодаря упрочнению грудной клетки
Эволюционный куст
Не вдаваясь в дальнейшее рассмотрение анатомо-физиологических преобразований, связанных с освоением суши, посмотрим, какие именно животные реализовали их. И тут мы столкнемся с парадоксом. Если бы мы знали только ихтиостегу, нам было бы просто сказать: вот она, переходная форма! А мы знаем уже много видов, находящихся на переломном этапе эволюции. Как ни крути, они не выстраиваются в единую последовательность. Рассматривая известные нам виды, мы видим мозаику из архаичных и продвинутых черт. Например, у ихтиостеги была еще рыбья, сжатая с боков форма тела, которая обеспечивалась благодаря очень прочной грудной клетке. А ее современник тулерпетон (Tulerpeton, рис. 6) имел намного более наземный вид, однако сохранял связь плечевого пояса с черепом — свидетельство того, что в его истории не было периода наземной жизни. Значит, существовало несколько эволюционных линий, эволюционирующих в направлении наземности рыб, и несколько — в направлении первых четвероногих. Точно выстроить ту траекторию развития, которая привела к нам, не удается. В параллельном развитии нет ничего удивительного: млекопитающие и цветковые растения, птицы и рептилии тоже развивались параллельными эволюционными ветвями.
Более того, очевидно, что скорость эволюции от рыб к четвероногим менялась и главные преобразования происходили быстро. Вспомним еще одну теорию — прерывистого равновесия, появившуюся в конце XX века. Она утверждает, что в истории видов периоды быстрых изменений чередуются с длительными промежутками стабильности. Вероятно, быстрые изменения связаны с неустойчивыми местообитаниями и небольшими изменчивыми популяциями. Шансов найти в летописи остатки животных из таких популяций крайне малы — мы встречаем в основном массовые формы.

Тулерпетон9
Нам были известны весьма продвинутые рыбы — например, пандерихт (Panderichthys, рис. 4), и очень примитивные четвероногие, как уже упомянутая акантостега. Однако не следует думать, что акантостега — потомок пандерихта. Кроме тех признаков, которые мы обсудили, для реконструкции эволюции важны многие другие — например, ход упрощения крыши черепа. У рыб она состоит из многих костей. У четвероногих одни из этих костей исчезают, другие сливаются друг с другом. Так вот, пандерихт успел избавиться от тех костей в крыше черепа, которые еще сохранялись у акантостеги…
Вот в этот-то разрыв (не генеалогической, а сравнительно-анатомической) преемственности от пандерихта к акантостеге и «легла» новая находка — тиктаалик.
Новый камешек в старую мозаику
Название тиктаалика происходит из языка эскимосов, ведь найден он в верхнедевонских отложениях северной Канады. Это крокодилообразное существо, продвинувшееся по пути приспособления к жизни на суше еще дальше, чем пандерихт. Почти-почти четвероногое, но еще с плавниками, а не с ногами. Как и у акантостеги, гибкость тела уменьшается благодаря мощной грудной клетке, а разрыв связи плечевого пояса и черепа свидетельствует о неплохой приспособленности к перемещению по суше.
Как оценить эту находку? Как очень интересную. Не надо только говорить, что она доказала происхождение четвероногих от рыб, как это делают многие СМИ, — эта новость изрядно запоздала. Не надо также думать, что тиктаалик — наш предок. Это было бы маловероятно, и этому противоречат некоторые особенности его анатомии.
В эволюции раз за разом повторялся один поучительный сценарий. Новую адаптивную зону вначале занимали животные, которые преобразовались относительно неглубоко, а потом их вытесняла группа, которая эволюционировала медленнее, но зато приобретала более удачную конструкцию. Рыбы, которые вышли на берег с плавниками, не имели эволюционного будущего. Они смогли успешно ползать и питаться на суше, но не смогли включить свои легкие. «Линию фронта» прорвали те, кто претерпели более глубокие преобразования во время жизни на мелководье и вышли на сушу, вооруженные конечностями. Сочетание ног и легких стало ключом к освоению суши. Среди них тоже был свой авангард и свой основной отряд. Те, кто накачивал воздух в легкие с помощью горлового насоса (вспомните, как двигает горлом лягушка), дали начало всяческим амфибиям. Различные рептилии произошли от животных, вентилировавших легкие благодаря изменению объема грудной клетки. Наши предки принадлежали именно к этой группе.
Экономна ли эволюция?
Изложенные выше взгляды не являются общепринятыми. Более того, если вы зайдете на западные сайты, то увидите кладограмы (реконструкции родственных отношений, возникающих в результате эволюции), где пандерихт, акантостега и тулерпетон выстраиваются в одну линию (см., например, tolweb.org/Terrestrial_vertebrates). Скоро на таких кладограмах займет свое место и тиктаалик. Получается, его все-таки будут рассматривать как нашего предка или его ближайшего родственника? Дело тут в методе реконструкции эволюции.
Большинство зарубежных авторов используют «объективные» методы кладистики (филогенетической систематики), якобы уменьшающие влияние точки зрения исследователя на научный результат. Мы описываем известные нам организмы по имеющимся признакам и заставляем компьютерную программу построить последовательность изменений, которая соединила бы известные точки самым экономным образом. Предполагается, что количество эволюционных изменений (приобретений или потерь каких-то признаков) должно быть минимальным. Параллельная эволюция в нескольких ветвях оказывается менее экономной, чем один ствол с разветвлениями (рис. 7). Принцип экономии (парсимонии), вырастающий из средневековой бритвы Оккама10, оказывается для многих ученых основой всей современной науки. Утрируя, скажем, что кладист предположит, будто тулерпетон приобрел множество черт четвероногих вместе с их общим стволом и затем почему-то восстановил связь плечевого пояса с черепом11.
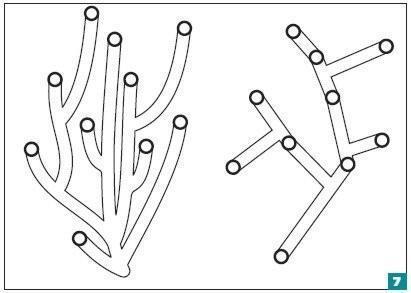
Условный пример «экономной» и «неэкономной» родословных. Нам известно 10 видов, показанных на рисунке кружками. Возможно, если бы мы точно знали, как шла эволюция этой группы, то построили бы левое, «неэкономное» дерево. Однако если мы стремимся выбрать наиболее экономный вариант, у нас получится такое дерево, как справа
Противоположная (консервативная) точка зрения не может доказать свою правоту с помощью компьютерных программ и вынуждена обходиться логикой и способностью к рассуждениям. С этой точки зрения, изучая эволюцию какой-то группы, следует понять, как и почему она происходила, и создать «эволюционный нарратив» — анализ причин и этапов эволюционного процесса. Мы не будем здесь подробно сравнивать эти подходы, но укажем на два обстоятельства.
Первое. Написать статью или прочитать лекцию, которые будут способствовать пониманию (а не просто запоминанию) хода эволюции, можно лишь на основе нарративного подхода. Результатом этого является не только обладание фактами, а их увязывание в единый смысловой узел, компактизирующий знание (в силу его внутренней связанности) и позволяющий делать прогнозы.
Второе. Те кладистические процедуры, которые опирались на действительно богатый набор данных (например, о химической эволюции нуклеиновых кислот и белков), доказывают, что эволюция сплошь и рядом идет неэкономно, параллельными путями.
Зачем уделять столько внимания тиктаалику? Во-первых, освоение суши — захватывающая история, а последствия ее перипетий отражаются в наших собственных телах. Во-вторых, чтобы на ее примере показать спорные вопросы теории эволюции. К примеру, креационисты не верят, что одна форма жизни может превратиться в другую. Ортодоксальные сторонники синтетической теории эволюции отрицают, что в эволюции было что-либо, кроме изменения генных частот в популяциях. От этих и других форм догматизма нет лучшего лекарства, чем изучение истории жизни. Действительность намного интересней наших представлений о ней!
1Edward B. Daeschler, Neil H. Shubin, Farish A. Jenkins Jr. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan // Nature. 2006. V. 440. P. 757-763; www.nature.com/nature/journal/v440/n7085/fig_tab/nature04639_F6.html#fig....
2 Перемещение Луны над земной поверхностью вызывает приливы-отливы и деформацию земной коры. На это тратится часть энергии вращения Земли. В результате скорость вращения замедляется.
3 Двоякодышашие и многопёры.
4 Анабасы, илистые прыгуны, мелкие сомы и другие.
5 Приведенные рассуждения объясняют многие особенности хода эволюции. Первый пример. В кембрийском периоде, когда животные со сложным планом строения только появились, за геологически короткое время возникло множество типов, отличавшихся планами строения. Со временем развитие представителей удачных типов оказалось хорошо зарегулированным. Теперь не очень крупное изменение строения требует больше времени, чем становление самого типа на заре эволюции многоклеточных животных. Второй пример — возрастание у продвинутых групп животных роли приспособления за счет software (поведения), а не hardware (изменения плана строения тела).
6 Воробьева Э.И. Проблема происхождения наземных позвоночных. — М.: Наука, 1992. — 344 с.
7 Шабанов Д.А., Шабанова Г.В., Шапаренко С.О. Біологія. Підручник для 7 класу загальноосвітних навчальних закладiв. — Х.: Торсінг, 2003. — 320 с.
8 Лишь бесхвостые амфибии, млекопитающие и птицы, которые приобрели жесткий позвоночный столб, а также черепахи, которые укрепили его панцирем, смогли отказаться от хвоста-противовеса. Развитие позвоночного столба к моменту освоения суши было уже хорошо зарегулировано, и для того, чтобы сделать его по-настоящему жестким, ушло много времени.
9 9 Воробьева Э.И., цит. соч.
10 Сущности не следует умножать без необходимости. Цитируя Оккама, обычно забывают, что он требовал и не уменьшать количество сущностей без необходимости.
11 В рамках науки, которая считает себя строго объективной, вопросы «почему» и «зачем» часто игнорируются.
Д. Шабанов. Тиктаалик и эволюция // Северо-Запад. — Компьютерра, М., 2006. – № 18 (638). — С. XII-XV