Усова Е. Е. Возраст и скорость роста зеленых лягушек (Pelophylax esculentus complex) Нижнего Добрицкого пруда (Змиевской район Харьковской области) // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2014 – Вип.20, №1100. С. 204–212.
УДК: (57.022+57.032):591.84:591.158.1:597.851
Возраст и скорость роста зеленых лягушек (Pelophylax esculentus complex) Нижнего Добрицкого пруда (Змиевский район Харьковской области)
Е.Е.Усова
Харьковский национальный университет имени В.Н.Каразина (Харьков, Украина)
e.e.usova@gmail.com
С помощью скелетохронологии изучено 104 представителя Pelophylax esculentus complex (18 P. ridibundus, 66 диплоидных P. esculentus и 20 триплоидных P. esculentus) из гемиклональной популяционной системы Нижнего Добрицкого пруда (N 49°33'22", E 36°18'39"; Змиевский район Харьковской области, пойма реки Гомольша), расположенной на территории Национального природного парка «Гомольшанские леса». По срезам 4-й фаланги длиннейшего пальца задней конечности установлен возраст изученных особей, рассчитаны их годовые приросты. Максимальная скорость роста и для самцов, и для самок наблюдается перед 4-й зимовкой, что соответствует достижению половой зрелости в возрасте 5 лет. Различия в скорости роста P. ridibundus, ди- и триплоидов P. esculentus статистически незначимы.
Ключевые слова: Pelophylax esculentus complex, Pelophylax ridibundus, рост, скелетохронология.
Вік і швидкість росту зелених жаб (Pelophylax esculentus complex) Нижнього Добрицького ставу (Зміївський район Харківської області)
О.Є.Усова
За допомогою скелетохронологіі вивчено 104 представника Pelophylax esculentus complex (18 P. ridibundus, 66 диплоїдних P. esculentus і 20 триплоїдних P. esculentus) з геміклональної популяційної системи Нижнього Добрицького ставу (N 49°33'22", E 36°18'39"; Зміївський район Харківської області, заплава річки Гомільша), розташованої на території Національного природного парку «Гомільшанські ліси». По зрізах 4-ої фаланги найдовшого пальця задньої кінцівки встановлено вік вивчених особин, розраховані їх річні прирости. Максимальна швидкість росту і для самців, і для самок спостерігається перед 4-й зимівлею, що відповідає досягненню статевої зрілості у віці 5 років. Відмінності в швидкості росту P. ridibundus, ди- і триплоїдів P. esculentus статистично незначущі.
Ключові слова: Pelophylax esculentus complex, Pelophylax ridibundus, ріст, скелетохронологія.
Age and growth rate of green frogs (Pelophylax esculentus complex) of the Lower Dobritsky pond (Zmiyiv district, Kharkiv region)
E.E.Usova
With the use of skeletochronology 104 specimens from the population of Pelophylax esculentus complex (18 P. ridibundus, 66 diploid P. esculentus, and 20 triploid P. esculentus) inhabited the Lower Dobritsky pond (N 49°33'22 ", E 36°18'39"; Zmiyiv district of the Kharkiv region, the floodplain of the Gomolsha River at the National Natural Park "Gomilshanski lisy") were studied. The age of the individuals was estimated histologically by the sections of the longest toe’s 4th phalanx and determination of annual growth marks. The annual growth rates were calculated. The maximum growth rate was observed between the 4th and 5th hibernations for both males and females. It has been shown that differences in the growth rates between P. ridibundus, di- and triploid P. esculentus are statistically insignificant.
Key words: Pelophylax esculentus complex, Pelophylax ridibundus, growth, skeletochronology.
Введение
Скелетохронологическое изучение возраста и скорости роста является достаточно известным, но все равно недооцененным методом исследований пойкилотермных позвоночных. С помощью этого метода можно получить эмпирические данные, которые не только характеризуют возраст и динамику роста исследуемых животных, но и дают возможность оценить другие важные с точки зрения популяционной биологии параметры, как, например, относительную выживаемость различных совместно обитающих форм. Однако доказательное проведение подобных исследований требует их тщательной организации, максимально возможной унификации методов обработки первичных данных. В данной работе мы детально рассматриваем обработку скелетохронологических данных, характеризующих зеленых лягушек (представителей Pelophylax esculentus complex) из популяционной системы, представляющей особый интерес для исследований.
В гибридогенный комплекс зеленых лягушек, Pelophylax esculentus complex (=Rana esculenta complex), входят два родительских вида, Pelophylax lessonae (Camerano, 1882) и Pelophylax ridibundus (Pallas, 1771), а также их различные ди , три и тетраплоидные гибриды (Plötner, 2005), для которых мы используем (Шабанов та ін., 2009) имя, аналогичное видовому, Pelophylax esculentus (Linnaeus, 1758). Совместное обитание гибридов с родительскими видами поддерживается благодаря гемиклональному наследованию (передаче гибридами определенных геномов клонально, без рекомбинации), что представляет значительный теоретический интерес. Совместно обитающие представители родительских видов и их различных гибридов, объединенные совместным размножением и передачей из поколения в поколение как клональных, так и рекомбинантных геномов, получили название ГПС, гемиклональных популяционных систем (Шабанов та ін., 2009; Шабанов, Литвинчук, 2010). Одни из наиболее сложных известных современной науке гемиклональных популяций зеленых лягушек зарегистрированы в бассейне верхнего течения р. Северский Донец в регионе, который получил название Северско-Донецкого центра разнообразия зеленых лягушек (Шабанов та ін., 2009; Шабанов, Литвинчук, 2010). Наивысшее разнообразие различных форм зеленых лягушек зарегистрировано на территории Национального природного парка «Гомольшанские леса», расположенного в Змиевском районе Харьковской области (Коршунов, 2010). Наиболее интересной из изученных ГПС Pelophylax esculentus complex является та, которая населяет Нижний Добрицкий пруд (49°33'22" северной широты, 36°18'39" восточной долготы), расположенный в Добрицком яру перед его выходом в пойму реки Гомольша (правого притока р. Северский Донец). В составе этой ГПС велика доля триплоидных гибридов (Коршунов, 2010), в ее окрестностях были зарегистрированы даже единичные тетраплоиды (Borkin et al., 2004).
Материал и методы
Состав изученной нами выборки охарактеризован в табл. 1. Большинство изученных особей (75 экземпляров) составляют одну выборку, близкую к случайной, собранную 10 июля 2013 года. Ко времени сбора этой выборки нерест лягушек был закончен. Лягушек ловили, обходя пруд по периметру в первой половине ночи с яркими ручными фонарями, собирая при этом всех замеченных особей, находящихся на берегу или в воде на небольшом (до 1–2 м) расстоянии от берега. Ослепляя лягушек фонарем, их хватали руками. При использованном способе ловли удавалось собрать около 90% замеченных лягушек. Можно предположить, что как таксономический (P. ridibundus или P. esculentus), так и половой (самки или самцы) и размерный (крупные или мелкие) состав особей, сбежавших от ловцов, приблизительно соответствовал составу собранной выборки.
Таблица 1. Изученный материал
|
Форма |
Выборка, год сбора |
Ювенильных, экз. |
Самок, экз. |
Самцов, экз. |
Всего, экз. |
|
P. ridibundus |
2004–2009 |
3 |
5 |
1 |
9 |
|
2013 |
- |
8 |
1 |
9 |
|
|
P. esculentus, 2n |
2004–2009 |
10 |
4 |
5 |
19 |
|
2013 |
- |
10 |
37 |
47 |
|
|
P. esculentus, 3n |
2004–2009 |
1 |
- |
- |
1 |
|
2013 |
- |
11 |
8 |
19 |
|
|
Всего |
14 |
38 |
52 |
104 |
|
Обработка собранных особей включала следующие этапы:
– определение таксономического положения (как P. ridibundus или P. esculentus) по внешним признакам (Коршунов, 2010), прежде всего – размеру и форме внутреннего пяточного бугра, относительной длине голени и бедра, характеру окраски бедер и всего тела;
– определение пола по внешним признакам (наличие брачных мозолей и резонаторов у самцов);
– измерение длины тела при помощи штангенциркуля;
– фотографирование спинной стороны тела каждой особи;
– удаление четырех фаланг длиннейшего пальца одной из задних конечностей (правой);
– выдавливание капли крови на предметное стекло и приготовление стандартного мазка путем размазывания капли по поверхности стекла краем другого стекла;
– прижигание раневой поверхности для остановки кровотечения.
После выполнения описанных действий изученных лягушек выпустили обратно в месте вылова. В результате этого часть особей данной ГПС оказалась помечена, что может быть (в случае повторных ловов) использовано для оценки ее численности. Удаление пальца ноги выполняет роль групповой метки, для индивидуального опознания изученных особей в случае их повторного вылова в будущем планируется использовать фотографии узора их спинной стороны тела.
В сборе выборки, обработке пойманных лягушек и изучении мазков крови принимали участие студенты II курса биологического факультета Харьковского национального университета им. В.Н.Каразина, проходившие учебно-полевую практику по зоологии позвоночных на биологической станции университета в окрестностях с. Гайдары под руководством доцента Д.А.Шабанова.
Обработка мазков по ранее описанной методике (Бондарева и др., 2012) велась ими под руководством Е.В.Мелешко. Мазки высушивали на воздухе, фотографировали под микроскопом, определяли средний размер эритроцитов и на основании этого размера устанавливали плоидность изученных особей.
Таким образом, для диплоидных особей, входивших в выборку, собранную в 2013 г., их определение как P. ridibundus или P. esculentus основывалось на определении по внешним признакам, а идентификация триплоидных P. esculentus (внешне трудно отличимых от диплоидных P. esculentus, а иногда и P. ridibundus) основывалась на результатах определения размеров их эритроцитов.
Кроме выборки 2013 г., нами были изучены коллекционные экземпляры лягушек, собранные на Нижнем Добрицком пруду в 2004–2009 гг. Генотип этих особей был определен С.Н.Литвинчуком и Ю.М.Розановым в Институте цитологии РАН (г. Санкт-Петербург) с помощью проточной ДНК-цитометрии (Borkin et al., 2004).
В ходе скелетохронологического исследования четвертую фалангу длиннейшего пальца задней ноги лягушек очищали от мягких тканей, декальцинировали, получали на замораживающем микротоме срезы толщиной 20–22 мкм средней части диафиза, окрашивали гематоксилином Эрлиха, заливали глицерином, накрывали покровным стеклом, фотографировали временные препараты, а затем проводили измерения и подсчеты линий склеивания по фотографиям с использованием программы Adobe Photoshop CS5. Результаты измерений переводили в микрометры с использованием коэффициента пересчета, определенного с помощью фотографии объект-микрометра, сделанной при тех же условиях, что и фотографии препаратов.
Результаты измерений заносили в электронную базу данных, сделанную в Microsoft Office Excel, 2010. Методика интерпретации и обработки полученных результатов, позволяющая ретроспективно вычислить прирост исследованных лягушек в разные годы их жизни, подробно описана ниже.
Совокупность данных о зарегистрированных приростах лягушек переносили в программу Statistica 8, StatSoft Inc. и обрабатывали с помощью дисперсионного анализа, рассматривая как зависимую величину прирост лягушек в определенные годы их жизни, а как группирующие факторы – возраст лягушек, их генотип, пол и календарный год, в который был зарегистрирован наблюдаемый прирост.
Интерпретация фотографий срезов фаланг пальцев
Метод скелетохронологии основан на том факте, что по мере роста особи происходит и увеличение поперечного размера ее трубчатых костей (Смирина, 1983; Roitberg, Smirina, 2006 и другие работы). Во время остановок роста, соответствующих зимовкам (и иногда – летних похолоданий, что значительно затрудняет интерпретацию препаратов) в кости образуются линии склеивания, хорошо заметные на окрашенных срезах (рис. 1).

Рис. 1. Микрофотография среза фаланги пальца. Обозначения: I – эндостальная часть кости; II – периостальная часть кости; A, B, C – линии склеивания в эндостальной части кости; г – граница между эндостом и периостом; к.с. – кровеносный сосуд; 1–10 – линии склеивания в периостальной части кости
Существенная сложность при трактовке препаратов заключается в том, что по мере роста наружной части кости (периоста) в ее середине разрастается эндостальная полость, уничтожающая некоторые из линий склеивания периоста. На определенном этапе разрастание эндостальной полости останавливается, и она заполняется эндостальной костной тканью, растущей в направлении середины полости.
Результат определения возраста особи по скелетохронологическим данным зависит от количества зарегистрированных линий склеивания и от предположения о количестве линий, резорбированных по мере разрастания эндоста. Ошибки в подсчете линий (например, связанные с тем, что след от краткосрочной остановки роста принят за результат зимовки), или в оценке количества резорбированных при разрастании эндоста линий склеивания приводят к ошибкам в определении возраста. По мнению авторов, более сложной задачей является именно определение количества резорбированных линий; решая ее, необходимо определить величину, не отраженную на препарате явно. Интерпретация линий склеивания в периосте в этом отношении проще, так как эти линии видны и на фотографиях, и на препаратах. В том случае, если по одной фотографии принять обоснованное решение затруднительно, помочь может получение дополнительных фотографий, сделанных при иных настройках микроскопа (в частности, при фокусировке на ином уровне анализируемого среза), а также фотографирование иных срезов той же кости.
При определении вероятного количества резорбированных линий следует принимать во внимание:
– количество видимых остатков резорбированных линий;
– размер эндоста;
– размеры сохранившихся линий склеивания и их соответствие размерам, характерным для линий склеивания, образующимся у представителей данной популяции в разном возрасте. Два последних показателя для изученных нами особей отражены на рис. 2.
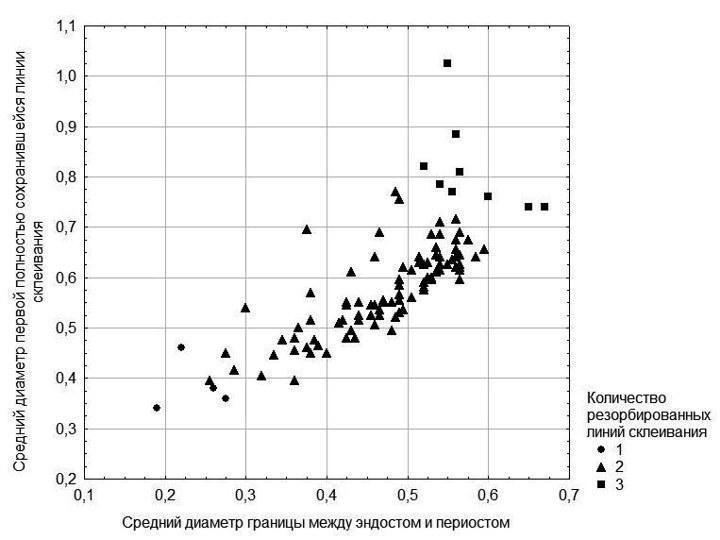
Рис. 2. Изменчивость параметров, на основании значений которых принималось то или иное предположение о количестве резорбированных линий у каждой особи
Длина тела лягушки во время зимовок вычисляется на основании предположения о том, что увеличение поперечного размера кости пропорционально увеличению длины тела (Roitberg, Smirina, 2006; Усова, Шабанов, 2009). Таким образом, мы принимаем, что lastlmed/L=ilmed/iL, где lastlmed – усредненный поперечник кости в момент исследования, L – длина тела в момент исследования, ilmed – установленный по размерам линии склеивания поперечник кости во время одной из зимовок, iL – длина тела, которую особь имела во время соответствующей зимовки. В таком случае, можно установить, что iL=L*.ilmed/lastlmed.
В том случае, если известны размеры двух последовательных линий склеивания в кости (и соответствующие им вычисленные размеры лягушки в целом), можно вычислить, насколько ее размеры увеличились за год, прошедший между этими зимовками. Кроме этого, можно определить размеры и динамику роста лягушек в календарные годы (в нашем случае – 2012, 2011, 2010 и т.д.).
Результаты и обсуждение
Общий результат скелетохронологического исследования показан на рис. 3.
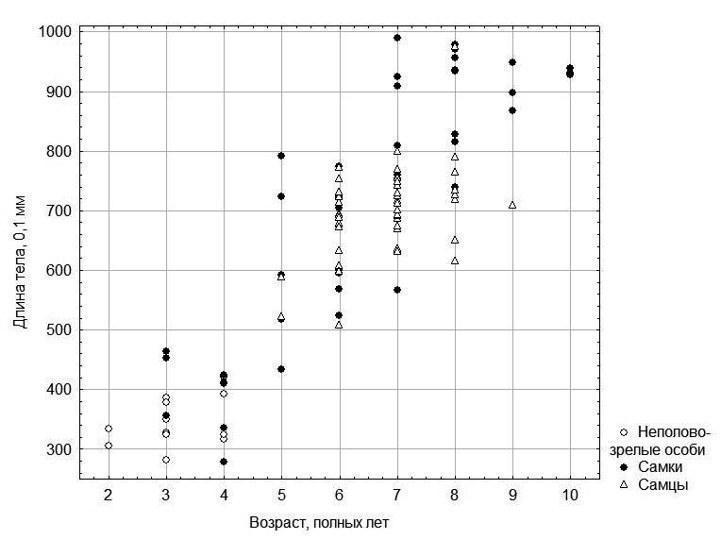
Рис. 3. Зависимость размера лягушек от их возраста
Величину, на которую увеличился размер лягушки за год, можно сопоставить с ее возрастом в этот год, общей продолжительностью жизни, календарным годом, полом, генотипом и другими параметрами. Совокупность данных о таких ежегодных приростах можно обрабатывать с использованием дисперсионного анализа. Так, на рис. 4 показаны результаты двухфакторного дисперсионного анализа, в котором величина ежегодных приростов рассматривалась как зависимый признак, а возраст и пол лягушек – как влияющие на него факторы. Как видно, динамика роста самцов и самок (объединенных в одну группу с неполовозрелыми особями) оказывается достаточно похожей. Максимальная скорость роста достигается перед четвертой зимовкой. Следует предположить, что эта зависимость свидетельствует о наступлении половой зрелости между четвертой и пятой зимовками. В анализе, отраженном на рис. 4, зависимость прироста от возраста, как и зависимость прироста от пола оказывается значимой; «взаимодействие факторов» (отличия в том, как приросты меняются с возрастом у представителей разных полов) статистически незначимо.
На рис. 5 показаны результаты двухфакторного дисперсионного анализа, в котором возраст и генотип рассматривались как основные факторы, влияющие на прирост лягушек. Влияние возраста, как и в предыдущем случае, осталось значимым, а влияние генотипа, как это ни удивительно, оказалось незначимо. На рис. 4 можно увидеть, что кривые, соответствующие разным генотипам, идут практически параллельно, и различие между ними можно рассматривать как результат статистического «шума».
Таким образом, данные скелетохронологического изучения всех форм зеленых лягушек, распространенных в Северско-Донецком центре их разнообразия (а именно P. ridibundus, а также ди- и триплоидов P. esculentus) можно рассматривать совместно.
Использованный в данной работе подход позволяет оценить также то, какие годы оказывались более или менее благоприятными для роста лягушек. Конечно, такое сравнение надо проводить по приростам в одном и том же возрасте. Если этого не делать, то замедление скорости роста лягушек с возрастом (отчетливо видное на рис. 4) создаст иллюзию, что каждый следующий календарный год оказывается хуже предыдущего. Для такого анализа мы использовали данные о приростах на 4-м, 5-м и 6-м году жизни, так как именно по этим годам мы имеем наиболее многочисленные данные. Результаты такого анализа показаны на рис. 6.
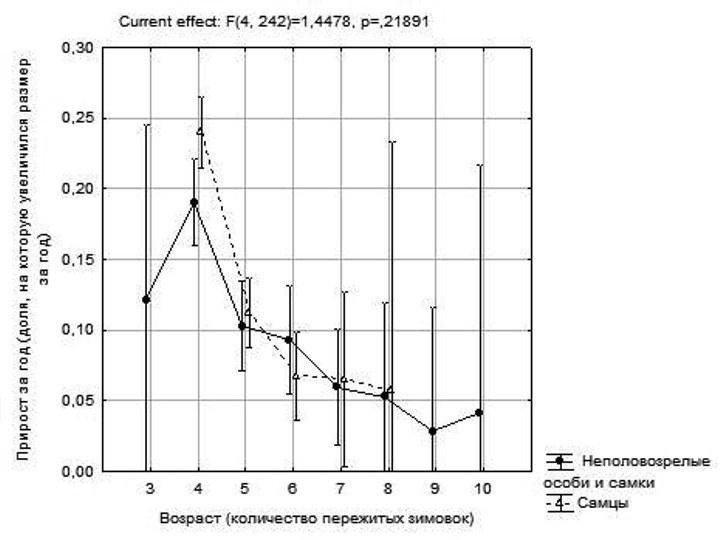
Рис. 4. Зависимость величины ежегодных приростов от возраста и пола лягушек
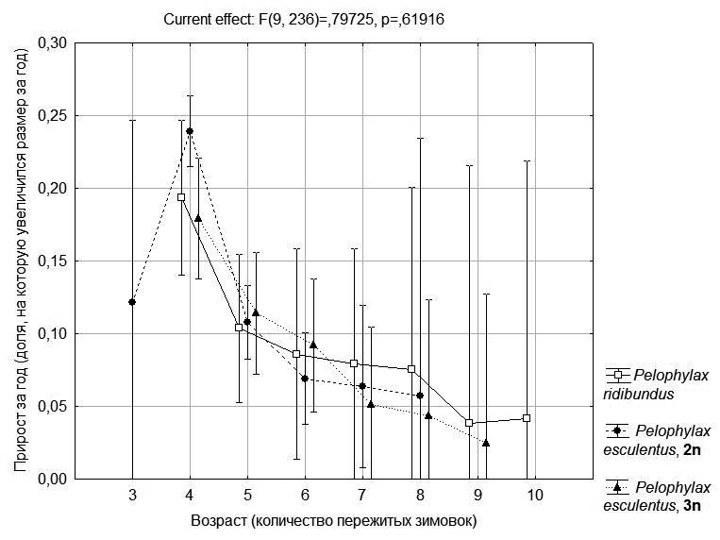
Рис. 5. Зависимость величины ежегодных приростов от возраста и генотипа лягушек
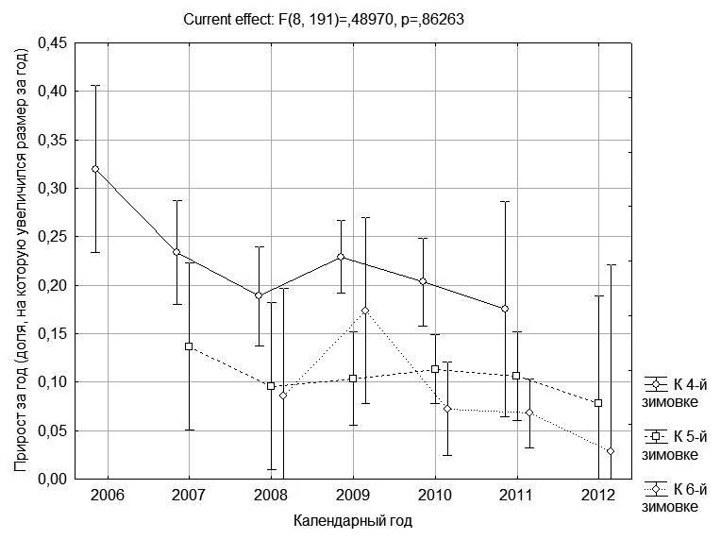
Рис. 6. Зависимость величины ежегодных приростов от календарного года
Сравнение годовых приростов за разные календарные годы показало незначимые отличия, однако определенная тенденция на рис. 6 все-таки заметна. Из последних лет наиболее благоприятным для лягушек оказался 2009 год, а после него регистрировалось определенная тенденция к снижению ежегодных приростов. Вероятно, аналогичное сравнение, проведенное на более обширном материале, позволит получить более надежные результаты.
Благодарности
Автор статьи выражает глубокую благодарность Д.А.Шабанову за научное руководство данной работой и помощь в интерпретации ее результатов. Кроме того, автор благодарит А.В.Коршунова и О.В.Михайлову за помощь в получении выборки 2013 года. В ее сборе и обработке принимали участие студенты-второкурсники, проходившие учебно-полевую практику по зоологии позвоночных: М.С.Альховская, М.Ю.Кит, А.О.Михайленко, А.А.Клыса, Д.В.Круговой, Ю.И.Костюк и А.О.Михайличенко. Особо ценный вклад в обработку этой выборки внесла Е.В.Мелешко. Для интерпретации результатов изучения сборов 2004–2009 годов чрезвычайно важны были результаты проточной ДНК-цитометрии, любезно предоставленные С.Н.Литвинчуком и Ю.М.Розановым.
В отработке описанной в данной статье методики автор получила неоценимую помощь от Э.М.Смириной, пионера скелетохронологических исследований в герпетологии.
Список литературы
Бондарева А.А., Бибик Ю.С., Самило С.М., Шабанов Д.А. Цитогенетические особенности эритроцитов зеленых лягушек из Северско-Донецкого центра разнообразия Pelophylax esculentus complex // Вісник Харківського національного університету імені В.Н.Каразіна. Серія «Біологія». – 2012. – Вип.15 (№1008). – С. 116–123.
Коршунов А.В. Экологические закономерности распределения Pelophylax esculentus complex в биотопах бассейна верхнего течения реки Северский Донец. Автореф. дисс. … канд. биол. наук / 03.00.16 – экология. – Днепропетровск, 2010.
Смирина Э.М. Прижизненное определение возраста и ретроспективная оценка размеров тела серой жабы (Bufo bufo) // Зоол. журн. – 1983. – Т.63, №3. – С. 437–444.
Усова Е.Е., Шабанов Д.А. Об оптимизации методики ретроспективной оценки динамики размеров тела представителей Pelophylax esculentus complex (Amphibia, Ranidae) при помощи скелетохронологии // Zoocenosis-2009. Біорізноманіття та роль тварин в екосистемах. – Дніпропетровськ, ДНУ, 2009. – С. 278–280.
Шабанов Д.А., Коршунов О.В., Кравченко М.О. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми // Біологія та валеологія. – Харків: ХДПУ, 2009. – Вип.11. – С. 116–125.
Шабанов Д.А., Литвинчук С.Н. Зеленые лягушки: жизнь без правил или особый способ эволюции? // Природа. – 2010. – №3. – С. 29–36.
Borkin L.J., Korshunov A.V., Lada G.A. et al. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian Journal of Herpetology. – 2004. – Vol.11, No 3. – P. 194–213.
Plötner J. Die westpaläarktichen Wasserfrösche. – Bielefeld: Laurenti-Verlag, 2005. – 161s.
Roitberg E.S., Smirina E.M. Age, body size and growth of Lacerta agilis boemica and L. strigata: a comparative study of two closely related lizard species based on skeletochronology // Herpetological Journal. – 2006. – Vol.16. – P. 133–148.