Українська (найновіша версія) / Русский язык (обновление прекращено)
2.14. (дополнение) Некоторые этапы истории земной жизни
Первое появление жизни при создании биосферы должно было произойти не в виде появления одного какого-либо организма, а в виде их совокупности, отвечающей геохимическим функциям жизни.
В.И. Вернадский
Распространено мнение, что на каком-то этапе истории Земли возник «первый» организм, потомки которого съели все запасы органических веществ («первичный бульон») и дали начало всем формам жизни. Конечно, такие взгляды очень наивны. Для возникновения отдельного организма нужна была невероятная случайность. Жизнь возникла в результате постепенного усложнения геохимических циклов в результате отбора автокаталитических реакций, обеспечивавших их отдельные этапы. Еще до возникновения живых организмов в этих циклах происходило как образование, так и разрушение органических веществ. Это означало, что жизнь возникала не в форме отдельных организмов, а в форме обеспечивающих круговорот веществ экосистем.
Биохимические «новшества», возникшие на одном этапе геохимического круговорота, могли передаваться и на другие этапы. Так должны были распространиться способы запасания энергии, матричного синтеза биополимеров и, наконец, клеточной организации живых систем.
В современных экосистемах происходит круговорот, основой которого является образование органических веществ автотрофами и их разрушение гетеротрофами. Отходы жизнедеятельности каждой из этих групп организмов являются ресурсами для другой группы. Их замечательное соответствие друг другу — следствие того, что такой круговорот старше самой жизни.
Одна из самих давних осадочных пород, известных современной науке, найдены в формации Исуа в Гренландии. Возраст этих пород — 3,7-3,8 миллиарда лет. Эти породы образовывались на глубине, рядом с аналогом черного курильщика; в них найдены углеродные включения. Изотопный анализ этих углеродных включений свидетельствует, что они являются остатками живых организмов. Самые древние остатки организмов, имеющие сложную структуру, найдены в Австралии, в формации Варравуна, имеющей возраст 3,5 млрд. лет, и в Южной Африке (формация Онфервахт, 3,4 млрд. лет; рис. 2.10.1). Это цианобактерии, весьма похожие на современные. Это сходство касалось даже биохимии. Так, в породах, образовавшихся 3,1 млрд. лет, найдены продукты распада хлорофилла и характерный исключительно для цианобактерий пигмент фикобилин.
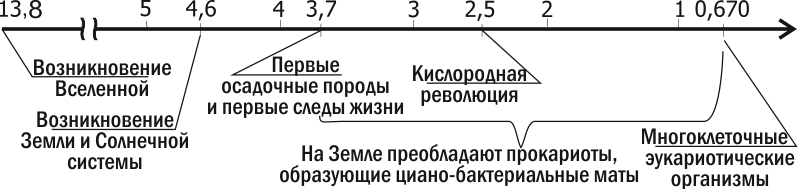
Рис. 2.10.1. Основные этапы эволюции жизни на Земле
Для протерозоя (а в некоторой степени и архея) характерны осадочные породы, которые называются строматолитами (буквально «каменными коврами»). Они имеют слоистую структуру и часто откладывались отдельными глыбами. Происхождение строматолитов долго оставалось неясным. Их образование прояснила находка современных строматолитообразователей в Заливе Акул (Шарк-Бей) в Австралии. Это изолированная от океана лагуна с очень соленой водой. На мелководье расположены осадочные глыбы, поверхность которых покрыта циано-бактериальным матом. На его поверхности находятся цианобактерии, а под их слоем — очень разнообразные бактерии других групп, а также архебактерии. Чтобы показать эту двойственность, мат называют циано-бактериальным, показывая с помощью дефиса, что он состоит из цианобактерий и других бактерий.
Минеральные вещества, которые оседают на поверхность мата и образуются при его жизнедеятельности, откладываются слоями (примерно 0,3 мм в год) на ее основание.
Другой тип бактериальных экосистем известен на территории Украины. На Арабатской стрелке, косе в заливе Сиваш, расположены заливаемые водой и пересыхающие лиманы. На поверхности почвы в них расположен циано-бактериальный мат толщиной до нескольких сантиметров.
Живые организмы не только зависят от среды своего обитания, но и сами влияют на нее. Земля исходно имела атмосферу восстановительного характера, в которой были устойчивы окисленные газы (углекислый газ CO2, водяной пар H2O, оксид серы SO2) и газы-восстановители (угарный газ CO, водород H2, сероводород H2S, аммиак NH3, метан CH4, циан HCN, хлороводород HCl и др.). В течение длительного периода истории Земли на ее поверхности могли образовываться легко окисляемые породы, такие, как графит (C), лазурит (Na2S), пирит (FeS2) и другие.
Первые организмы Земли, как автотрофы, так и гетеротрофы, были анаэробами (приспособленными к жизни в бескислородных условиях). В ходе фотосинтеза автотрофов выделялся свободный кислород (O2), который был токсичным для анаэробных организмов. Первоначально он быстро окислялся восстановителями, которые в избытке находились в среде. После того как кислород окислил основной запас восстановителей в среде, сложились относительно нейтральные условия. В результате бактериальные экосистемы приспособились к существованию в условиях избытка кислорода, и распространение получили аэробы (организмы, живущие в кислородных условиях). Поскольку фотосинтез происходил в воде, кислород мог окислять растворенные в ней вещества, способствуя их осаждению. Важнее всего, что при этом происходило окисление двухвалентного (хорошо растворимого) железа до трехвалентного, выпадавшего в осадок. Так образовывались джеспилиты — полосчатые железные руды, являющиеся важнейшим источником этого металла для современного человечества. Они состоят в основном из гематита (Fe2O3) и магнетита (FeO×Fe2O3).
С распространением фотосинтетиков-аэробов накопление кислорода в атмосфере продолжилось. Около 2 млрд. лет назад гравитационная дифференциация Земли привела к тому, что практически все не связанное в осадочных породах железо ушло в ядро нашей планеты. Прекращение поступления железа на поверхность Земли означало, что живые организмы могут окислить практически всю биосферу и накопить избыток кислорода в атмосфере. Этот перелом (происходивший 2,5–2 млрд. лет назад; рис. 2.10.1) называется кислородной революцией. Впрочем, не следует думать, что такая революця была одномоментным изменением. Она проходила через длительное балансирование, когда в разных частях биосферы складывались кое-где окислительные, кое-где восстановительные условия.
Кислородная революция была важнейшим переломом в истории Земли. Поменялся не только состав атмосферы, но и состав пород, формирующихся на поверхности Земли. Следствием кислородной атмосферы стало образование озонового слоя в атмосфере — предпосылка для заселения суши.
И для прокариот, и для эукариот характерно образование сложных систем. У прокариот это бактериальные экосистемы из тесно связанных особей разных видов и даже царств (как эубактерий, так и архебактерий). Морфофункциональное различие клеток в них — следствие их независимой эволюции. У эукариот это многоклеточные организмы — клоны потомков одной клетки, различия между которыми обусловлены реализацией различных вариантов одной и той же наследственной программы. Для развития эукариотической многоклеточности (точнее - многотканевости) требуется намного большая сложность управляющих клеточных систем.
В последние годы удалось убедительно доказать симбиогенетическую теорию происхождения эукариотической клетки. Каждая эукариотическая клетка содержит геномы различного происхождения: в клетках животных и грибов это геномы ядра и митохондрий, а в клетках растений — также и пластид. Небольшая кольцевая ДНК содержится (по многим данным) и в базальном тельце жгутиков клеток эукариотических.
Метод молекулярных часов (учета нейтральных, с точки зрения естественного отбора, и ненаправленных изменений последовательностей ДНК) свидетельствует, что эукариоты возникли тогда же, когда и прокариоты. Несмотря на это, очевидно, что в течение значительной части истории Земли на ней доминировали прокариоты. Первые клетки, соответствующие по размерам эукариотическим (так называемые акритархи), имеют возраст 3 млрд. лет, но их природа остается неясной. Почти несомненные остатки эукариот имеют возраст около 2 млрд. лет. Лишь после кислородной революции на большей части поверхности нашей планеты сложились благоприятные для эукариот условия. Эпоха их господства началась около 1 млрд. лет назад.
Вероятно, «главным» предком эукариотических клеток стали архебактерии, которые перешли к питанию путем заглатывания пищевых частиц. Изменение формы клетки, необходимое для такого заглатывания, обеспечивал цитоскелет, состоящий из актина и миозина. Наследственный аппарат такой клетки переместился вглубь от ее изменчивой поверхности, сохранив при этом свою связь с мембраной. Это послужило причиной возникновения ядерной оболочки с ядерными порами (связанной благодаря эндоплазматической сети с наружной мембраной клетки).
Бактерии, поглощенные клеткой-хозяином, могли продолжить свое существование внутри нее. Так, предками митохондрий стала группа фотосинтезирующих бактерий, приспособленных к жизни в кислородных условиях — пурпурные альфапротеобактерии. Внутри клетки-хозяина они утратили способность к фотосинтезу и приняли на себя окисление органических веществ. Благодаря им эукариотические клетки стали аэробными. Симбиозы с другими фотосинтезирующими клетками стали причиной приобретения растительными клетками пластид. Вероятно, жгутики эукариотических клеток произошли в результате симбиоза клеток-хозяев с бактериями, которые, как современные спирохеты, были способны к извивающимся движениям.
Современная фауна дает множество примеров существования проглоченных органелл и клеток в цитоплазме клеток хищника. «Выращивают» в себе проглоченные водоросли некоторые инфузории, радиолярии, кишечнополостные, плоские черви, моллюски и представители других групп. Обитающие в кишечнике термитов жгутиконосцы вступают в тесный симбиоз со спирохетами, которые прикрепляются к их поверхности. У некоторых из таких симбиогенных комплексов зарегистрировано даже сокращение генетического материала у симбионта и попадание его в зависимость от веществ, синтезируемых клеткой-хозяином. Похожие процессы происходили и при возникновении эукариотических клеток.
Первоначально наследственный аппарат эукариотических клеток был устроен примерно так же, как у прокариот (на этой стадии до сих пор находятся динофлагеляты, группа одноклеточных жгутиковых водорослей). Позже, в связи с необходимостью управления более крупной и сложной клеткой, изменилась организация хромосом, а ДНК оказалась связана с белками-гистонами. Прокариотическая организация сохранилась у геномов внутриклеточных симбионтов, однако часть их функций (у жгутиков — почти все) оказалась передана ядерному геному.
Различные группы эукариотических организмов возникали в результате различных актов симбиогенеза. В результате симбиогенеза эукариотической клетки с цианобактериями возникли красные водоросли. Зеленые водоросли возникли в результате симбиоза с бактериями-прохлорофитами. Эта недавно обнаруженная группа включает всего несколько современных видов, но зато находится в близком родстве с хлоропластами зеленых водорослей и высших растений. Наконец, хлоропласты золотистых, диатомовых, бурых и криптомонадовых водорослей возникли вследствие двух последовательных симбиозов, о чем говорит наличие у них 4 мембран. Эндосимбионтами их предков стали эукариоты, внутри которых находились симбиотические золотистые бактерии.
Развитие жизни привело к коренному преобразованию земной поверхности. Что мы увидим, оглядевшись вокруг себя за пределами человеческих поселений? Тот или иной ландшафт, покрытый характерной для каждого региона растительностью. В подавляющем большинстве мест горные породы покрыты слоем почвы. Кое-где земную поверхность прорезают водотоки — ручьи и реки. Животные заметны гораздо менее растений, но, присмотревшись к растительности, мы почти наверняка увидим насекомых, а подняв глаза вверх, разглядим птиц на фоне голубого неба. Что в этой картине является следствием обитаемости нашей планеты? Все! Осваивая сушу, жизнь существенно изменила ее. Даже голубой цвет неба является следствием накопления в атмосфере кислорода. Результатом действия живых организмов является почва. Она удерживает на поверхности суши воду и биогены в форме, оптимальной для их потребления организмами.
Плащевой сток с континентов, при котором вода двигалась многими непостоянными руслами, был заменен русловым. До заселения суши дождевая вода собиралась в потоки, которые размывали и выносили остатки горных пород. Рыхлые породы быстро сносились к океану, где ток воды резко замедлялся, и начиналось отложение осадков. В современном мире подобные условия возникают там, где реки, несущие воду с большим количеством частиц, впадают в море. Это приводит к формированию речных дельт — вдающихся в море участков «ни суши, ни моря». Примером таких ландшафтов может быть дельта Дуная. Сейчас такие участки покрывает буйная растительность, которая стабилизирует в них временные русла. До появления сосудистой растительности этого эффекта не было, и приливно-отливные волны должны были все время преобразовывать такую переходную среду, облегчившую выход из воды на сушу. Суша же представляла собой подвергающиеся интенсивному выветриванию остатки твердых пород. Массовый выход на сушу явился результатом мутуализма (см. пункт 4.7) растений и грибов — микориза обнаружена еще у риниофитов. Пройдя через промежуточную среду, растительность заселила континенты. Остатки растительных тканей послужили основой для формирования почвы. Почвенный покров закрыл от выветривания горные породы, а растения защитили почву от размывания. Сток с поверхности континентов стал русловым. Почва задерживает дождевую воду, а листовая поверхность увеличивает площадь для ее испарения (примерно в два раза превышая площадь поверхности суши). Водный обмен на континентах оказывается изменен, причем в благоприятном для организмов направлении.
Дополнительные материалы:
Учебная модель: Кислородная революция