| ← | Дмитрий Шабанов | → |
| Живём длинно. Об украинском протесте, "коктейлях Молотова" и окситоциновой волне | Обсуждение перехода от гермафродитизма к раздельнополости как пример неклассического развития естественнонаучной гипотезы | О природе мужского и женского, или от конъюгации к оогамии |
| Колонка для Компьютерры #136 | Колонка для Компьютерры #137 | Колонка для Компьютерры #138 |
Я рад тому, как была принята позапрошлая колонка (не прошлая, политическая, а именно позапрошлая, посвящённая переходу от гермафродитизма к раздельнополости). Кажется, мне удалось убедить в её основной идее многих читателей. Впрочем, убедительность в популярном изложении для научной идеи — ещё не самое главное.
Чтобы гипотеза прошла нормальный цикл созревания и развития, необходимо следующее:
— вписать её в историю вопроса, проанализировать аргументацию, высказанную по поводу этой и аналогичных идей;
— тестировать гипотезу на совместимость с накопленным знанием и имеющимися фактами;
— выдвинуть основанные на гипотезе прогнозы, которые можно подтвердить или опровергнуть.
Это большая работа, и сам я с ней практически наверняка не справлюсь. Прохождение этих этапов может обеспечить взаимодействие между разными специалистами. Думая о том, как его организовать, я понял, что созревание этой идеи шло очевидно «неклассическим» образом. Ключевую роль в нем сыграло взаимодействие через Сеть людей, для которых изучение этой проблемы не является основной задачей. Поэтому в этой колонке я хочу подробно описать траекторию, которую эта идея прошла к настоящему моменту.
По своей узкой специальности я популяционный эколог бесхвостых амфибий. Моя диссертация была посвящена популяционному разнообразию жаб (я заинтересовался им, разбираясь в изменчивости состава жабьего яда). В 1999 году я познакомился на научной конференции с коллегами из санкт-петербургского Зоологического института РАН и присоединился с их подачи к изучению гибридизации у лягушек. Вместе мы открыли то, что потом назвали Северско-Донецким центром разнообразия гибридогенного комплекса зелёных лягушек.
Специфика ситуации, в которой оказались мы с моими харьковскими коллегами, состоит в том, что мы работаем едва ли не с самыми интересными популяционными системами лягушек в мире, но серьёзно уступаем и западным, и даже российским специалистам в доступе к современной технике и необходимым финансам. На Западе можно собрать партию материала, прогнать его через супер-пупер-современные технологии и получить интересный и публикабельный результат. Мы, чтобы удержаться на плаву, должны очень хорошо думать, осмысливая проблему, над которой работаем.
Увы, занятия наукой не являются для меня основными. Я преподаватель, а ещё и популяризатор (поэтому-то вы сейчас и читаете мой текст). Многие вещи приходится раз за разом пересказывать студентам. Кроме стандартных истин, прописанных в учебниках, со временем нарабатываешь свои способы объяснения. Для чтения лекций я стал собирать нарезку из популярных фильмов, скачанных через торрент-трекеры. В мой арсенал попал и фрагмент из фильма National Geographic, посвящённый сражениям гермафродитов (тот самый, на который я ссылался в прошлой колонке).
Благодаря синопсисам Александра Маркова на «Элементах», а также в результате работы обозревателем научных новостей для «Компьютерры» я узнал, какие споры связаны с проблемой происхождения раздельнополости. С моей точки зрения, ответ со всей очевидностью следовал из факта боев гермафродитов. Несколько лет я повторял его студентам, откладывая на будущее поиски источника, где излагается это объяснение. Преподаватели и иные профессиональные пересказыватели часто забывают источники своих идей, и иногда приходится себя перепроверять. Я исходил из того, что раз эта идея пришла в голову мне, то она должна была приходить и в чьи-то головы до меня.
В 2009 году благодаря очередной новости Маркова я осознал, что сидящее в моей голове объяснение как минимум не является широкоизвестным. Примерно тогда же я пришёл к пониманию, что эволюция гемиклонального наследования у гибридных лягушек является частью общего процесса эволюции рекомбинации (от горизонтального переноса до культурного наследования). Раз так, я решил, что разобраться в этой теме будет полезно для «лягушачьей» работы.
Я посоветовался с Марковым. Александр написал, что в явном виде эта идея ему незнакома, и посоветовал, в каком направлении стоит искать её аналоги. Я выступил с докладом на конференции и опубликовал статью в её материалах (выразив благодарность Александру за критику её чернового варианта).
Я решил: главное, что я могу сделать для развития представлений о переходе от гермафродитизма к раздельнополости, — моделировать этот переход с применением примерно того же инструментария, который мы использовали для изучения трансформаций популяционных систем лягушек. Попробовал. Сделал одну, другую, третью модель... Меня ожидало разочарование. Самцы не вытесняли гермафродитов, а устанавливали с ними динамичное равновесие, достигнув определённой устойчивой «концентрации». В этих условиях не получали ожидаемого преимущества и самки. Я оказался в тупике. Мне стало ясно, что надо выяснить, при каких условиях гермафродитам выгодно конкурировать за мужскую роль. Самому решить эту проблему не получалось: не с кем было её обсуждать. Я попытался сманить на изучение этой проблемы с помощью моделей вначале разумную студентку, а потом разумного студента. Увы, дело у них тоже не пошло.
Вывел меня из этого тупика опять же Марков. В своём ЖЖ-блоге он поместил следующую запись:
«Всё же сперматозоиды гораздо дешевле, чем яйцеклетки, и поэтому каждый гермафродит потенциально может оставить гораздо больше потомков, выступая в роли отца, чем в роли матери. Отсюда конкуренция за мужскую роль. Отсюда эволюционная нестабильность гермафродитизма у активных животных. Если появится мутация, перераспределяющая ресурсы организма в пользу повышения “мужской конкурентоспособности” и в ущерб женской функции, эта мутация с большой вероятностью будет поддержана отбором. Популяцию гермафродитов заполонят “социальные паразиты” — специализированные самцы-мутанты.
Оставшимся гермафродитам при этом станет уже незачем сохранять мужскую функцию. Им всё равно не победить новоявленных “специалистов” в конкуренции за мужскую роль. Соответственно, оставшимся гермафродитам станет выгодно специализироваться на женской функции.
Может быть, именно поэтому среди животных (особенно подвижных, быстрых, таких как хордовые, большинство членистоногих, головоногие...) всё-таки преобладает раздельнополость, а не гермафродитизм с перекрёстным оплодотворением. При всех его достоинствах как в смысле эффективности, так и в смысле гендерного равенства».
Эта идея была с энтузиазмом воспринята читателями блога Маркова. Я напомнил ему, что она вроде как моя. Он тут же согласился, сославшись на то, что не всегда помнит, откуда взялась та или иная мысль, сидящая в его голове. Не сомневаюсь, что так оно и есть: сам от этого страдаю.
Многозначительной для меня была реакция одного нашего с Марковым общего знакомого. В своём ЖЖ он восхитился процитированным фрагментом у Маркова. Я послал ему ссылку на свою статью. Он вспомнил, что даже её прочёл. Я ответил в духе «Вот какова роль авторитета: когда эту мысль высказывал я, Вы не поверили, а со слов Маркова сразу согласились». Мой корреспондент уверил меня, что дело не в недоверии ко мне, а в том, что моя статья требовала усилий для понимания ссылок на теорию игр, а у Маркова все оказалось изложено намного проще.
Проще! Но, по-моему, из-за этой простоты в его объяснении отсутствуют существенные аргументы в пользу обсуждаемой идеи. Кроме прочего, я бы делал акцент не на «дешевизне» сперматозоидов (о ней я ещё планирую поговорить подробнее), а на возможности самца участвовать во многих спариваниях (что, конечно, тесно связано с этой самой «дешевизной»). С другой стороны, может быть, Марков и прав, допуская не одномоментное отключение женской функции у гермафродитов, а постепенную сдвижку приоритетов в пользу мужской...
Реакция на пост Маркова заставила меня вернуться к этой идее. Я решил воспользоваться служебным положением. В двух лекционных курсах, которые я читал в осеннем семестре прошлого года (курсах батрахологии и истории биосистем), я выделил по три занятия на объяснение тех идей, которые хотел довести до ума. От проговаривания этой проблемы я сам, как мне кажется, продвинулся в её понимании. С декабря прошлого года я стал писать серию колонок, посвящённых этому вопросу (вот первая, а эта — уже седьмая). Существенную часть новогодних каникул пришлось потратить на моделирование, в котором я постарался отразить своё новое понимание. Кое-что получилось; что — сейчас расскажу.
Ещё до того, как я описал обсуждаемую здесь идею, два читателя изложили её в комментариях. Здесь я вижу и свою заслугу: я к ней качественно подвёл. Более того, ещё до того, как я описал свою модель, результаты своего моделирования прислал один из читателей «Компьютерры», Касум Ахмедли из Азербайджана!
Модель Касума построена неожиданным для меня образом. С помощью остроумных приёмов она создана на основе программы для моделирования химического равновесия. С некоторыми аспектами этой модели (и связанными с ними выводами Касума) я не согласен. Сейчас мы потихоньку продвигаемся в обсуждении и понимании наших разногласий. К моделированию перехода от раздельнополости к гермафродитизму выразил желание приобщиться ещё и один математик из Киева. Посмотрим, что получится. Мне очень интересно.
Итак, в этой колонке я хочу поделиться с читателями двумя удивлениями. Первое — как там оно получается с эволюцией размножения у гермафродитов. Второе — в какой степени взаимодействие компетентных людей, объединённых подходящими узлами в Сети, способствует развитию интересующих их идей. Если я прав, это какой-то новый способ «делать науку». Конечно, результатом такой работы должна быть научная статья в пристойном журнале, но путь к её написанию оказывается неклассическим. Я рад: такое взаимодействие помогает мне не вариться в собственном соку, а в ходе взаимодействия с другими людьми продвигаться в понимании интересной для меня проблемы.
Кому по-настоящему принадлежит обсуждаемая мной идея, сказать тяжело. Наверняка она приходила в разные головы и до меня. Явных предшественников я пока не нашёл. Мои рассуждения напоминают идею Г. А. Паркера и соавторов, опубликованную в 1972 году, — ту самую, которая касается «дешевизны» сперматозоидов. Если всё сложится, как я хочу, мы ещё обсудим её подробнее.
А теперь настало время рассказать о том, что получилось в ходе моего моделирования. Свою модель я подробно описал в сетевом конспекте курса моделирования в экологии с использованием Microsoft Excel. Там же приведена ссылка, по которой можно скачать саму модель.
Идея такая. Задаётся начальное распределение типов оплодотворённых яйцеклеток. С его учётом случайным образом формируется стадо производителей из 100 особей, которые могут принадлежать к одному из трёх полов: быть гермафродитами, самцами или самками. Из этого стада 100 раз случайно извлекаются пары особей, которые скрещиваются (или пытаются скреститься) друг с другом. Гермафродиты и самки, забеременев, выходят из игры; самцы могут совершать попытки спаривания неограниченное количество раз. По итогам состоявшихся спариваний определяется распределение типов оплодотворённых яйцеклеток, и цикл повторяется. По итогам двух сотен циклов строится график динамики трёх полов. Модель позволяет легко изменять состав потомков при разных типах скрещивания, а также учитывать разную выживаемость полов и их разную привлекательность друг для друга.
Я так подробно обсуждал историю идеи, что на пересказ результатов моделирования почти не остаётся места. Скажу о главных выводах. После экспериментов с моделью они представляются вполне тривиальными. Как ни странно, до моделирования я до таких выводов додуматься не смог; в этом эффекте для меня и проявляется один из смыслов мороки с моделями.
Вот, смотрите.
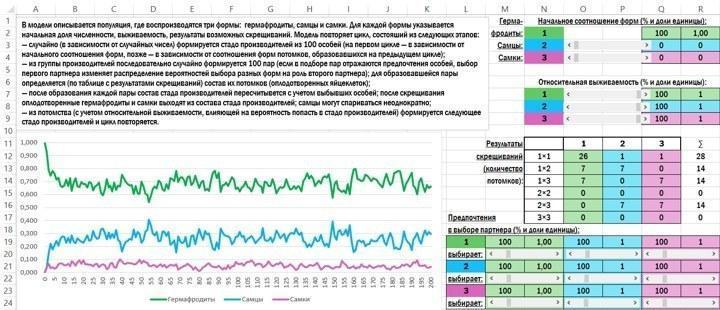
Обратите внимание на две вещи — на табличку, где задаются результаты скрещиваний, и на график, отражающий динамику трёх полов. При всех типах скрещивания распределение полов потомков соответствует распределению полов родителей, за тем исключением, что у гермафродитов понемногу появляются и самцы, и самки. Мы видим, что при этих условиях устанавливается равновесное соотношение полов, вокруг которого происходят случайные колебания.
Быть самцом в этой ситуации выгодно: самцы могут оставить больше потомков, чем гермафродиты. Но поскольку в потомстве от скрещивания самцов и гермафродитов половина потомков — гермафродиты, вытеснить их самцы не могут. Раз самцы не получают существенного перевеса, не возникает и ситуация, при которой получат преимущество самки.
Пробуем иначе. При скрещивании самцов и гермафродитов самцы должны иметь хотя бы небольшое преимущество (или в численности потомков, или в их выживаемости). При скрещивании самок и самцов не должны снова появляться гермафродиты; только в этом случае популяция может от них избавиться. От чего это зависит? От механизма наследования пола.
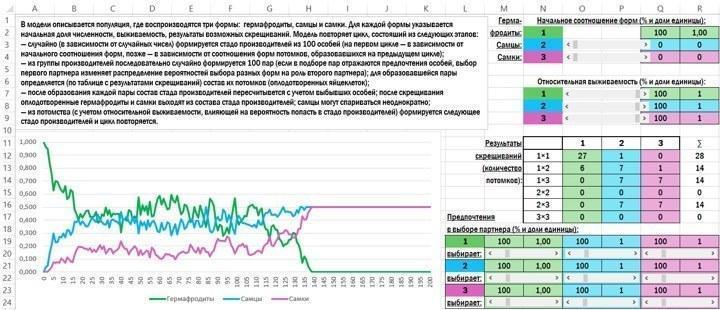
Так или иначе, при некоторых условиях (которые могут касаться не только состава потомков, но и различия в выживаемости и (или) взаимной привлекательности разных форм) переход от гермафродитизма к раздельнополости, которого мы ожидали, всё-таки происходит.
Что из этого следует? В иной раз...
| ← | Дмитрий Шабанов | → |
| Живём длинно. Об украинском протесте, "коктейлях Молотова" и окситоциновой волне | Обсуждение перехода от гермафродитизма к раздельнополости как пример неклассического развития естественнонаучной гипотезы | О природе мужского и женского, или от конъюгации к оогамии |
| Колонка для Компьютерры #136 | Колонка для Компьютерры #137 | Колонка для Компьютерры #138 |
Коментарі
Переход
Дмитрий, интересная гипотеза
Одним из факторов, который может привести к исчезновению гермафродитов во второй модели, мне кажется, может быть энергетическая невыгодность поддержания органов для женских и для мужских гамет, как показано на примере пещерных слепых рыб, выгода по энергетическим затратам на поддержание органа даже в несколько процентов может привести к его исчезновению
Да, конечно.
Если одна из половых систем станет ненужна, она со временем окажется нефункциональной. Все, что сохраняется, существует благодаря тому, что отбор поддерживает нормальное функционирование адаптивных структур. Исчезнет отбор - рассыпется и ненужная адаптация, а если она будет еще и "дорогой", то включится отбор, направленный на ее демонтаж.
Но я думаю, что могут быть еще более важные факторы изменений, которые и обсуждал в колонке, поэтому эту тему я в ней не затрагивал...