| ← | Дмитрий Шабанов | → |
| Трудности профориентации | Парадокс ускорения эволюции | Когда отбор становится неэффективным? |
| Колонка в КомпьютерреOnline #45 | Колонка в КомпьютерреOnline #46 | Колонка в КомпьютерреOnline #47 |
В позапрошлый раз я обосновывал, что в движении к III синтезу эволюционная биология должна преодолеть ограничения II синтеза - СТЭ, синтетической теории эволюции, сформировавшейся около семидесяти лет назад. Пора поговорить о недостатках СТЭ.
Для нас пока будет достаточно определения СТЭ, которое дал Александр Павлович Расницын. Согласно этой теории,
эволюционный процесс представляется как динамика аллельных частот в популяциях, контролируемая отбором (через оценку жизнеспособности и, соответственно, дифференциальное воспроизводство фенотипов) и стохастическими процессами".
Не вполне ясно? Я поясню, старательно изложив классические для СТЭ взгляды (даже те, с которыми не полностью согласен). Это проще сделать, комментируя рисунок.
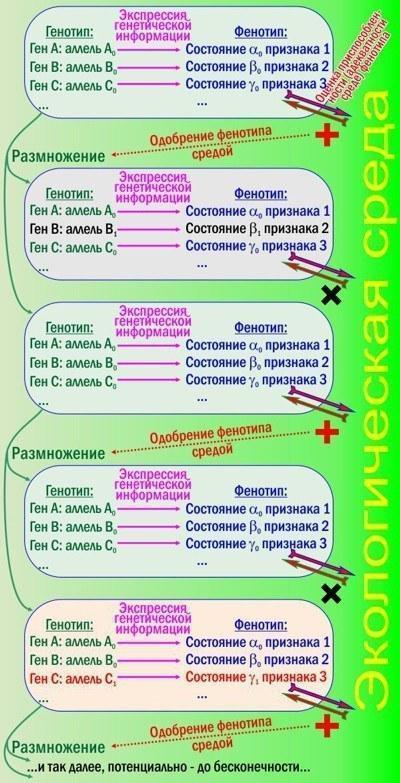
Рис. 1. Дарвиновская эволюция согласно СТЭ. Отбирается фенотип, воспроизводится генотип
Пять прямоугольников на рисунке - три поколения очень простых организмов: бактерий, к примеру. Терминология понятна? Комплекс наследственных задатков организма называется генотипом, а совокупность его проявленных признаков - фенотипом. Генотип - совокупность генов; гены могут находиться в разных состояниях - аллелях. В фенотипе можно выделить отдельные признаки, которые могут находиться в различных состояниях признака.
В ходе развития организма его фенотип меняется, причём его изменения зависят от генотипа. Организм взаимодействует со средой, строит себя за её счёт. Если фенотип адекватен тем возможностям, которые предоставляет среда, организм успешно развивается и оставляет потомство; неадекватные (неприспособленные) фенотипы уничтожаются или отстраняются от размножения.
Видите на рисунке петли обратной связи? Генотип определяет фенотип, среда оценивает фенотип и позволяет размножаться носителям оптимальных генотипов. При размножении возникают ошибки генотипа. Отбор на рисунке отсеял изменение гена В и поддержал изменение гена С. Коли так, мы увидели элементарное эволюционное событие: изменение аллельных частот в популяции.
Вы помните, что новая информация - это всегда следствие запоминания последствий случайного выбора? Дарвиновская эволюция - неизбежное следствие размножения, изменчивости, наследственности и отбора. Эта тетрада на рис. 1 налицо - значит, будет идти эволюция. Конечно, успех и неудача особей не всегда закономерны; отбор действует через изменение их вероятностей; поэтому-то в определении Расницына упомянуты стохастические процессы.
Общая картина ясна, проблема - в деталях. Чтобы отбор фенотипов эффективно менял генотипы, надо, чтобы фенотипы и генотипы были однозначно связаны друг с другом. Так бывает не всегда.
Увы, до сих пор не выработана общепринятая классификация признаков (и их возможных состояний) в зависимости от того, как управляется их развитие. Показанные на рис. 1 признаки управляются самым простым способом: каждый из них зависит от определённого гена. Так, возможно, что ген В определяет структуру фермента, позволяющего бактерии питаться каким-то веществом. Когда ген нормален (аллель В0), всё хорошо. Во втором поколении мутация В1нарушила работу фермента, нарушила усвоение пищи и привела организм к гибели.
Мутация С1 интереснее: она привела не к снижению, а к повышению жизнеспособности. Может быть, усилила связи между соседними цепочками аминокислотных остатков в каком-то белке, отчего этот белок стал устойчивее к повышенным температурам.
Равновероятны ли неблагоприятные и полезные мутации? Чем сложнее и совершеннее организм, тем менее вероятно, что случайное изменение пойдёт ему на пользу. Некоторые авторы даже считают вероятность полезного изменения астрономически малой. Пожалуй, это не так, и убедительные эксперименты (посвящённые как раз приспособлению бактерий к повышенной температуре) показывают, что полезные мутации - явление нечастое, но наблюдаемое.
Но любой ли признак зависит от одного гена? Нет. К примеру, в только что упомянутом эксперименте по эволюции термоустойчивости бактерий эффект многих мутаций невозможно было оценить по отдельности, без учета аллельного состояния других генов. Одна и та же мутация в комплекте с одними аллелями иных генов оказывается благоприятной, в сочетании с другими - вредной. Это значит, что определяющее судьбу состояние признака зависит от нескольких генов.
Снижают ли взаимодействия между генами эффективность отбора? Да, и особенно для видов с половым размножением. Я приведу пример именно для таких организмов, каждый из которых имеет по два одинаковых или разных аллеля каждого гена и передаёт потомкам лишь один из них.
Представим, что некий счастливый организм приобретает сочетание двух редких (возможно, недавно возникших) аллелей, каждый из которых поодиночке неблагоприятен: А1 и В1. В популяции распространены аллели А0 и В0. Сочетания А1 и В1 благоприятны, а вот А1 и В0, как и А0 и В1 - вредны (и то и другое - в сравнении с нормой, А0 и В0).
Давайте я не буду объяснять логику решения генетических задач, а просто распишу соотношения численностей групп потомков от скрещивания счастливого обладателя удачного сочетания двух мутаций с наиболее вероятным партнёром. Рассмотрим случаи, когда оба этих аллеля доминантны (влияют на фенотип даже у особей, у которых присутствуют в одной копии) и когда они рецессивны (проявляются в фенотипе только у организма, имеющего две их копии).
Если аллели А1 и В1 доминантны, только четверть потомков будет обладать улучшенным фенотипом (показаны полужирным шрифтом), а две четверти - ухудшенным (выделены подчёркиванием):
А1А0В1В0 × А0А0В0В0 → А1А0В1В0 : А1А0В0В0 : А0А0В1В0 : А0А0В0В0.
Если рассматриваемые аллели рецессивны, у всех потомков будет нормальный фенотип:
А1А1В1В1 × А0А0В0В0 → А1А0В1В0.
А что проявится в следующем поколении?
А1А0В1В0 × А1А0В1В0 → А1А1В1В1 : 2А0А1В1В1 : 2А1А1В1В0 : 4А1А0В1В0 : 2А1А0В0В0 : 2А0А0В1В0 : А0А0В1В1 : А1А1В0В0 : А0А0В0В0.
Благоприятный признак проявится лишь у одного потомка из шестнадцати, а шесть из шестнадцати потомков будут иметь ухудшенный фенотип! В этих (и иных, не рассмотренных нами) случаях подобного взаимодействия аллелей они чаще будут встречаться в популяции порознь, когда отбор будет работать против них, чем вместе, в одобряемом отбором состоянии. Ситуации, когда отбор будет поддерживать взаимодополняющую пару аллелей, окажутся редки по сравнению со случаями, когда он будет уничтожать эти аллели поодиночке.
Взаимодействие генов снижает эффективность отбора! И чем сложнее организмы, чем сложнее определяются их признаки, тем в меньшей мере направленный отбор фенотипов будет приводить к направленному отбору аллелей.
У по-настоящему сложных организмов между генотипом и фенотипом вклинивается ещё несколько механизмов регуляции, затрудняющих сортировку аллелей отбором. Один из них на языке СТЭ называется расширением нормы реакции. Организмы с одинаковым генотипом могут отличаться из-за развития в разных условиях (или просто в силу случайности). Представьте себе трёх человеческих близнецов-тройняшек с одинаковыми генотипами. Одного вырастило племя первобытных охотников, другой попал в интернат для спортсменов-силовиков, третий - в спецматшколу. Несмотря на идентичную наследственность, они вырастут очень разными - и физически, и по поведению. В отличие от ненаправленных мутаций, их отличия повысят приспособленность их носителей. При отборе (например, половом - покорении сердца потенциально плодовитой красавицы) эти три близнеца будут иметь разные шансы, ведь отбор (и половой в том числе) оценивает именно фенотипы!
Второй механизм связан с эпигенетической регуляцией генной активности, особенно характерной для млекопитающих. В зависимости от состояния организма, он передаёт потомкам геномы с разными химическими "метками" на генах (метилированием или иными изменениями "букв" генетического текста). Эти метки влияют на активность генов и, значит, на признаки. Обычно эпигенетические метки обеспечивают тот вариант развития (в пределах нормы реакции), который более соответствует данным условиям среды. При изменившихся условиях эти метки могут быть стёрты или переставлены.
Как эти изменения влияют на эффективность отбора? Новую схему со многими поколениями рисовать не буду, покажу лишь логику формирования признаков одной особи.
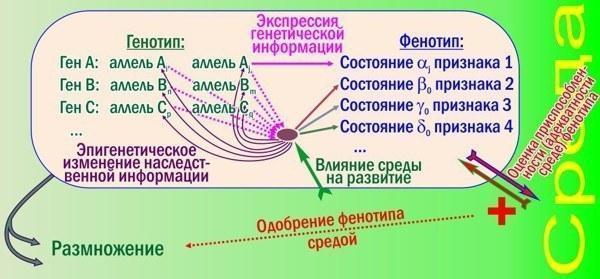
Рис. 2. Тут показаны лишь некоторые усложняющие работу отбора по сортировке аллелей механизмы, характерные для высокоразвитых видов
Кроме основной петли обратной связи (генотип влияет на фенотип; отбирая лучшие фенотипы, среда отбирает лучшие генотипы) появились новые уровни регуляции. Смотрите: развитие признака 1 по-прежнему управляется достаточно просто, он зависит от одного аллеля. Зато признаки 2, 3 и 4 зависят и от всех показанных генов, и от среды, причём в разной степени. Взаимодействие генных продуктов и средовых влияний формирует сложнейший узел взаимопереплетённых причин и следствий. От результатов такого взаимодействия зависит и сама генная активность: в зависимости от него гены подвергаются эпигенетическому редактированию - обратимому наследственному изменению.
Эти изменения снижают эффективность сортировки аллелей отбором. Связь фенотипов с генотипами становится очень неоднозначной (кроме "простых" признаков, как признак 1 на рис. 2). Механизм эволюции, эффективно работавший на рис. 1, будет пробуксовывать на рис. 2.
Эволюция - это соревнование в скорости выработки новых приспособлений. Вернитесь к первому рисунку: во втором показанном там поколении выигрывал нормальный генотип, а в третьем носитель недавней нормы уже проиграл, так как появились более приспособленные организмы. Как же могли возникнуть организмы, у которых эффективность отбора (и эффективность выработки новых приспособлений) понизилась, - они ведь неминуемо должны были проигрывать тем, кто эволюционировал быстрее?
Дело в том, что у сложных организмов эффективность отбора не снижается, а вырастает. Опять процитирую Расницына:
...возраст (в миллионах лет) полусовременной фауны (в которой половина видов - современные, половина - вымершие) у крупных млекопитающих (хоботные и копытные) - 0,2, у мелких - 0,5, у птиц и рыб - 0,7, у насекомых - 3-7, у моллюсков - 3,5-5, у диатомовых водорослей - 15. Время полувымирания (по аналогии с полураспадом, время вымирания половины исходного числа видов) в тех же единицах наименьшее у слонов - 0,18, а в среднем у млекопитающих - 0,54, у костистых рыб - 3,5, у граптолитов - 1,3, иглокожих - 4,2, двустворок - 7, у планктонных фораминифер - 5, бентосных - 18-24, у диатомовых водорослей - 5,5, у динофлагеллат - 9. На старших таксономических уровнях различия подобные, но часто более резкие. Например, возраст полусовременной фауны для родов млекопитающих - 4 млн лет, для птиц - 10, рептилий - 20, рыб - 30-50, насекомых - 40, моллюсков - 60, фораминифер - 230"
Кроме изложенных причин, есть ещё целый ряд наглядных факторов, которые должны были бы сделать эволюцию крупных животных очень медленной (о них подробно пишет Расницын). К примеру, у каждой особи хоботных бывает лишь немного потомков, а продолжительность поколений катастрофически велика. За то время, пока растёт один слонёнок, сменятся десятки (если не сотни) поколений насекомых, диатомей или фораминифер с бесчисленным количеством особей. У хоботных просто не будет достаточного материала для отбора, они должны эволюционировать крайне медленно...

Рис. 3. За несколько десятков миллионов лет истории хоботных возникло более сорока их родов и более трёхсот видов!
По скорости их эволюции (измеренной и в скорости образования новых видов, и в скорости изменения признаков) хоботные - одни из рекордсменов в биосфере. С точки зрения СТЭ этот феномен необъясним. Тут нужна идея эволюции эволюции - совершенствования по мере развития живого самих механизмов выработки приспособлений. Приведу лишь один пример. Семейство Люди резко повысило скорость выработки приспособлений, когда усовершенствовало механизмы культурного наследования и добавило его к общему, генетическому наследованию. Вероятно, это усовершенствование эволюционного механизма было не первым в нашей истории.
Слоны, как и другие млекопитающие, эволюционируют и с использованием механизмов, описанных СТЭ, и как-то иначе. Дополнительные петли обратной связи в реализации их генетической информации не снижают, а повышают эффективность их эволюции. Как - мы знаем ещё не до конца. Что же, нашему знанию есть куда эволюционировать!
| ← | Дмитрий Шабанов | → |
| Трудности профориентации | Парадокс ускорения эволюции | Когда отбор становится неэффективным? |
| Колонка в КомпьютерреOnline #45 | Колонка в КомпьютерреOnline #46 | Колонка в КомпьютерреOnline #47 |
Коментарі
Пути эволюции бывают заковыристы
Как же могли возникнуть организмы, у которых эффективность отбора (и эффективность выработки новых приспособлений) понизилась...
А почему нет? В принципе, это возможно. Например, при длительных более-менее стабильных условиях отбор может предпочесть другие качества, а эффективность выработки новых приспособлений уйдёт на второй план. Да, в результате вид окажется втянутым в эдакую генетически-фенотипическую яму, из которой шансов выбраться может оказаться маловато. Да, в случае серьёзных изменений условий это может быть приговором таким видам. И такой вид будет кандидатом на вымирание. Сколько хоботных вымерло? ;)
Можно в пример привести и симбионтов, которые нередко теряют самостоятельность, теряют кучу генов, именно потому что в силу изменения условий обитания (которые становятся более стабильными) особой необходимости в этом нет.
Это я к тому, что одно другого не исключает.
Насчет заковыристости согласен
Мне стоило подчеркнуть, что я говорю о "заторможенной" группе, а не о виде. Да, вид может войти в состояние живого ископаемого и стать из-за этого по-настоящему хрупким. А достаточно большая группа, состоящая из многих видов (которые должны же были возникать) - нет.
А хоботные, конечно, вымерли все, кроме двух (или трех в зависимости от статуса лесных африканских слонов). Но ведь их многочисленные виды, перед тем, как вымереть, должны были возникнуть!
Я же не против эпигенетики. Я - за. ;)
Но вопрос, конечно, интересный.
Цугцванг?
Возможно, что в этой самой группе и был "открыт" один из высоко-адаптационных механизмов. Который мог оказаться настолько чувствительным что чрезмерно приспособил конкретные виды этой группы к конкретным условиям. Но при быстрых изменениях условий (по причинам, которые Вы статье уже изложили) обратно приспособиться многие из них уже не смогли/не успели.
Кстати, такое вымирание вполне соответствует рис.2, если оценивать его исходя из теории систем автоматического управления. Если система сконструирована (в данном случае эволюционировала) из расчёта на определённые параметры, находящиеся в определённых диапазонах и в определённом сочетании/зависимости друг от друга, то любые отклонения от расчётных могут завести систему в непредвиденное состояние, или попросту говоря, сломать её.
Видимо, не все учли
Наверное, скорость выработки новых приспособлений слабо зависит от продолжительности поколений. Новые приоспособления завоевывают место очень быстро, в течение десятков - сотен поколений. Далее достигается относительный оптимум, равновесие, которое нарушается только изменениями в среде. Другое дело, что разные группы организмов, эволюционируя, по-разному влияют на эту среду. Ведь взаимодействие организмов со средой обоюдные, возникают вторичные, третичные и т.д. обратные связи. Тогда не удивительно, что более крупные организмы сильнее влияют на среду, изменяют ее, среда снова влияет на них, они снова на среду и т.д. Эта цепочка долго не утихает. Если бы можно было измерить степень такой индуктивности организмов, думаю, человек бы был чемпионом. По скорости изменения окружающей среды ему нет равных.
Комментарий к своему комментарию
Видимо, неправильно написал. Такая индуктивность зависит не от организма и его генотипа, а от случая, который сводит этот генотип с различными окружающими условиями. Иногода возникает своего рода резонанс, который и приводит к ускорению эволюции.
Уточнение
С тем, что Вы пишете, согласен. Но уточню свою мысль.
С точки зрения СТЭ продолжительность смены поколений не важна. Важно количество поколений. А у животных наподобие слонов в один миллион лет уложится намного меньшее количество поколений, чем у насекомых.
К уточнению
Может, и количество поколений не всегда важно? Десять - сто поколений у разных видов по времени отличаются на тысячелетия, затем, видимо, наступает равновесие, но это ведь 1000 лет - ерунда по геологическим меркам.
Но так или иначе,
с точки зрения СТЭ, увеличение срока смены поколений должно замедлять эволюцию, а именно те животные, у которых он наиболее велик (не только хоботные, но и киты, и даже люди) эволюционируют быстрее большинства прочих.
Я об этом и пишу
Я об этом и пишу. Видимо, фактор срока смены поколений менее важен. А вот то, что они резонируют со средой, влияет сильнее. Причем больше резонируют те виды, у которых каждая особь больше со средой взаимодействует, т.е. с более длинными поколениями.
Тут надо разобраться с тем, что означает "резонируют"
Может быть, Вы пишете примерно о том, о чем Марков писал вот тут: http://macroevolution.livejournal.com/61022.html#cutid1
Другое дело (можете найти мой комментарий, хотя бы по тому маркеру, что я упоминаю там Красилова), что я с ним не вполне согласен.
Сложное поведение и забота о потомстве повышают селективность смертности (ее зависимость от фенотипа), а не эффективность отбора (его влияние на генотип). Да, надо бы написать об этом подробнее...
Марков писал о К-стратегии
В статье Маркова обращается внимание на то что, при К-стратегии (которой, упощенно говоря, придерживаются животные с большой продолжительностью жизни и разнообразным поведением) вкладывается много ресурсов в защиту от хищников, вынашивание, кормление, постройку гнезд и т.д. Но упускается, что много ресурсов тратится на конкурентную борьбу и половой отбор. Ведь такие животные в конкуренции могут гораздо активнее использовать ресурсы, тем самым сильнее влияя на среду. И половой отбор (причина взрывной эволюции) у них должен быть гораздо более изощренным, разнообразным, и чаще возникать у разных видов в разных проявлениях.