| ← | Дмитрий Шабанов | → |
| "Экологические задачи" для студентов и школьников | Альтруизм и парадокс Симпсона | Подпорки альтруизма |
| Колонка в КомпьютерреOnline #49 | Колонка в КомпьютерреOnline #50 | Колонка в КомпьютерреOnline #51 |
В спорах по поводу моих колонок кто-то из борцов с наукой утверждал, что для эволюционной биологии необъяснимо альтруистическое поведение людей. Этот добрый человек не знал, что изучение эволюции альтруизма – одна из быстро развивающихся отраслей эволюционной психологии. Много интересного по этому поводу написано Александром Марковым. Хороший обзор этой темы содержится в его недавнем двухтомнике об эволюции человека. Не пересказывая то, что можно прочитать у Маркова, я все же посвящу пару колонок этой теме.
Еще во времена Дарвина появилась ложная трактовка дарвинизма, как войны всех против всех. Из числа тех, кто считал, что Дарвин недооценивал взаимопомощь, самым ярким был князь Петр Алексеевич Кропоткин, географ и теоретик анархизма. Кропоткинская «Взаимопомощь как фактор эволюции», написанная в 1902 г., не содержит альтернативы или опровержения дарвинизма, она лишь иначе расставляет акценты, уделяя большее внимание гармонии в отношениях между особями. С Кропоткиным нельзя не согласиться в том, что разные формы взаимопомощи широко распространены в нашем мире. В ходе эволюции они возникали неоднократно.
Отрицают ли дарвинизм и более поздняя эволюционная биология взаимопомощь? Конечно, нет. Они рассматривают возникновение приспособительных признаков, в том числе, и поведенческих. В разных условиях наибольший успех обеспечивают разные формы поведения – иногда чисто эгоистические, иногда – антагонистические, но довольно часто – и альтруистические.
Как альтруистическое поведение могло закрепиться в ходе эволюции? В конце XIX-начале XX века его объясняли отбором между группами (видами, популяциями, стадами и др.) особей. Казалось естественным, что эволюция должна была поддерживать объединенные взаимопомощью группы.
С распространением СТЭ объяснять становление альтруизма стало сложнее. Эволюцию поведения стали представлять, как распространение генов, вызывающих определенные формы поведения. Может ли распространиться ген, который способствует поведению, уменьшающему долю носителей этого гена в следующих поколениях? Нет! А разве альтруизм (принесение своих интересов в жертву интересам сородичей) не приведет к такому эффекту? Некоторые наиболее ретивые сторонники СТЭ решили, что возможность группового отбора опровергнута в принципе. Они часто ссылаются на Ричарда Докинза. К счастью, Докинз не отрицает групповой отбор как таковой. В «Расширенном фенотипе» он лишь утверждает, что групповой отбор существенно менее эффективен, чем отбор особей.
Сразу скажу, что существование группового отбора доказано. Для примера сошлюсь на сделанный Марковым пересказ исследования, в котором было доказано наличие отбора видов. Пасленовые неоднократно переходили от перекрестного опыления к самоопылению. Во фрагментах эволюционного дерева пасленовых, где практиковалось самоопыление, и возникновение новых видов, и их вымирание шло быстрее, чем у их перекрестноопыляющихся родственников. Тем не менее, то, насколько групповой отбор распространен и эффективен - предмет споров. Можно ли объяснить альтруизм без группового отбора? Как минимум две формы альтруизма вполне объяснимы на уровне индивидуального отбора.
Родственный альтруизм. Может ли эволюция поддержать поведение самки, при котором она будет жертвовать жизнью ради выводка своих потомков? Вполне. Сторонники СТЭ объясняют это так. Представьте себе ген, вызывающий готовность к жертвенному поведению матери. Если она является носителем такого гена, он с высокой (и вычисляемой) вероятностью оказывается присущ и ее потомкам. Жертвуя собой ради потомства, такая особь при определенных условиях будет распространять ген готовности к самопожертвованию.
Если как следует поскрести реципрокный (взаимный) альтруизм, выяснится, что он, на самом деле, не альтруизм, а расчетливый эгоизм. Речь идет об отношениях «ты – мне, я – тебе». В популяциях, где особи способны узнавать друг друга индивидуально, возможны отношения, при которых особь, к примеру, делится пищей с сородичем, зарабатывая возможность получить долю от добычи соседа, когда тот окажется более удачливым. Индивидуальный отбор будет способствовать распространению в популяции генов, обуславливающих способность помогать ближним, запоминать их репутацию и требовать возвращения «долга» при возможности.
Можно ли объяснить все проявления альтруизма этими механизмами? Вряд ли. Что делает нормальный мужчина, увидев чужого ребенка в опасности? Кидается на помощь, невзирая на опасность для себя. Видимо, такая форма поведения является врожденной, инстинктивно предопределенной. (Реплика в сторону: некоторые читатели испытывают возмущение от утверждения, что человек – животное, и многие особенности его поведения обусловлены эволюционно. Им почему-то кажется, что такое поведение обязательно должно быть «скотским», аморальным. К счастью, многие наши «животно» предопределенные формы поведения неплохо согласуются с требованиями морали). Кажется, для объяснения такого альтруизма нужен групповой отбор.
Тут есть одна проблема. Мне кажется, сложнее объяснить не то, почему возникают группы, состоящие из альтруистов, а то, почему эти группы не разрушаются эгоистами. Представьте себе: в группе, где все особи помогают друг другу, преимущество может получить та, которая будет принимать помощь, не оказывая ее взамен. Такие особи могут оставлять больше потомков, чем альтруисты, и вытеснять их со временем! Почему же альтруисты в нашем мире не перевелись совсем?
Один из ответов на этот вопрос связан с парадоксом Симпсона. Я узнал о нем из статьи Маркова; отсылаю и вас к ней. Это обсуждение статьи американских авторов, создавших экспериментальную модель для демонстрации этого парадокса. Он может проявляться в популяции, которая то подразделяется на субпопуляции, на развитие каждой из которых влияет ее состав, то воссоединяется заново. Вот иллюстрация из упомянутой статьи.
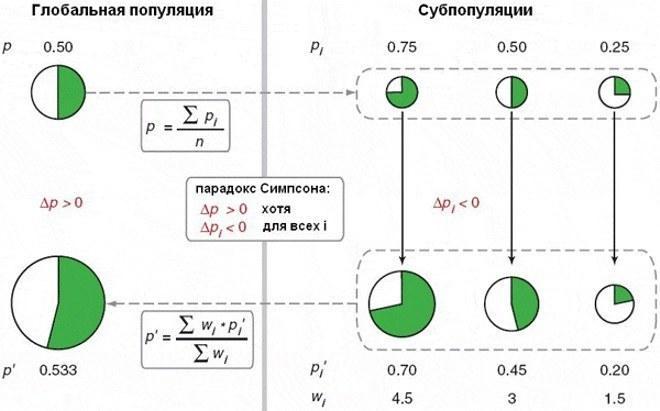
Парадокс Симпсона: в каждой из трех субпопуляций доля альтруистов (показана зелеными секторами) сокращается, а в целом она растет. Причина – в ускоренном росте тех субпопуляций, где выше доля альтруистов
Авторы статьи, которую пересказывает Марков, показали действие парадокса Симпсона на культурах бактерий. Они выращивали смесь из двух штаммов. Одни (альтруисты) производили вещество, включающее механизм защиты от антибиотика, другие (эгоисты) – нет. При совместном «эгоисты» пользовались веществом, произведенным "альтруистами". Колонии, где было много альтруистов, росли быстрее. Хотя в каждой из них доля альтруистов сокращалась, в целом для популяции она росла. Красиво, правда?
Чтобы дать студентам возможность «пощупать» парадокс Симпсона, я сделал иллюстрирующую его модель в Excel. Скачать ее можно тут; увы (возможно – из-за моей программистской неискушенности) она оказалась довольно громоздкой.
Как вы поняли, парадокс Симпсона проявляется, когда популяция то подразделяется на субпопуляции, то объединяется снова. Я решил начать с какого-то простого случая разделения-объединения, и вспомнил о жизненном цикле многих донных морских животных – от коралловых полипов до асцидий.
Представьте себе животных, образующих группы на подходящих для них участках дна. В положенный срок они выбрасывают в воду половые клетки (гаметы). В толще воды развиваются личинки, оседающие на дно. Шансы каждой особи на выживание зависят не только от ее наследственных задатков, но и от ее ближайшего окружения (а свойства этого окружения зависят от наследственности образующих группу особей). Как раз то, что нужно для проверки работы парадокса Симпсона!
Группы таких донных животных могут страдать от хищников. К примеру, при нападении морской звезды какие-то из наших животных могут эгоистично прятаться в грунт, а какие-то – альтруистично (с точки зрения сородичей) нападать на агрессора. Альтруисты будут чаще гибнуть, но зато хищники станут предпочитать группы, где альтруистов немного. Хватит ли этих предпосылок, чтобы парадокс Симпсона поддержал гены альтруизма?
Забегая вперед, скажу, что по мере отладки модели я понял, что различий в смертности в разных группах недостаточно для поддержки альтруистов. В модели возникают субпопуляции, численность которых определяется случайно, от 0 до некой переменной величины. Повышенная доля альтруистов чаще достигается в мелких группах. После того, как смертность сократит численность всех групп, вклад «везунчиков» в общий результат становится малосущественным. Мне пришлось модифицировать модель, добавив в нее размножение внутри субпопуляций. Это реалистичное предположение. Донные животные, о которых мы говорили, от кораллов до асцидий, кроме полового размножения часто используют и бесполое. В хороших условиях удачливая особь почкуется, образуя группу своих генетических копий. Ко времени полового размножения гаметы в воду выпустит не одна особь, а несколько.
Если хотите – скачайте файл Excel и поиграйтесь с моделью сами. Тут я расскажу лишь о нескольких сделанных с ее помощью выводах.
В модели независимо задается влияние аллелей гена A на шансы особи выжить и размножиться внутри той группы, в которую она попала, и уровень смертности и количества потомков, характерных для групп в целом. Если аллель A обеспечивает альтруистическое поведение, его носители должны проигрывать своим конкурентам на индивидуальном уровне. Такой результат показан на рисунке. Скачки на графике связаны с высокой долей случайности при формировании субпопуляций, гибели и размножении особей. При ином запуске модели траектория окажется несколько иной, но конечный результат (искоренение гена альтруизма) вероятнее всего останется тем же самым.
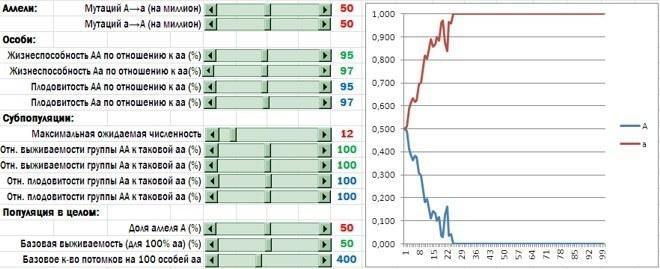
Носители аллеля A проигрывают носителям аллеля a. Этот эффект сильнее выражен у генотипа AA, послабее - у Aa (иными словами, имеет место кодоминирование). Вначале доли аллелей A и a равны, но в течение первых 25 лет отбор удаляет аллель A из популяции
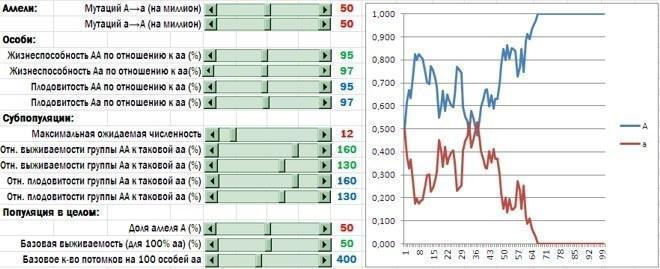
Субпопуляции с высокой долей альтруистов (носителей аллеля A) получили преимущество в выживаемости и плодовитости. Теперь чаще всего в популяции закрепляется аллель альтруизма (хотя на результат конкуренции аллелей сильно влияет случайность)
Что в итоге? Первый вывод я уже назвал. Групповой отбор в пользу альтруистов работает намного эффективнее, когда он проявляется не только в меньшей смертности групп, состоящих из альтруистов, но и в их большей плодовитости.
Второй вывод состоит в том, что групповой отбор пересиливает отбор на уровне особей только тогда, когда он связан с весомыми отличиями между группами. Если особи-альтруисты уступают эгоистам на единицы процентов от уровня их выживаемости и плодовитости, то парадокс Симпсона будет устойчиво проявляться при преимуществах групп альтруистов в десятки процентов.
Третье (картинками не иллюстрирую; хотите – проверьте). Альтруисты чаще побеждают, чем проигрывают, только если субпопуляции оказываются маленькими. Если группы будут достаточно велики, и при их образовании не будет действовать никакой механизм, собирающий альтруистов вместе, доля альтруистов во всех группах будет примерно одинакова. Перевесит эффект отбора против альтруистов внутри субпопуляций. Увы, поскольку группы, в которых может проявиться парадокс Симпсона, невелики и эфемерны, на их эволюцию серьезное влияние оказывает простая случайность.
Итак, одним парадоксом Симпсона закрепление альтруизма в эволюции не объяснишь. Но все ли факторы мы учли?
| ← | Дмитрий Шабанов | → |
| "Экологические задачи" для студентов и школьников | Альтруизм и парадокс Симпсона | Подпорки альтруизма |
| Колонка в КомпьютерреOnline #49 | Колонка в КомпьютерреOnline #50 | Колонка в КомпьютерреOnline #51 |
Коментарі
_
Возможно, отбор благоприятствовал только первым двум из указанных разновидостей альтруизма, родственному и реципрокному. Все остальное - их издержки. Если брать в качестве примеров альтруизм людей, то люди формировались в маленьких родовых группах. Там ген самопожертвования ради группы приносил пользу группе (и самому себе). Но маленькие группы сильно разрослись, а ген остался, заставляя людей жертвовать собой ради своего многомиллионного государства. Сейчас его доля в генофонде должна постепенно снижаться. Наверное, моралисты правы, мы становимся эгоистичнее :)
Это, кстати, в принципе можно проверить. По идее такой необъяснимый альтруизм должен быть характерен только для тех видов, условия обитания которых за последнее время сильно изменились. Скорее всего, не удастся обнаружить вид, обитающий в стабильных условиях, где альтруизм был бы не реципрокным или родственным.
Поскольку мы живем в условиях, которых раньше не было, ...
...все наши свойства сформированы отбором, который можно считать анахроничным. Знаете, как генералы готовятся к прошедшей войне, так свойства организмов вызваны отбором к прошлым условиям.
Но идет ли сейчас отбор против врожденных альтруистов - не знаю. Это не так-то просто установить.
Последняя идея кажется мне настолько сырой и неконкретной, что о ее реализации говорить трудно.
Очепятка
1. Одни (альтруисты) производили вещество, включающее механизм защиты от антибиотика, другие (эгоисты) – нет. При обитании с «альтруистами» «эгоисты» пользовались веществом, произведенным [эгоистами].
Спасибо!
И никто же до сих пор не увидел...