| ← | Дмитрий Шабанов | → |
| От хаоса наблюдений к динамической типологии: обсуждение на примере популяционных систем зелёных лягушек | О чем можно судить, измеряя флуктуирующую асимметрию? | Как будут изучать биологию и другие мировоззренчески значимые науки в такой школе будущего, о которой стоит мечтать? |
| Колонка для Компьютерры #113 | Колонка для Компьютерры #114 | Колонка для Компьютерры #115 |
В последнее время я выпал из привычного режима сочинения колонок. Защита Марины, TEDxKyiv и… практика. Месяц на биостанции, на берегу Северского Донца. Занятия со студентами и борьба с непрерывным желанием лечь поспать; на всё остальное остаётся совсем немного времени и душевных сил. По идее, практика проводится для того, чтобы студенты ходили на экскурсии и разбирали собранный там материал. Экскурсии были, материал обрабатывали, но важнее, на мой взгляд, было выполнение студентами исследовательских работ. Раньше их делали весьма формально. В последние годы мы с коллегами меняем ситуацию, и работы студентов становятся всё серьезнее. По итогам прошлогодней практики у студентов вышло 12 публикаций (тезисов) с моим научным руководством (без моего авторства, естественно) и одна статья с моим участием. Посмотрим, сколько получится в этом году.
И прошлогодняя статья, и три студенческие работы в 2013-му были посвящены изучению флуктуирующей асимметрии. Довольно популярная тема, вполне простая методика… Одна проблема: по-моему, никто до конца не понимает, что именно оценивают, измеряя флуктуирующую асимметрию. Попытаюсь разобраться в этом здесь — и для читателей, и для «флуктуирующих» студентов, и для себя самого.
Начну с мысли великого австрийского (американского) биолога Людвига фон Берталанфи. Организм — динамическое выражение онтогенеза, процесса самостановления. Изучая свойства организмов, полезно задумываться, как они задаются, что является их причиной. Увы, обычно мы не можем сказать это определённо. Может быть, интересующее нас свойство — результат реализации наследственной программы, а может быть — случайная вариация или результат воздействия внешней среды.

Людвиг фон Берталанфи (1901–1972), создатель общей теории систем, и одна из принадлежащих его перу мыслей
Хорошо, когда мы можем изучить изменчивость генетически идентичных организмов, клонов. Когда такие исследования удавалось проводить, регистрировалась весьма широкая внутриклональная изменчивость. Каждый клон характеризуется не специфичными для него значениями тех или иных признаков, а распределением их вероятностей!
Последнюю мысль можно пояснить с помощью метафоры эпигенетического ландшафта, принадлежащей крупнейшему английскому теоретику биологии К. Х. Уоддингтону.
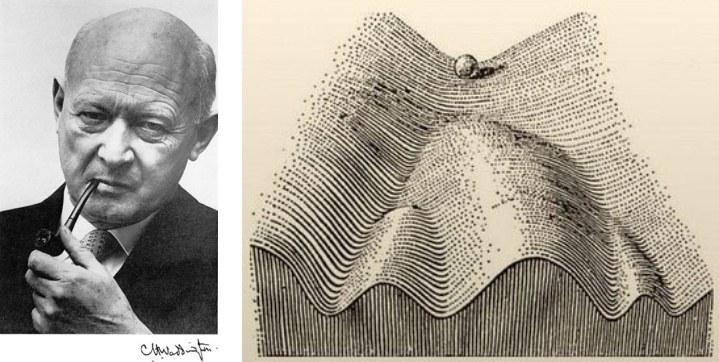
Конрад Хэл Уоддингтон (1905–1975) и его метафора эпигенетического ландшафта, предложенная в 1957 году
Онтогенез в этой метафоре уподобляется скатыванию шарика по сложной поверхности. Судить о её форме можно по распределению вероятностей исходов развития. Каждый генотип и каждый клон характеризуются своей формой такой поверхности.
Хорошо изучать организмы, которые не сложно клонировать, например, путём вегетативного размножения у многих растений, партеногенеза у тутового шелкопряда или, возможно, скрещивания полуклонов у зелёных лягушек. А что делать, если клонирование невозможно или затруднено? Генетики человека прямо-таки охотятся за естественными клонами людей — однояйцевыми близнецами. На их изучении основан близнецовый анализ, но на решение всех задач близнецов не хватает…
Одна из альтернатив изучению близнецов связана с тем, что в организмах с билатеральной (двусторонней) симметрией есть два «клона» - правая и левая половины тела. В целом они симметричны, а в деталях наблюдаются различия, проявления асимметрии. Некоторые из них позволяют судить об устойчивости/неустойчивости развития. Распространенная классификация типов асимметрии предложена известным американским теоретиком биологии Ли Ван Валеном.
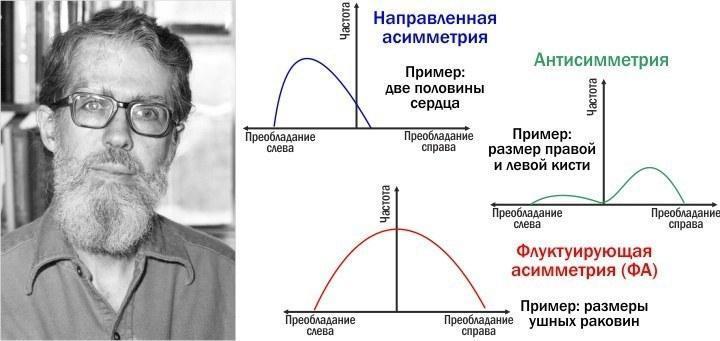
Ли Ван Вален (1935–2010) и его классификация типов асимметрии, предложенная в 1962 году
Чтобы понять, с каким типом асимметрии мы имеем дело в том или ином случае, нам надо рассмотреть распределение отклонений от симметрии в правую и левую сторону. Некоторые структуры демонстрируют устойчивое отклонение в каком-то направлении. Например, всем известно, что сердце у человека находится слева. На самом деле ситуация несколько сложнее: у человека два сердца, правое и левое, и каждое состоит из предсердия и желудочка. Правое сердце прокачивает кровь через малый, легочный круг кровообращения, а левое — через большой, охватывающий все тело. Естественно, левое сердце в норме крупнее правого, и вся парная структура смещена влево. Это функциональная асимметрия.
У меня кисть правой руки значительно крупнее левой. Вероятно, это связано с тем, что я как правша больше её нагружаю - а следовательно, интенсивнее тренирую. У левшей чаще всего крупнее левая кисть. Значительно реже встречается ситуация одинакового развития обеих структур. Это пример антисимметрии, когда симметричность структуры встречается реже, чем преобладание какой-то из сторон.
Интереснее ситуация с формой ушных раковин. У большинства людей уши относительно одинаковы, но у некоторых крупнее правое, у иных — левое. Небольшие различия в размерах встречаются чаще, значимые — реже. Весьма вероятно, что эти различия являются проявлением флуктуирующей асимметрии (сокращенно — ФА). Итак, ФА — это проявление случайных и ненаправленных отклонений от строгой билатеральной симметрии, результат неустойчивости (точнее — неидеальной устойчивости) развития. Её можно рассматривать как макроскопические последствия микроскопических стохастических событий.
Оптимистичный взгляд на ФА связан с тем, что, находя удачно отражающие её признаки, мы получаем замечательный инструмент для исследования устойчивости развития.
Представьте себе огромный берёзовый лес. Каждый лист каждой берёзы — билатерально симметричный орган. Мы собираем из разных мест этого леса небольшие выборки листьев, измеряем их асимметрию и узнаём, где в этом лесу расположены проблемные точки: их маркирует возрастание ФА. И загрязнение почвы, и заболачивание, и поражение паразитами вызовут сходные изменения симметрии листьев. Но как метод массового анализа с ответом на вопрос «Всё ли в норме?» оценка ФА может оказаться полезной.
Есть целый ряд факторов, которые могут влиять на устойчивость развития, а через нее - на ФА. ФА уменьшают оптимальные условия развития, чистая среда, высокая приспособленность индивида, оптимальное генетическое расстояние между его родителями. А повышают - неблагоприятные условия, загрязнения, неприспособленность особи, её появление в результате близкородственного размножения или отдалённой гибридизации.
Всё понятно? А теперь я расскажу о трёх работах, в которых я принимал то или иное участие.
Первый пример. Два года подряд на полевой практике студенты исследуют ФА рыб. В прошлом году работу выполняли на речных окунях. По её результатам опубликована уже упомянутая статья на украинском языке, и я сейчас перескажу её содержание. В этом году работу с подобным дизайном сделали и на окунях, и на лещах; прошлогодние результаты убедительно подтверждены.
Мы наловили достаточно большую выборку мелких окуней, построили их распределение по размерам и убедились, что они относятся к трём хорошо различающимся размерным группам. Перепроверили их возраст по линиям на чешуях; выяснили, что три размерные группы соответствуют и группам возрастным. Самыми мелкими были окуни, появившиеся в год исследования, крупнее — прошлогодние, еще крупнее — позапрошлогодние. Посчитали для всех рыб справа и слева количество лучей в брюшных и грудных плавниках, а также число чешуй вдоль боковой линии. Рассмотрели, как распределены отклонения от симметрии этих счётных признаков; убедились, что они в общем соответствуют критериям ФА. Сравнили уровень ФА, характерный для рыб разных возрастов, а также (внутри каждого возраста) для средних по размеру, мелких и крупных (то есть разнящихся скоростью роста).
Выяснилось, что рыбы того года, когда проводилось исследование, были самыми асимметричными; у окуней 2012-го ФА была ниже, а 2013-го — еще ниже. Внутри каждой группы одногодков ФА была ниже всего у рыб со средней скоростью роста, а у тех, что росли медленнее или быстрее, — несколько выше.
Мы интерпретируем это так. Рыбы различаются по устойчивости развития (и связанной с ней ФА). Более устойчивые (и более симметричные) особи имеют больше шансов дожить до следующего года. Те, кто растёт с нормальной для данной популяции скоростью, оказываются и более устойчивыми, и более симметричными. Таким образом, мы зарегистрировали и связь ФА с жизнеспособностью, и проявления стабилизирующего отбора, устраняющего из популяции более асимметричных представителей.
Второй пример. Моя кандидатская диссертация была посвящена популяционному разнообразию зелёных жаб. В ней детально исследованы 26 выборок зелёных жаб, собранных в Харьковской области и в её недалеких окрестностях. Среди изученных признаков были и те, по которым можно характеризовать ФА: симметричность/асимметричность рисунка и форма надлопаточных желёз (паротид) — производящих яд органов, расположенных в передней части туловища сзади и по бокам от головы.
26 выборок — это 26 местообитаний, где я успешно ловил жаб. Из них мне острее всего запомнилось одно, которое мы нашли во время наших ночных поездок. Останавливаешься в хорошем месте и слушаешь: не доносится ли откуда-нибудь пение зеленых жаб? Услышав их трели, отправляешься искать источник…
Это была крупная выкопанная в глине яма, заполненная бурой зловонной жидкостью: я вначале думал, что фекальными массами, а позже узнал, что это были отходы сахарного производства. С одного края в эту яму сбрасывали негашёную известь — вероятно, чтобы погасить кислую реакцию её содержимого. Спокойно дышать возле этой ямы было невозможно, все время приходилось бороться с рвотными позывами. Тем не менее в ней нерестились жабы!
Я был в бродовом костюме, закрывавшем тело по грудь. Слез в эту яму, страшно опасаясь поскользнуться и зачерпнуть её содержимое в костюм, и залез в ужасную жидкость по пояс. Я наловил хорошую выборку жаб, вылез, дошел до пруда и долго там отмывался, пока коллеги, ожидавшие меня в машине в отдалении от ямы, не разрешили мне к ним приблизиться. Так или иначе, собранная там выборка вошла в мой диссертационный комплект.
И это была самая симметричная выборка жаб, которую я держал в руках! Вероятно, будучи головастиками, они развивались в столь же ужасающих условиях — и оказались более симметричными, чем любые другие. Вы предположите, что это случайность? Кроме прочего, я убедился, что выборки зелёных жаб, собранные в черте города, оказываются более симметричными, чем привезённые из относительно «чистых» загородных местообитаний.
Сработает ли для этого эффекта то же самое объяснение, что и в предыдущем случае, — устранение более асимметричных особей отбором? Мне кажется, что нет. Если отходы сахарного производства и негашёная известь, а также городские загрязнения влияют на развитие головастиков, получающиеся жабы должны становиться асимметричнее. Может быть, в «грязных» местообитаниях потомство жаб прореживается с такой силой, что остаются лишь самые симметричные особи? Это потребовало бы столь высокой смертности, что такие популяции не смогли бы выжить.
А если не отбор, то что? Такое впечатление, что у зелёных жаб загрязнение включает некий механизм, берущий развитие под жесткий контроль и повышающий его устойчивость, несмотря на дисбалансирующий эффект внешних воздействий.
Третий пример. Комментатор предыдущей колонки веселился над тем, что я готов писать статьи «Пользование тачскрином: опыт зелёных лягушек» и «Аналитика финансовых рынков с точки зрения зелёных лягушек». Ну понятно же, что мне захотелось изучить уровень ФА у зелёных лягушек!
Я работаю в регионе, для которого характерно совместное проживание межвидовых гибридов зелёных лягушек и одного из родительских видов. Среди гибридов есть те, которые имеют 2 и 3 хромосомных набора. Естественно ожидать, что развитие гибридов (которые обладают хромосомными наборами, относящимися к двум разным видам) должно быть менее устойчивым, чем развитие представителей родительских видов. Особой неустойчивости развития следует ожидать от гибридов с тремя хромосомными наборами. Эти наборы эволюционировали в особях родительского вида, имевших по два набора, а затем они встретились по трое в каждой клетке - и, как ни странно, могут руководить развитием особи.
Зелёные лягушки имеют очень характерный рисунок на верхней поверхности тела. Неустойчивость развития гибридов можно оценить по ФА их рисунка?
Ничего подобного. Гибриды (с 2 и 3 хромосомными наборами) и представители родительского вида, как кажется, практически не отличаются друг от друга по уровню ФА рисунка, оценённой с помощью нескольких разных мер. Более того, разные меры асимметрии рисунка очень слабо коррелируют друг с другом. Если бы на эти меры влияла неустойчивость развития, у менее устойчивых особей возрастала бы асимметрия, оцениваемая по большинству из этих мер; увы, этого не наблюдается.
Как это сочетается с такими хорошими теоретическими соображениями, которые я изложил в этой колонке? А никак. Теория теорией, но эмпирические данные — упрямая вещь.
Что в сухом остатке? Измерение флуктуирующей асимметрии — интригующий метод, разработанный, чтобы докопаться до самых интимных механизмов развития. Увы, он работает не всегда. Часто действительность оказывается «непослушной» и выдаёт результат, прямо противоположный ожидаемому. Вероятно, в таких случаях в игру вмешивается какой-то неизвестный нам фактор. Как его изучать? Хотел бы я знать…
| ← | Дмитрий Шабанов | → |
| От хаоса наблюдений к динамической типологии: обсуждение на примере популяционных систем зелёных лягушек | О чем можно судить, измеряя флуктуирующую асимметрию? | Как будут изучать биологию и другие мировоззренчески значимые науки в такой школе будущего, о которой стоит мечтать? |
| Колонка для Компьютерры #113 | Колонка для Компьютерры #114 | Колонка для Компьютерры #115 |