| ← | Дмитрий Шабанов | → |
| Ещё раз о природе этноса, а также о разнообразии надсистем | Как в ходе эволюции возникают адаптивные признаки, или Какую из теорий эволюции подтверждают данные современной генетики? | Краткое изложение эпигенетической теории эволюции, или ЭТЭ для занятых |
| Колонка для Компьютерры #126 | Колонка для Компьютерры #127 | Колонка для Компьютерры #128 |
В последних колонках я не раз обращался к теме ЭТЭ — эпигенетической теории эволюции. Её сторонники (и я в их числе) считают эту теорию серьёзной альтернативой распространённым в эволюционной биологии «по умолчанию» взглядам. СТЭ (синтетическую теорию эволюции, дефолт-взгляды, которые преподают в школе и вузах) давно уже нельзя считать продуктом современной биологии: она на восьмом десятке. За семьдесят с лишним лет после создания этой теории биология узнала едва ли не больше, чем за всю свою предшествовавшую историю. Излагая СТЭ, приходится или возвращаться назад во времени, или серьёзно ретушировать поправками и заплатками старый костяк теории.
Да, я критично настроен к СТЭ, но, пожалуйста, не ищите в этой колонке ни отрицания эволюции, ни скандальных новостей. Речь идёт о проблемах, связанных с развитием живой и сложной науки — эволюционной биологии. Увы, в наше время внутринаучные дискуссии (как противостояние СТЭ и ЭТЭ) теряются за шумными нападками на науку со стороны лженауки, всяких «научных креационизмов». Здесь я не буду размениваться на обсуждение аргументов профессиональных отрицателей факта эволюции, а поговорю о спорах относительно её механизмов.
Я весьма скептически отношусь к цитированию Википедии в научных спорах, но тут не могу удержаться. Вот как неизвестные мне авторы этой энциклопедии определяют СТЭ. Выделение в цитате моё: я хочу, чтобы вы оценили, как сочетаются друг с другом подчёркнутые слова в тексте сторонника обсуждаемой теории.
"Синтетическая теория эволюции (также современный эволюционный синтез) — современная эволюционная теория, которая является синтезом различных дисциплин, прежде всего генетики и дарвинизма. <…> Синтетическая теория в её нынешнем виде образовалась в результате переосмысления ряда положений классического дарвинизма с позиций генетики начала XX века".
История становления СТЭ началась в 1926 году (году публикации статьи С. С. Четверикова), а закончилась в 1942-м, с публикацией книги Джулиана Хаксли. Творцы этой теории вызывают у меня искреннее уважение. Те упрощения, которые были приняты ими семь с лишним десятилетий назад, способствовали росту понимания и доказали (для своего времени) свою полезность. Но те, кто до сих пор считают их достижения «современным» эволюционным синтезом, демонстрируют просто леность мысли.
Очень коротко, СТЭ — это представление об эволюции как результате изменения частот аллелей (вариантов генов), вызываемого естественным отбором организмов по признакам, которые определяются этими аллелями. Помните опыты Менделя в школьном изложении? Один аллель гороха вызывает зелёную окраску семян, другой — жёлтую. Сохраняя в ряду поколений преимущественно зелёные семена, мы повысим частоту «зелёного» аллеля и сдвинем равновесие в сторону преобладания этого признака…
ЭТЭ рассматривает свойства организма как результат индивидуального развития (онтогенеза). Популяционная норма может развиваться на весьма разной генетической основе вследствие длительного отбора на устойчивость её воспроизведения (стабилизирующего отбора). При изменении условий отбор начинает дестабилизировать прежнюю норму, а затем стабилизировать новую, которая возникает как ответ на изменения среды. Хотите разобраться в этой теории получше — читайте её краткое описание в одной из прежних колонок, а ещё лучше — разберитесь в работах М. А. Шишкина, А. П. Расницына и А. С. Раутиана, которые выложены у меня на сайте.
Ключевые для ЭТЭ идеи были высказаны И. И. Шмальгаузеном и К. Х. Уоддингтоном во второй половине прошлого века. В отдельную теорию эти взгляды сложились в работах московского палеонтолога Михаила Александровича Шишкина, опубликованных начиная с 1984 года (относительно полное изложение — в статье 1987 года).
Итак, ЭТЭ подходит к тридцатилетнему рубежу. Казалось бы, возраст зрелости… Увы, у неё относительно немного сторонников. Одна из причин состоит в том, что для того, чтобы её понять, надо посмотреть на вещи, описанные в школьных учебниках, под непривычным углом зрения. Многие на это неспособны и компенсируют недостаток своего понимания с помощью издёвки (как, к примеру, тут) или странных обвинений (хотел привести примеры, описал кое-что обидное… и убрал, чтоб не раздувать ссоры).
Одной из специфических причин слабой известности ЭТЭ является то, что сторонники СТЭ любят обсуждать критику своих взглядов с точек зрения альтернативных теорий эволюции, являющихся её «сверстниками», то есть имеющих тот же или даже больший возраст. Процитированная мной статья Википедии относительно успешно «отбивается» от неодарвинизма (понимая его как вейсманизм, перешагнувший столетний юбилей), мутационизма (сальтационизма, столетней во возрасту теории, сводя к ней и сорокалетнего «юношу» пунктуализм), и упоминает номогенез (празднующий свою девяностолетнюю годовщину). Получается довольно убедительно, если не знать, что обсуждаемые проблемы представляют в основном исторический интерес. ЭТЭ принято замалчивать, если не затыкать её сторонникам рот. Вот, посмотрите. На сайте А. В. Маркова в подборке материалов по ЭТЭ висит ссылка (непрямая) на сетевую дискуссию, в которую я когда-то ввязался (с 13-й страницы). Дискуссия связана со статьёй М. А. Шишкина, опубликованной в 2006 году в журнале «Онтогенез» (саму статью можно прочитать и у Маркова, и на моем сайте). Меня искренне поразило, что ряд сторонников СТЭ настаивает на запрете публикации статей, столь сильно не соответствующих их взглядам.
Конечно, СТЭ очень неоднородна. Авторы, принимающие её на словах, могут отстаивать весьма разные взгляды, которые, как мне кажется, не могут совмещаться в одной теории. Одной из современных версий СТЭ (может быть, школой внутри СТЭ, может быть, «дочерней» теорией, а может, даже и альтернативой) является концепция Evo-Devo.
Evo-Devo (evolution-development, то есть «эволюция-развитие»), или эволюционная биология развития — относительно молодое направление исследований, сложившееся в конце прошлого века, но уходящее своими «корнями» ещё в XIX столетие. Основное внимание в Evo-Devo уделяется изменению онтогенеза в эволюции. Неплохое представление об этом направлении даёт caenogenesis, ЖЖ Сергея Ястребова (чтобы разобраться в материале, записи в этом блоге лучше читать от самых старых к более новым).
ЭТЭ часто сравнивают с Evo-Devo, и это не случайно. И тот и другой подход рассматривают эволюцию как перестройку онтогенеза. Однако классики ЭТЭ категорически не согласны со сближением их детища и Evo-Devo. Глубокие различия этих подходов перевешивают отдельные черты их сходства. Главное различие состоит в том, как они рассматривают возникновение адаптивных, приспособительных признаков. По этому вопросу подход Evo-Devo не отличается от такового у классической версии СТЭ. Вот, посмотрите на схемы. В них показаны причинно-следственные цепочки, приводящие к элементарному эволюционному событию с точек зрения сравниваемых концепций.
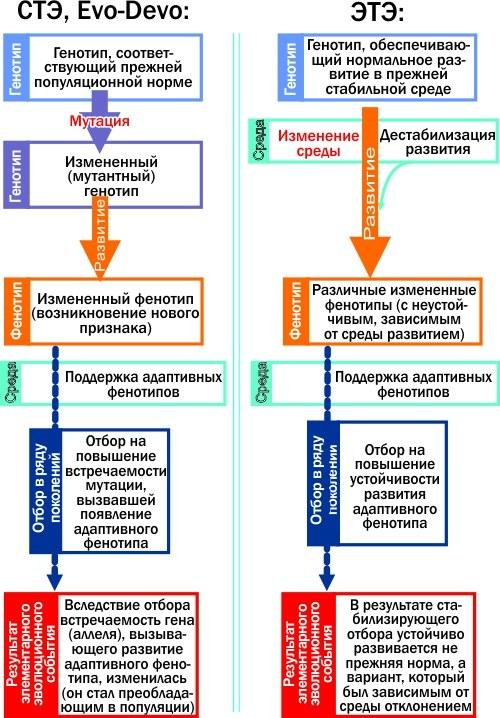
Ключевые этапы элементарного эволюционного изменения согласно СТЭ и Evo-Devo, с одной стороны, и ЭТЭ, с другой. В левой колонке причиной приспособительного признака является создающая его мутация. В правой — дестабилизация прежней нормы и целостный ответ на неё организма (на который влияет созданный предшествующим отбором целесообразный комплекс внутренних взаимосвязей)
И СТЭ, и Evo-Devo видят причину появления новых признаков в мутациях. Классическая СТЭ в основном рассматривает возникновение новых аллелей, характеризующихся относительно небольшим эффектом. Для Evo-Devo характерно внимание к мутациям ключевых регуляторных генов (с этой точки зрения эволюционная биология развития продолжает традиции сальтационизма, идущие от таких классиков генетики, как Х. Де Фриз и Р. Гольдшмидт). Если мутация окажется полезной, отбор будет преимущественно сохранять её носителей.
С точки зрения ЭТЭ все выглядит иначе. Мутация практически никогда не может создать сложный новый признак. Его создаёт изменение онтогенеза, после того как изменение среды делает неустойчивым нормальное развитие. А затем, если поколение за поколением отбор будет поддерживать новый признак, произойдёт то, что К. Х. Уоддингтон называл «генетической ассимиляцией»: генотип изменится так, что устойчивым окажется тот вариант развития, который был востребован средой.
Слева на схеме мутация является причиной признака (в том числе изменения онтогенеза), справа первичным является изменение онтогенеза, а уже потом отбор создаёт генные механизмы, способствующие его устойчивому осуществлению.
Различие понятно? Тогда посмотрим, какую из точек зрения подтверждают последние результаты сравнительной геномики.
Сформулирую прогнозы.
СТЭ и Evo-Devo. Если гены являются причиной признаков, одновременно (в эволюционных масштабах) с появлением соответствующих генов должны проявляться и их эффекты.
ЭТЭ. Если гены лишь перестраиваются, чтобы быть переключателями изменений онтогенеза, происходящих в ответ на изменения среды, важные эволюционные нововведения сплошь и рядом должны опираться на давно существовавшие гены.
Далеко ходить не буду, процитирую несколько свежайших обзоров последних достижений, выложенных на «Элементах». Сергей Ястребов (тот самый, caenogenesis) пересказывает следующее.
У губок, самых-самых простых многоклеточных животных, нет ни нервной системы, ни клеток, специализирующихся в роли органов чувств. Тем не менее у губок есть гены (и даже генные сети, пусть даже состоящие всего из двух взаимодействующих друг с другом генов), которые управляют развитием глаз. Очевидно, что гены, которые кажутся кому-то «причиной» развития глаз, появились несравнимо раньше.
Может, в регуляции развития отдельных органов возможно выстраивание генных сетей «под готовый результат», а общая конструкция организма определяется иначе? Нет. Гомеозисный (работающий в определённых частях тела, то есть определяющий план строения организма) ген Brachyury определяет у нас и других хордовых самую нашу характерную особенность — образование хорды. Можно было поверить, что он характерен не только для хордовых, а и для всей огромной группы заднежгутиковых эукариот (куда относятся все-все животные, грибы и воротничковые жгутиконосцы)? Некоторые грибы этот ген потеряли, но иные — сохранили! Естественно, никакой хорды у них нет и в помине. Просто, когда у наших предков появилась хорда, они использовали для включения её развития один из генов, который имелся у них давным-давно и который сохранился у множества их родственников!
Почитайте Ястребова - и узнаете, что если в зародыш лягушки с заблокированным геном, который якобы является «причиной» развития хорды, ввести ген одноклеточного капсаспоры (у которой хорды отродясь не было и быть не может), этот ген «заработает» в лягушке и обеспечит начало формирования столь важного органа.
А ещё вы можете узнать, что многие черты сходства разных групп организмов в работе их Hox-генов, самой важной группы регуляторов плана строения, не унаследованы от общих предков, а вырабатывались независимо.
Ещё один интересный результат излагает Елена Наймарк. Оказывается, разные организмы, вырабатывая сходные приспособления, могут использовать для них одни и те же гены. В обсуждаемой работе это показано на примере развития способности к эхолокации у дельфинов и у летучих мышей — не таких уж близких родственников. Эхолокацию развивали независимо, а гены используют одни и те же!
Таким образом, эволюционные нововведения не являются следствием мутаций (хоть и используют их). Отбор выстраивает механизм генной регуляции под такую перестройку онтогенеза, которая «затребована» средой. Эта мысль кажется вам невероятной? А можно я, чтобы усилить ваш когнитивный диссонанс, процитирую И. И. Шмальгаузена - авторитета, которого сторонники СТЭ считают «своим»? Это из статьи «Стабилизирующий отбор и эволюция индивидуального развития».
"Мутации вводятся все более в русло направленных изменений. В основном они используются для наследственного “фиксирования” индивидуальных приспособлений, приобретающих постоянное значение в данных условиях существования. Вместе с тем усложняется система внутренних факторов развития, всё более занимающих место “недостаточно надёжных” внешних факторов, основные морфогенетические процессы защищаются всё более сложной системой регуляций. Благодаря регуляторным механизмам создаётся более прочная основа не только для индивидуальных онтогенезов, но и для дальнейшего процесса исторического образования организмов, и вся эволюция приобретает характер устойчивого движения". И. И. Шмальгаузен
К сожалению, эти мысли Шмальгаузена просто игнорируются сторонниками СТЭ (впрочем, и сам Шмальгаузен временами сам себе противоречил).
СТЭ пытается объяснить онтогенез на уровне отдельных генов. Чем дальше уходит в своём развитии генетика развития, тем более сложной становится эта задача. Сказанное мной не означает, что онтогенез не опирается на гены. Опирается, но на их уровне он необъясним. Приведу аналогию. Свойства газов можно изучать на уровне их отдельных молекул и на уровне газа как физического тела. Гены и их эффекты взаимодействуют намного сложнее, чем молекулы газа. Поэтому взаимодействие влияющих на онтогенез причин намного сложнее описать «на языке» отдельных генов, чем, например, описать цикл Карно «на языке» отдельных молекул.
Итак. Критики ЭТЭ упрекают её (увы, в большой мере справедливо) за избыточную теоретичность. Но ситуация в науке меняется: сейчас благодаря успехам сравнительной геномики получен огромный массив данных о том, как меняются гены организмов по мере их эволюции. Пока этот массив изучают в основном для реконструкции филогенеза (определения «траекторий» эволюции), но его анализ может дать много ценной информации и о механизмах регистрируемых изменений.
Означает ли то, что я написал, недоверие к результатам, полученным в рамках концепции Evo-Devo? Нет, конечно же, речь идёт о чрезвычайно ценном материале! Просто, по моему мнению, для её сторонников настало время переоценить ценности и отказаться от упрощённой логики СТЭ.
…
Интересно, а как то, что я здесь написал, будет восприниматься через десять и через двадцать лет?
| ← | Дмитрий Шабанов | → |
| Ещё раз о природе этноса, а также о разнообразии надсистем | Как в ходе эволюции возникают адаптивные признаки, или Какую из теорий эволюции подтверждают данные современной генетики? | Краткое изложение эпигенетической теории эволюции, или ЭТЭ для занятых |
| Колонка для Компьютерры #126 | Колонка для Компьютерры #127 | Колонка для Компьютерры #128 |
Коментарі
Комментарий А.В. Маркова
На следующий день после публикации этой колонки я получил критический комментарий от Александра Владимировича Маркова, которому очень благодарен. Этот комментарий требует серьезного обсуждения, и поэтому я скопирую его сюда.
Дмитрий, спасибо за ссылку. Прочел. Очень интересно и полезно.
Мне кажется, вы все-таки слишком примитивизируете "противника", т.е. современные б-м общепринятые взгляды на эволюцию, которые вы называете "СТЭ".
Рассмотрим ваши прогнозы:
Сформулирую прогнозы.
СТЭ и Evo-Devo. Если гены являются причиной признаков, одновременно (в эволюционных масштабах) с появлением соответствующих генов должны проявляться и их эффекты.
ЭТЭ. Если гены лишь перестраиваются, чтобы быть переключателями изменений онтогенеза, происходящих в ответ на изменения среды, важные эволюционные нововведения сплошь и рядом должны опираться на давно существовавшие гены.
Мне кажется, неправильно говорить, что из СТЭ следует такой прогноз. Так многим раньше казалось, это правда, но сейчас установлено, что многие гены - ключевые регуляторы развития животных крайне консервативны (по своим кодирующим последовательностям) и удивительно легко приобретают новые функции в онтогенезе (путем изменения системы межгенных взаимодействий). Вопрос - почему это так? Потому что верна ЭТЭ и все новшества в эволюции возникают "по Шишкину"? Или есть другое объяснение? Конечно, есть. И оно вполне в духе общепринятых взглядов.
Новые функции у этих генов появляются очень легко, потому что они - "профессиональные переключатели", и им все равно, что переключать. Чтобы какой-то дополнительный ген попал под их управление, достаточно, чтобы у этого гена появился новый сайт связывания ТФ, а это высоковероятная вещь. Часто новые энхансеры возникают из-за вставок МГЭ. Прекрасный недавний пример: http://elementy.ru/
Ген sox2 никогда раньше не занимался регуляцией экспрессии пролин-дегидрогеназы в мозге. Но встроился вирус перед геном PRODH - и появилась такая связь. Теперь у гена sox2, вовсе не страдавшего от недостатка важных функций, появилась еще одна, новая: он теперь отвечает за повышенную продукцию пролин-дегидрогеназы в мозге человека, что с большой вероятностью как-то влияет на работу мозга и было полезно нашим предкам.
Отсюда - многофункциональность генов - регуляторов развития, а из многофункциональности следует консерватизм белок-кодирующей части: если такой белок изменить, слишком много разных последствий одновременно будет, поэтому такая мутация практически наверняка окажется вредной.
Предполагать "шишкинский" механизм в развитии этого новшества не вижу оснований. Разумеется, возможно, что итоговое полезное свойство - допустим, чисто для примера, что это была улучшенная эпизодическая память - имело до этого события какую-то изменчивость в популяции гоминид. Особи с новой мутацией (под мутацией в данном случае понимается встроенный вирус) с большей вероятностью оказывались обладателями хорошей памяти (хотя и до мутации у некоторых особей была хорошая память). Может быть даже, что мутация поначалу имела невысокую пенетрантность - не у всех людей с этой мутацией память оказывалась лучше, чем была бы без мутации. А потом шел отбор на "стабилизацию" данного фенотипического проявления этой мутации, т.е. хорошой памяти - подбирались гены-модификаторы, которые усиливали изначально неустойчивый фенотипический эффект встройки вируса. Но все это - нормальные процессы, описываемые в рамках общепринятых представлений, без необходимости "выкидывать СТЭ на помойку" и водружать на опустевший пьедестал ЭТЭ. ЭТЭ - не альтернатива, а занятное дополнительное соображение, "а вот еще и так бывает, вот еще и это надо учитывать".
Еще один пример - появление узора на крыльях D. guttifera в результате появления у гена yellow энхансера, активирующего ген в ответ на присутствие морфогена wingless: http://elementy.ru/
А по ЭТЭ как появился этот узор? Произошел катаклизм, развитие дестабилизировалось, и у некоторых мух с дестабилизированным развитием вдруг обнаружились пятнышки на жилках крыльев, а затем отбор стал эти пятнышки постепенно стабилизировать? Мне такой сценарий представляется менее вероятным.
Сколько вы ни мучайте дрозофилу без пятнышек, каким стрессам ни подвергайте, не станут у нее крылья от этого пятнистыми. А вставьте ей энхансер vs перед геном yellow - и вуаля, вот вам муха с пятнистыми крыльями.
Об "отдельных генах"
Спасибо за очень интересную <для биологов> колонку! Я с её посылами не очень согласен, но прочитал с большим удовольствием.
А.В. Марков Вам всё правильно пишет. +1.
И тут вот ещё что... Вы от колонки к колонке повторяете, что строение организмов, их онтогенез, их эволюцию и т.п. нельзя содержательно объяснить "на уровне отдельных генов". Уже и дилемму Холдейна вспомнили, и с молекулами газа в цикле Карно сравнили.
Возразить по существу тут, кстати, совершенно нечего. Пояснить сложные процессы развития работой отдельно взятых генов действительно едва ли возможно. Но откуда Вы взяли, что та же Evo-Devo пытается разбирать онтогенез и эволюцию "на уровне отдельных генов"? По-моему, в современных работах такого нет и близко. Напротив, многие современные классики (хоть Э. Дэвидсон, хоть А. Уилкинс, хоть прочие) рассматривают развитие с позиции работы довольно сложных, иерархически организованных генных сетей. И на языке генных сетей онтогенез как раз вполне объясним. И изменения онтогенеза (то есть эволюция) - тоже. Не знаю, как Вас, а меня лично объяснения, приводимые в статьях Дэвидсона за последние 10 лет, вполне устраивают. Там есть и общие теоретические выкладки и конкретные примеры. И логика там не такая уж "упрощённая". И рассуждения Шмальгаузена, и все новые примеры, описанные в новостях на "Элементах", прекрасно в эту логику вписываются.
Да, взаимоотношения между геномом (геномами) и условиями среды, дающие на выходе фенотип, - очень сложны. Вы и сами это признаёте. Но при этом реконструировать развитие на языке генетики развития - реально. А почему нет? Лет через 10-20, я думаю, в этом направлении будет очень большой прогресс.
Принципиальная возможность изменения онтогенеза без закрепления в генах, классической генетикой развития (и тем более Evo-Devo), кстати, тоже не отвергается. Та же концепция "фенокопий" - это ж по сути оно и есть. Видимо, часто с таких развивающихся в определённых условиях среды "фенокопий" и начинаются эволюционные изменения. Но потом-то всё равно генные сети перестраиваются. В чём здесь принципиальное противоречие между СТЭ и ЭТЭ, я понять не могу. Иными словами, есть ли какой-то особый путь ЭТЭ, отличный от изучения генных сетей в рамках Evo-Devo? А если да - в чём он, интересно, состоит?
Ну, разговор о генных сетях не так-то далек от разговора о генах
Я, кстати, совершенно не собираюсь опровергнуть Evo-Devo, я только против трактовки ее результатов, как "изучения генов и генных сетей, изменения которых являются причиной перестроек онтогенеза". Что такое "фенокопии", честно говоря, вообще не могу понять. Видимо, это слово, которое отражает уверенность исследователя в линейной логике причинно-следственных цепочек. Анализируя неразложимое переплетение эффектов наследственных задатков, средовых воздействий и случайностей, человек, который использует слово "фенокопия" может сказать: вот в этом случае причиной проявления признака является "фенокопия", а вот в этом - "мутация".
"Фенокопия" - это такое старое, не вполне приличное слово.
Придумал его Гольдшмидт чёрт знает когда. Сейчас оно вышло из моды и уже не особо в ходу. Это слово раньше относили к тому же феномену, на который сейчас смотрят "другими глазами" и в свете проблем эволюции именуют кто "генокопированием модификаций", кто "генетической ассимиляцией преобразований формы", кто "воплощениями эффекта Болдуина", а кто ещё как-то.
У меня сложилось впечатление, что все эти разные слова про то, что к одному и тому же каскаду онтогенетических преобразований (читайте - запуску батареи "низкораноговых" <"рабочих"> генов дифференцировки) могут вести разные пути: короткие - почти напрямую от условий среды и длинные - через большой набор "высокоранговых" регуляторных генов. То есть непосредственная причина проявления признака у "мутанта" и "фенокопии" может и не быть такой уж различной. Различается размер и сложность регуляторной надстройки.
Вообще, конечно, Вы правы - в том, что процессы развития мы ещё довольно слабо понимаем. Ну я - так точно!
P.S. А про лекцию в институте Богомольца могли бы и пораньше предупредить - я бы тогда из Киева в пятницу не уезжал.
И ещё - не помню, писали ли Вы уже об Аристотеле
Этот дядька, поговаривают, написал про четыре типа причинности: материальная, формальная, движущая и целевая. Мне кажется, что спорах о причинах развития признаков в онтогенезе, непонимание между сторонами часто бывает вызвано тем, что каждый имеет ввиду свой тип причинности.
Вот я, например, всё больше о материальных и формальных причинах люблю поговорить, а Вы - о целевых и движущих.