  |
||||
|
|
Дмитро Шабанов, Марина Кравченко. |
|
||
|
I. Vertebrates Zoology |
IV.01. Походження чотириногих, історія та основні викопні групи амфібій |
IV. Amphibia sensu lato |
||
| ← III.09. Підклас "Sarcopterygii" — Лопатепері | IV.02. Походження лягвоподібної форми тіла і деякі особливості Anura → | |||
Походження чотириногих
«Найграндіознішою з усіх ризикованих витівок, що зробили хребетні за всю свою довгу історію, був, мабуть, розвиток чотириногих і вихід на сушу; цей крок був пов'язаний з корінними функціональними змінами та викликав глибокі структурні перебудови»
А. Ромер, Т. Парсонс, 1992, т. 1, с. 80
Ми розглянули ріпідістій, які були пристосовані до виходу на сушу. Як здійснювався цей перехід? Якими були проміжні між рибами і чотириногими істоти?
У цих уривках з реклами однієї з марок автомобілів ситуація показана, м'яко кажучи, спотворено (зате весело й наочно).
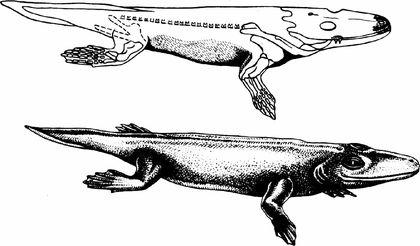
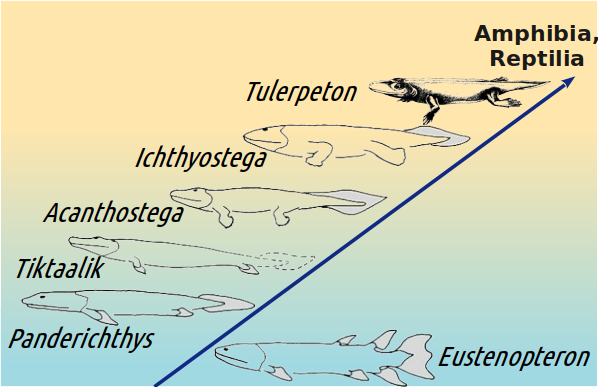
Зараз нам відомі тетраподоподібні риби (як показаний внизу пандеріхт) і рибоподібні чотириногі (як показана вище акантостега).

Ви згодні, що перехід від типових ріпідістій до елпістостегалій (між нижньою та середньою тваринами на наступному слайді) набагато зрозуміліше наступного?

Який спосіб життя вели ці перехідні форми? Як вони виглядали?
Кладістичні реконструкції філогенії, як уже вказувалося, будуються так, щоб аналізовані форми шикувалися в «економну», лінійну філогенію. Ось лінійна філогенія тварин ...
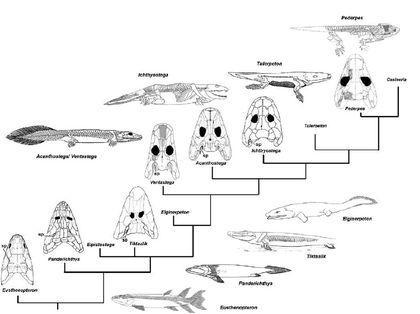
... а ось вишикувані в лінію схеми будови їх кінцівок. Чи означає це, що всі форми в ряду від риб до чотириногих використовували їх однаково, і просто все більше і більше удосконалювалися в такому пересуванні?
Щоб відповісти на це питання, варто проаналізувати наступний уривок з фільму BBC.
Знято, звісно, красиво, але не цілком достовірно. Почати слід з того, що для того, щоб так піднятися на ногах, та ще й тримати хвіст паралельно поверхні, як це робить показана тут чотиринога тварина (мається на увазі Hynerpeton), потрібно мати дуже глибоко перетворену опорнорухову систему. Від перших чотириногих очікувати такої прудкості не доводиться. До речі, гінерпетон — відомий по фрагментарним залишкам північноамериканський верхньодевонський вид; на наступному слайді показані всі відомі на сьогодні кістки цієї тварини і його художньою реконструкцією.

А чи правильно рухається хінерія (Hyneria), гігантська ріпідістія із групи Rhizodontiformes? Протиріччя містяться в самому фільмі. Тіло хінерії в самому фільмі сплощене латерально, з боків (і, ймовірно, саме таким воно і було насправді). Важила така риба дуже багато. Вагу різодуса (Rhizodus), її близького родича, реконструкція якого показана на наступному слайді, оцінюють приблизно в дві тонни. Якби такі риби спробували переміщатися на суші так, як показано у фільмі, вони пошкодили б собі ребра. Основна мускулатура цих риб згинала їх тіла з боку в бік, а не вгору-вниз. Глибоко невірно реконструювати переміщення хінерії по суходолу за аналогією з мулястими стрибунами (маленькими рибками з тілом, що сплощене дорзолатерально, тобто в спинночеревному напрямку).

Звичайно, хінерія взагалі не вилазила на сушу. Порівняння з китом-вбивцею (косаткою), наведене у фільмі, є неправильним — біомеханічно косатка «зроблена» набагато краще, але і для неї часткове вистрибування на берег є дуже важким і надзвичайно небезпечним.
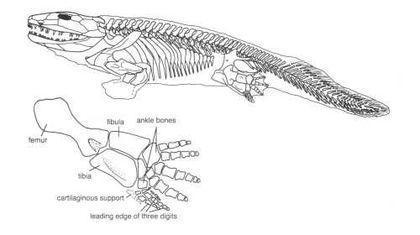
А як повзали ріпідістії, які дійсно виходили на сушу, як пандеріхтіс і тіктаалік?
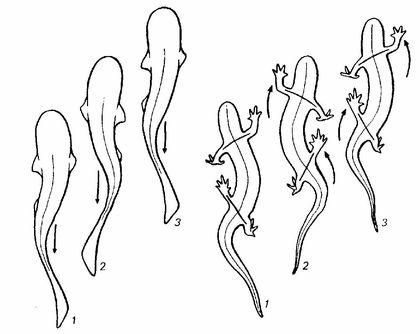
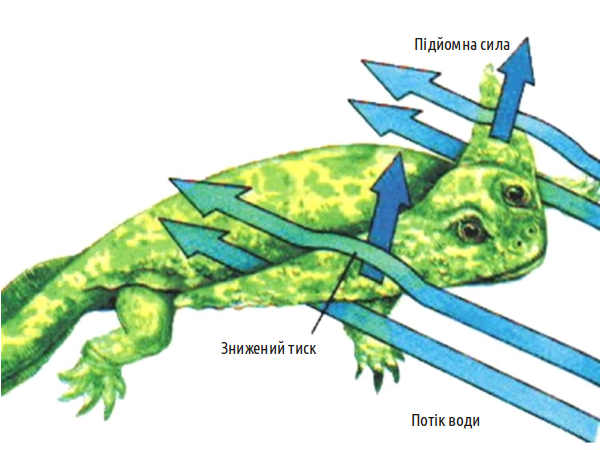
Зверніть увагу: при переміщенні примітивного чотириногого і риби задіяна мускулатура, яка забезпечує бічні вигини тіла.
Риби на зразок пандеріхтіса використовували спосіб пересування, який називається «концертино»: тіло вигинається з боку в бік, і при цьому плавець того боку, в яку здійснюється вигин, відштовхується від опори і просуває тіло вперед.

Простий спосіб уявити рух концертино — роздивитися його використання арапаїмою, Arapaima gigas, яка потрапила на суходіл.
В відео з хінерією є ще одна неясність. Як така риба могла захоплювати їжу, що знаходиться на суходолі? Не випадково в той момент, коли риба хапала свою жертву, «оператор» відвів в сторону «камеру» (якщо ви вирішили, що він це робить, щоб пощадити ваші почуття, ви попалися у пастку). А тим рибам, які дійсно виходили на суходіл, доводилося брати свою здобич бічним рухом, як пандеріхтісу на цій ілюстрації.

В іншому випадку таким рибам доводилося б брати тільки ту здобич, яка послужливо відірветься від землі, як багатоніжка на наступній реконструкції тіктааліка.

Ймовірно, конструкція тіла тіктааліка була оптимальна для того, щоб хапати здобич різким ривком (з відштовхуванням плавцями-ногами від дна) на мілководді, а також переповзати суходолом.
Отже, ми дізналися, якими були «останні» риби (з цієї еволюційної гілки, на якій з часом з’явилися й ми самі). А перші чотириногі?
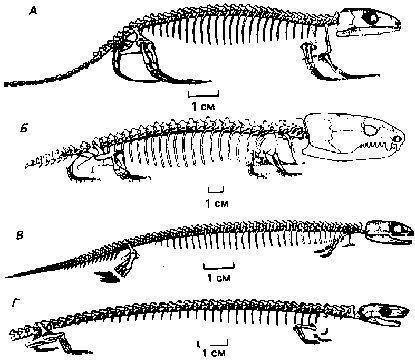
Їх в даний час відомо досить багато, але більша частина — лише за фрагментарними залишкам. Ми розглянемо тут чотири (не рахуючи згаданого раніше гінерпетона) кандидати на роль найдавнішого відносно вивченого чотириногого: акантостегу (Acanthostega gunneri), вентастегу (Ventastega curonica), іхтіостегу (Ichthyostega, описано кілька видів цього роду) і тулерпетона (Tulerpeton curtum).
Акантостега — тварина завдовжки близько 60 см, знайдена в верхньому девоні Гренландії.
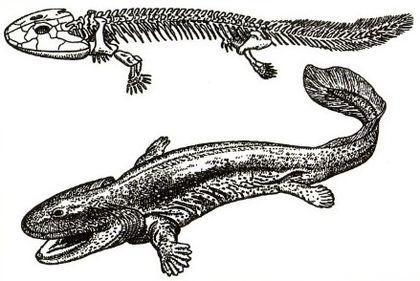
Її ноги з вісьмома пальцями були зручні швидше для загрібання, ніж для ходьби.
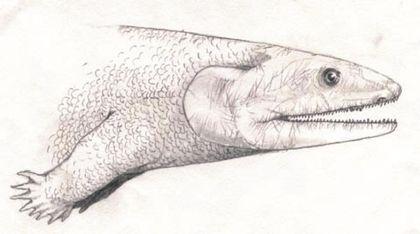
Після довгих суперечок про те, як виглядала акантостега, її стали реконструювати приблизно так:
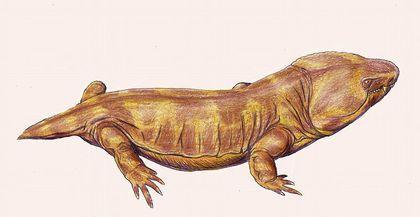
Отже, ноги акантостеги сформовані не пересуванням суходолом. А чим? Скоріше за все, переміщенням по мілководдю, як на цій реконструкції гінерпетона.

Голова акантостеги, ймовірно, була широкою і плоскою.
Вентастегу знайшли в Латвії. Ця тварина, досить схоже на акантостегу, але відрізняється від неї будовою черепа. Кінцівки вентастеги невідомі (і їх реконструюють за аналогією з акантостегою).
Ось як її реконструюють:
А це та сама реконструкція, тільки вписана в пейзаж і показана поруч з фрагментом черепа.

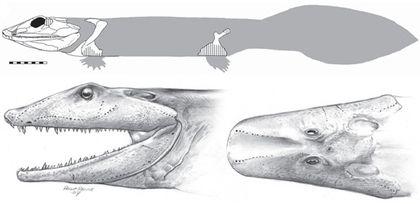
Іхтіостеги були описані раніше всіх ранніх тетрапод. Вони були виявлені в 1930-х роках в Гренландії. Це були досить великі, приблизно півтораметрові хижаки. Ось стара реконструкція іхтіостеги...
... а ось нова.
Звертає на себе увагу жорстка грудна клітина, посилена завдяки виростам на ребрах. За деякими даними, іхтіостега дуже погано згинала тіло в бічному напрямку (і, може бути, дійсно пересувалася на зразок BBC-шной хінерії).
А в воді іхтіостеги плавали завдяки «риб'ячому» хвосту.

Перераховані тетраподи схожі один на одного і відносяться не тільки до одного ряду Ichthyostegalia, але і до одного сімейства Ichthyostegidae. Череп іхтіостегалій досить спеціалізований: в ньому втрачені деякі кістки, які зберігаються у значно більш пізніх амфібій (втрачена межсконева кістка, а дві задньотім'яні кістки злилися в одну непарну кістку).
Тулерпетон у багатьох відношеннях не схожий на цих тварин.
Ця тварина близько 60 сантиметрів завдовжки, знайдена в морських верхньодевонських відкладеннях в Тульській області. На передній кінцівці 6 пальців, на задній, ймовірно, 7 (або 6).

Цікава деталь. Тулерпетон, мабуть, зберігав зв'язок плечового пояса з черепом, характерний для риб, в той час як, наприклад, іхтіостега цей зв'язок втратила. Про що говорить такий зв'язок? Про те, чи був в історії даної тварини період регулярного переміщення по суші.
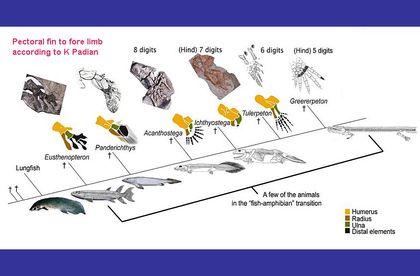
Спробуємо зібрати воєдино міркування, пов'язані з будовою кінцівок. Зараз зрозуміло, з якого «матеріалу» утворилися кінцівки.
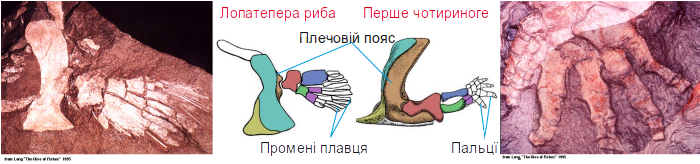
Складніше зрозуміти, навіщо вони знадобилися. Для ходьби суходолом? Але відомий ряд первинноводних чотириногих, які ніколи по суші не ходили, а кінцівки у них залишилися. Чи не найбільш характерним прикладом може бути ранньокарбоновий крассігірінус (Crassigyrinus scoticus), ймовірно, родич тулерпетон. Це двометровий водний, причому, як видно, первинноводний хижак. Його крихітні передні кінцівки сполечені з черепом.
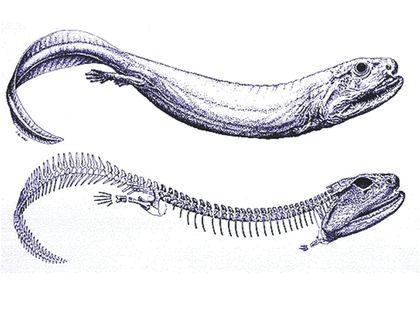
Ті, хто має досвід плавання на мілководді, добре знають, наскільки легше відштовхнути тіло від дна, ніж від води. Для такого відштовхування незручний плавник, що складається з багатьох відносно м'яких променів: вони зігнуться і поглинуть енергію поштовху. Не годиться і жорсткий плавник з однією віссю: вона просто потоне в мулі. Людина, що пливе над мулистим дном буде відштовхуватися від нього рукою з широко розчепіреними пальцями, використовуючи кілька відносно міцних осей — пальців. Яка кількість пальців оптимально? Залежно від особливостей дна та від жорсткості самих пальців. Реалізовані у ході еволюції варіанти — від 8 до 4.
При такому використанні плавця-кінцівки риб зручно згинати його, виносити вперед, а потім відштовхуватися від дна.
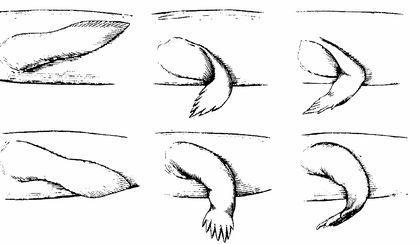
Отже, кінцівки виникають у хижаків, що переміщаються по мілководдю (другий зверху малюнок на наступному слайді). А що буде, якщо таку тварина опиниться на суходолі? Нічого доброго. Еластичний хребет зігнеться, черево притулиться до опори, а кінцівки зможуть відірвати від неї тільки голову (з якою пов'язаний плечовий пояс) і хвіст.
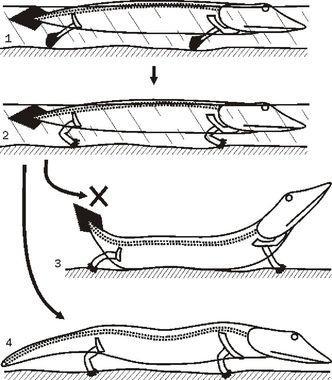
Коли чотириногі дійсно переходять до наземного переміщення, з ними відбуваються разючі зміни. Зв'язок плечового пояса з черепом розривається. Голова виявляється балансиром, що дозволяє підняти тулуб в районі плечового пояса над опорою. До речі, при такому положенні голови значно легше стає збирати з землі їжу.
«Риб'ячий» хвіст теж подовжується, перетворюючись в довгий м'язистий хвіст, типовий для більшості гадів. Насправді, укорочення хвоста у всіх чотириногих виявляється можливим тільки тоді, коли він перестає бути балансиром. Для яких груп це характерно? Або для тих, у кого тулубовий відділ хребта укорочений і малорухомий (безхвості амфібії і птиці), або для тих, у кого його рухливість забезпечується його глибокою морфофізіологічною перебудовою, як у ссавців. Деякі ссавці зберігають м'язистий хвіст, але це відбувається в тих випадках, коли він допомагає виконувати якісь специфічні функції (опори і балансира при стрибку у кенгуру, рушія в воді у китів, опори у трубкозубів, хапального органу у панголінів тощо).
Отже, з цієї точки зору швидке освоєння суші пов'язано з пересуванням «концертино», і дало напівназемних риб. Сушу ж з часом освоїли нащадки мешканців мілководдя, які загрібали плавниками-лапами по мулу, залишаючись на плаву. Більш того, і ці тварини могли розвиватися не однією еволюційною гілкою, а кількома.
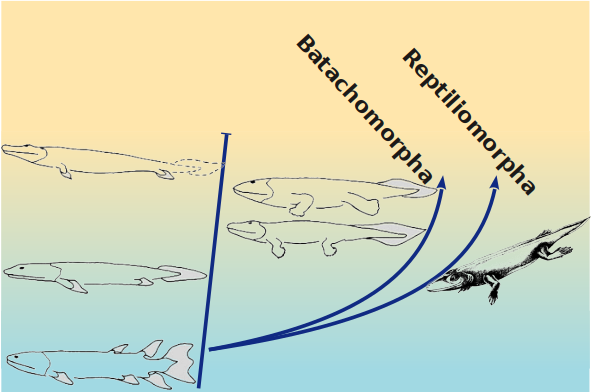
Внаслідок цього більш наближеною до дійсності може виявитися не "лінійна", найекономніша філогенія, як та, що показана на наступному рисунку. Блакитним кольором тут умовно показано водний спосіб життя, а жовтим — частково наземний
Автору цієї лекції здаються вагомими аргументи на користь "кущистої" версії філогенії, де припускається можливість паралельної тетраподизації. На якому етапі розділилися гілки, що ведуть до сучасних амфібій і сучасних рептилій — дуже складне питання. Елпістостегалії (пандеріхтіс та тіктаалік) з цієї точки зору можуть бути навіть більш наземними тваринами, ніж іхтіостегалії та тулерпетон, але при цьому не мати сучасних нащадків.
Так чи можна сьогодні твердо відповісти, які тварини були першими чотириногими? Одна з досить переконливих версій полягає в тому, що ми їх ще не знаємо. Правдоподібно виглядає припущення, що під час становлення нашого надкласу мілководдя планети були заселені багатьма формами напівриб-напівчотириногих. Ряд знахідок (наприклад, дуже древні сліди невідомих чотириногих на мулистому ґрунті) говорить про те, що описані тут тварини зовсім не були найпершими чотириногими, а, можливо, представляли різні еволюційні лінії з цілого «букету». До нашого часу збереглися лише деякі з цих ліній, і їх найперші представники можуть бути до цього часу невідомими.
Підкласи амфібій
Систематика амфібій тривалий час була предметом суперечок, і до сьогодні її не можна вважати усталеною. Перш за все, слід сказати, що сам термін «амфібії» може використовуватися в декількох різних сенсах. З одного боку, поняття «амфібії» характеризує рівень організації (граду). У цьому випадку воно використовується для позначення анамніотичних чотириногих, життя яких часто пов'язана як з водним, так і з наземним середовищем. Звичайно, це поняття характеризує групу, яка не відповідає суворим критеріям монофілії.
З іншого боку, поняття «амфібії» можна використовувати для позначення групи тварин, пов'язаних родинними зв'язками (філи, еволюційної гілки). Зараз стало ясно, що амфібійні за способом життя представники були як у власне амфібійній, так і рептильній лініях розвитку. Якщо говорити тільки про тих тварин, до яких належать сучасні амфібії і їхні викопні родичі, краще використовувати термін Batrachomorpha — батрахоморфи. У цьому курсі протягом декількох останніх років лектор вважав сучасних амфібій представниками класу Batrachomorpha. Однак більшість авторитетних прихильників компромісної еволюційної систематики вважає правильним включати до складу амфібій і безпосередніх предків рептилій, ми будемо застосувати більш поширену назву — клас "Amphibia" (лапки, як ви пам’ятаєте, свідчать, що це не справжня монофілетична група). У такому разі поняття «амфібії» тут характеризує збірну таксономічну групу, що виділяється швидше за все саме за способом життя. Ще один спосіб показати широке трактування цього терміну — назвати його Amphibia sensu lato. Sensu lato (у широкому розумінні) та sensu stricto (у вузькому розумінні) — це парні латинські вирази, що використовуються у систематиці дозволяють розрізняти такі відміни у використанні таких понять, як ті, що ми зараз обговорюємо.
А у якому разі ми можемо говорити про Amphibia sensu stricto? У разі, коли ми виключимо з групи, що розглядаємо, представників рептильної гілки. Це те ж саме, що Batrachomorpha.
На які підкласи слід розділяти Amphibia sensu lato? Найстаріша класифікація усіх (і сучасних, і викопних) амфібій на групи пов'язана з їх поділом на древніх (стегоцефалів, панцирноголових) і нових (ліссамфібій, безпанцирних або голих амфібій).
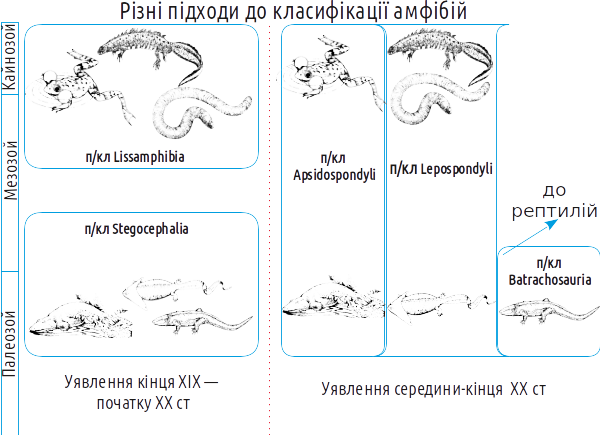
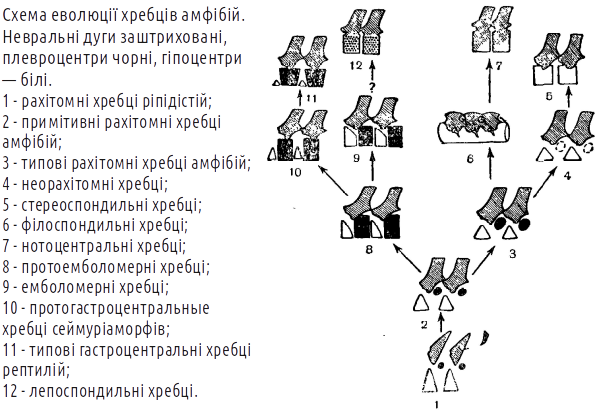
Однак з 30-х років XX століття стало ясно, що еволюція стегоцефалів проходила як мінімум двома гілками. Виявилося, що кожній з гілок викопних амфібій за низкою ознак відповідають певні сучасні групи. З ім'ям Альфреда Ромера (1884-1973), видатного американського палеонтолога, пов'язане уявлення про те, що батрахоморфи діляться на апсидоспондильних (дугохребцевих) і лепоспондильних (тонкохребцевих); предки рептилій об'єднувалися в групу Batrachosauria (жабоящери). У апсидоспондильних тіло хребця утворюється з окремих дугоподібних окостенінь (апсида — склепіння, звід), а у лепоспондильних — утворюється як єдине ціле. Здавалося, що безхвості належать до апсидоспондильних, а хвостаті і безногі — до лепоспондильних.
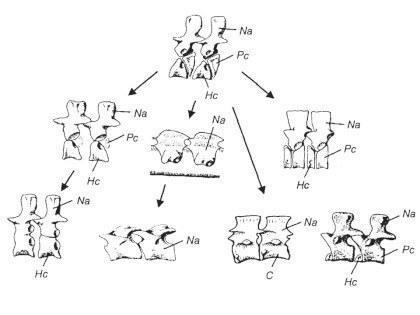
Більш того, в ході досліджень ріпідістій стало ясно, що апсидоспондильні амфібії нагадують остеолепіформних риб, а лепоспондильні — поролепіформних (тих, яких зараз вважають не ріпідістіями, а діпноморфами). З будовою хребців виявилося тісно пов'язаною будова нюхової області черепа ріпідістій і амфібій. Значить, є як мінімум дві гілки, що простягнулися від лопатеперих риб до сучасних амфібій? Не все так просто. Альтернативна точка зору полягає в тому, що основною гілкою еволюції амфібій була апсидоспондильна, а лепоспондильні представники виникали від різних апсидоспондильних предків в результаті педоморфозу (збереження личинкових ознак в дорослому стані). В усякому разі, зараз говорити про походження хвостатих амфібій від поролепіформних риб не доводиться.
Відносно недавно був знайдений пермський стегоцефал геробатрахус (Gerobatrachus), який буквально «проситься» на роль близького родича спільних предків безхвостих і хвостатих. Значить, і хвостаті амфібії належать до апсидоспондильної гілки.


А ось безногі виявилися, в тому числі за генетичними даними, досить далекими і від безхвостих, і від хвостатих. Таким чином, можна прийняти таку систему і уявлення про филогению амфібій, яка показана на наступному слайді. Еволюційні гілки, які відносяться до групи Batrachomorpha, показані зеленим кольором.
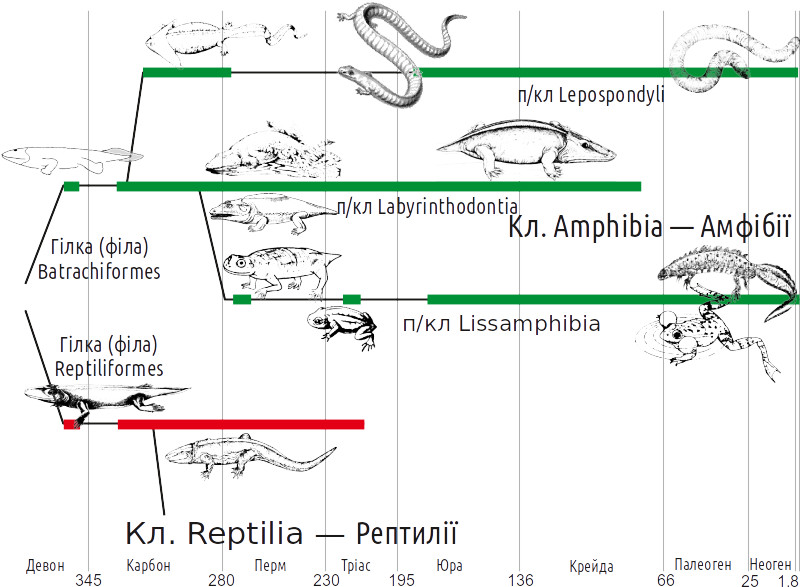
В даному курсі ми приймаємо такі ряди в складі виділених підкласів
Клас “Amphibia” (Amphibia sensu lato) — Амфібії
†Підклас “Labyrinthodontia” — Лабірінтодонти
† Ряд “Ichthyostegalia”
† Ряд “Temnospondyli” — Темноспондили, Розщепленнохребцеві
Підклас Lepospondyli — Тонкохребцеві
† Ряд Nectridea
† Ряд Aistopoda
† Ряд Adelospondyli
† Ряд Lysorophia
† Ряд “Microsauria”
Ряд Gymnophiona — Безногі
Підклас Lissamphibia — Безпанцирні
† Ряд “Proanura” — Первиннобезхвості
Ряд Anura — Безхвості
Ряд Caudata — Хвостаті
†Підклас “Reptiliomorpha” — Рептиліоморфи
† Ряд “Anthracosauromorpha” — Антракозаври
Як вже вказано, три перших підкласи можна об'єднати в монофілетичну групу Batrachomorpha =(Amphibia sensu stricto) — батрахоморфів.
Як і раніше, лапки на назвах таксонів маркують парафілетичні групи, а ті таксони, які повинні знати студенти ІІ курсу, виділені цегляним кольором. Втім, щодо гадів (амфібій і рептилій) вимоги, що висуваються до студентів, включають також знайомство з низкою родин (які будуть перераховані відповідних сторінках цього онлайн-підручника).
Крім системи, цікаво обговорити різноманітність життєвих форм амфібій. На наступному слайді мова йде про різноманітність форм батрахоморфов (втім, різноманітність рептиліоморфів в загальному укладається в ту ж схему).

Підклас “Labyrinthodontia” — Лабірінтодонти
Представників ряду “Ichthyostegalia” ми вже розглянули. Назва іхтіостегалій узята у лапки хоча б тому, що, ймовірно, темноспондили (і ще багато груп) є їхніми прямими нащадками.
Ряд Temnospondyli — Темноспондили, Розщепленнохребцеві
Звичайно, найрізноманітніша група викопних амфібій — темноспондили. На наступному слайді показано різноманітність черепів темноспондилів. Серед них були і великі, і дрібні форми, хижаки-засідники з широкими черепами і крокодилоподібні рибоядні амфібії з витягнутими черепами.

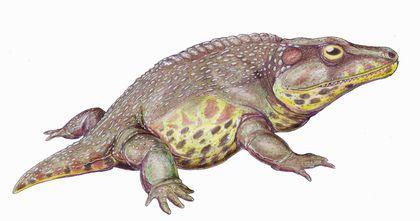
Детально розглядати різноманітність темноспондилов ми не будемо, але наведемо деякі приклади. Ось хелідерпетон (Cheliderpeton), пермський темноспондил довжиною близько 1,2 метра з території сучасної Чехії. Це був водний крокодилоподібний хижак.

Пізнекарбоновий-ранньопермський еріопс (Eryops megacephalus) — один з найвідоміших темноспонділов. Це двометровий хижак з півметровим черепом.
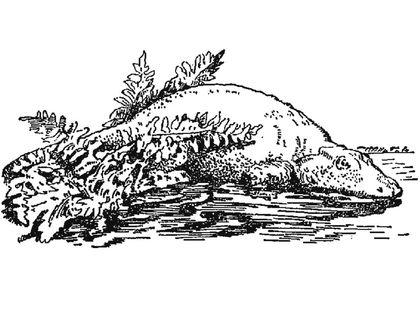
Раніше еріопса вважали напівводним хижаком, тепер, швидше за все, реконструюють як водного засідника.

Бранхіозавр (Branchiosaurus) був дрібнішою (близько метра) і також, ймовірно, переважно водною твариною.

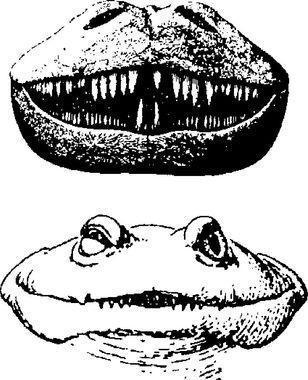
Одними з найназемніших темноспондилов були представники сімейства діссорофіди (Dissorophidae). Їх чудовим представником є какопс (Cacops aspidephorus). Розгляньте реконструкцію його скелета...

...та його прорисовану схему.
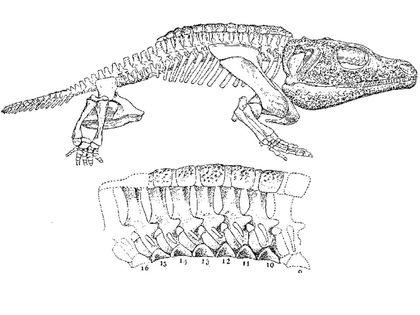
Довжина какопса була близько 50 см. Це була тварина з компактним тілом, потужними кінцівками, укороченим хвостом, великою головою з великими очима і барабанними перетинками.
Одна з версій, що пояснює особливості какопса, трактує його як берегового зістрибувача в воду, тобто раннього (і ще дуже досконалого) представника тієї ж життєвої форми, до якої належать і сучасні зелені жаби. Відхід від переслідувача одним стрибком має сенс в тому випадку, якщо тварина знаходиться на межі середовищ, і одним різким рухом можна втекти від небезпеки. Для стрибка потрібен жорсткий скелет, який передасть зусилля кінцівок на все тіло. Гнучкий хребет темноспондилів погано підходив для цього. Перебудова тіл хребців вимагала тривалої еволюції. Какопс реалізував простіший варіант, який додав його тілу жорсткісті: на остистих відростках хребців розвивався міцний спинний панцир, щось на кшталт «другого хребта».
Як буде вести себе хижак, жертва якого може піти від нього одним стрибком? Буде підкрадатися. Що потрібно робити, щоб врятуватися від хижака, що підкрадається? Вчасно його побачити чи почути. Саме цим пояснюються величезні очі і великі барабанні перетинки і какопса, і сучасних жаб.
До діссорофід належав і згаданий вище геробатрахус, і таке дивовижна тварина, як платигистрикс (Platyhystrix rugosus).

Призначення розширених остистих відростків платигістрікса лишається неясним. Коли мова йде про пелікозаврів, істот рептильного рівня організації, спиннї вітрила, утримувані подібними відростками, реконструюють як засіб терморегуляції. Чи могла використовувати такий механізм амфібія? Малоймовірно. Статевий відбір? Демонстрація хижакам збільшеного контуру тіла? Не дуже переконливо...
Деякі амфібії перетворилися в рухомих рибоїдних хижаків. Показаний на наступному слайді пермський платіопс (Platyops) досягав довжини близько метра,...
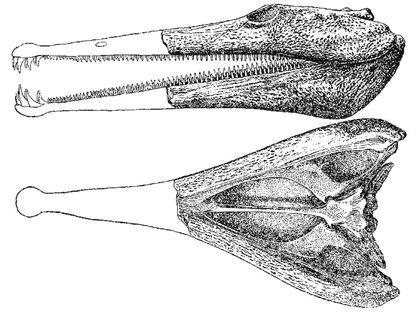
а тріасові трематозаври (Trematosauridae) мали черепа до 90 см, і, отже, сягали кількох метрів в довжину. Це були морські рибоїдні мисливці. Для сучасних амфібій морська вода є непереборною перешкодою; мезозойські темноспондили якось подолали цей бар'єр.

До цієї групи був близький кандидат на роль найбільшої амфібії — пермський пріонозух (Prionosuchus plummeri) з Бразилії, що досягав, ймовірно, 7 м. в довжину.

У пермі кількість великих амфібій на суші різко зменшилася, а в мезозої збереглися або водні, або дрібні види. Це, ймовірно, результат конкуренції з рептиліями, які у наземних умовах мають низку вагомих переваг перед амфібіями.
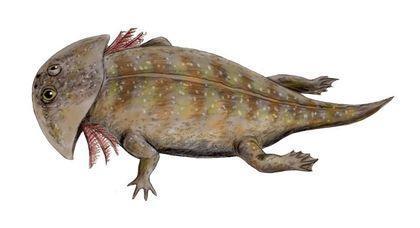
Крім морських активних хижаків, в мезозої поширилися «живі капкани», великі донні засідники. Тріасовий герроторакс (Gerrothorax) сягав у довжину близько метра. Показано, що протягом усього свого життя він, як личинка, зберігав зовнішні зябра (наслідок педоморфозу).
Витиснені в воду амфібії досить довго залишалися серйозними хижаками. Яскравим їх представником був мастодонзавр (Mastodonsaurus). Довжина його черепа досягала 1,4 м (тобто на фотографії нижче зображений далеко не найбільший), а довжина тіла, ймовірно, близько 6 м.

На піднебінні мастодонзавра знаходилися великі ікла, які явно служили для утримання серйозної здобичі.

Хоча на реконструкціях мастодонзавра часто зображують на суходолі,...

...він, ймовірно, все життя проводив у воді, різкими рухами хапаючи свою здобич.

Плоске тіло мастодонзаврід (як показаного на наступному слайді паротозуха, Parotosuchus) дозволяло їм бути непомітними на дні.

Ще більшими були брахіопоїди (Brachyopoidea), остання група темноспондилов. Відомі залишки раннеюрського брахіопоїда, довжину якого оцінюють в 7-8 м. Втім, конкуренції з крокодилами ці тварини не витримали...

Останній відомий темноспонділ — брахіопоїд кулазух (Koolasuchus cleelandi), який описаний з нижньої крейди Австралії, яка в цей час перебувала за Південним полярним колом. Його довжина сягала 4-5 м. Крокодили в Австралії не винесли б холодної полярної ночі, а кулазух, ймовірно, міг зимувати під льодом.

Цю тварину показано в «Прогулянках з динозаврами» BBC, але припущення про те, що вона виходила зимувати на сушу, здається малоймовірним. Захиститися від промерзання холодною зимою можна або на дні водойми, або в глибокій норі. Нору кулазух викопати не міг (чим копати?), а ось опуститися на дно і загальмувати обмін речовин, ймовірно, був здатним.
Підклас Lepospondyli — Тонкохребцеві
Лепоспондильні амфібії — не дуже чисельна, але різноманітна група. Дрібні та середні за розміром, великі представники відсутні. Усі вони тією або іншою мірою педоморфні, тобто зберігають в дорослому віці личинковий стан тих чи інших ознак. Саме на представниках цієї групи була описана неотенія, крайній випадок педоморфозу, коли дозріває лише одна репродуктивна система, а усі інші зберігають личинковий стан. Тіла хребців котушкоподібні. До цього підкласу відноситься кілька викопних груп і, відповідно до прийнятого тут трактування, один сучасний ряд.
На цій картині відомого чеського палеоанімаліста Зденека Буріана показані лепоспондильні амфібії і один темноспондил (бранхіозавр).

Ряд Nectridea
Постійноводні тритоноподібні амфібії довжиною до 1 м. Відомі від середнього карбону до ранньої пермі. Характерний довгий хвіст з великими невральними і гемальними дугами — ефективний рушій у воді; дуги не дозволяли йому скручуватися. Цікава деталь: деякі роди, як Urocordilus (на картині Буриана вгорі, скелет — на наступному слайді) мали по п'ять пальців в кисті, а більш пізні форми — по чотири, як усі батрахоморфи.

У деяких нектрідій, як у показаного тут діплокаулюса (Diplocaulus) розвивалися «роги» черепа, утворені виростами лускатих і таблитчатих кісток. На наступному слайді реконструкція діплокаулюса поміщена в пластмасовий таз (ніби тільки що зловили...).

Призначення цих рогів неясно. За однією з версій, вони були чимось на кшталт «підводних крил».
Ряд Aistopoda
Безногі карбонові-ранньопермські лепоспондили. Довжина до 1 м. Жили або в зарослих водоймах або на вологих «луках». Один з найвідоміших видів — Ophiderpeton, показаний тут у воді,...

... а Oestocephalus зображений тут на суші.

Ряд Lysorophia
Вугроподібні амфібії з дуже своєрідним черепом і дрібними кінцівками. Існували з карбону; один рід, за деякими даними, дожив до нижнього тріасу. Lysorophus показаний тут під час охорони кладку ікринок (по аналогії з черв’ягами, хоча особливих підстав для такої аналогії, ймовірно, немає).

А це представник іншого роду, Brachydectes.

Ряд Adelospondyli
Карбонові водні лепоспондили з подовженим тулубом довжиною близько 30 см; чи мали вони кінцівки, точно не відомо. Живилися, проціджуючи воду через тонкі зуби. Нижче показаний реконструйований Adelospondylus.

Ряд Microsauria
Дуже різноманітна група карбонових і пермських амфібій. Відомо більше десятка родин. Саламандроподібні, ящіркоподібні і навіть риючі тварини. Є й інші уявлення щодо систематичного положення та філогенетичних зв'язків цих тварин. Можливо, вони загалом є окремою гілкою чотириногих; втім, у такому разі незрозуміло, як оцінювати їх ймовірні зв'язки з сучасними безногими амфібіями.
Pantylus мав дуже потужні щелепи.
Pelodosotis, найбільший з мікрозаврів (сягав 60 см в довжину), вів риючий спосіб життя.

Ostodolepis, близький до попереднього рід. На черепі добре видно пристосування до копання в м'якому ґрунті.
А це — вже не мікрозавр, а найраніший відомий представник безногих, Gymnophiona. Це безноге (ще з ніжками), Eocaecilia, знайдено в ранній юрі Північної Америки. Вважається, що воно демонструє, що безногі є безпосередніми нащадками мікрозаврів.
Підклас “Reptiliomorpha” — Рептиліоморфи
Якщо об'єднати перші три підкласи амфібій в групу Batrachomorpha, ця група виявиться (з точки зору філогенетичної систематики) «гарною» (справжньою монофілетичною), адже вона об'єднує усіх нащадків певного предка. Підклас Reptiliomorpha в складі класу Amphibia з цієї точки зору все одно «поганий» (парафілетичний, такий, що не включає деяких нащадків видів, які до нього належать).
Ряд “Anthracosauromorpha” — Антракозаври
Склад підкласу Reptiliomorpha остаточно ще не встановлено. У деяких системах в нього (ймовірно, з достатніми підставами) включають кілька рядів. Тут ми розглянемо його в мінімальному обсязі, виділяючи один ряд Antracosauromorpha. Різні групи рептиліоморфів ми будемо розглядати в якості підрядів: Embolomeri (емболомери), Chroniosuchia (хроніозухіди), Gephyrostegida (гефіростегіди) і Seymouriamorpha (сеймуриаморфи).
Ще одна складність у трактуванні підкласу Reptiliomorpha пов'язана з тим, що не існує однозначних критеріїв, за якими його можна відокремити від наступних груп, яких ми відносимо вже до рептилій. Деякі автори включають Seymouriamorpha або їх окремих представників в число рептилій.
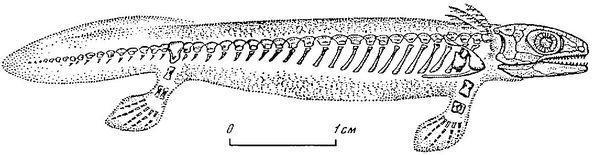
Емболомери — група середніх за розміром і великих водних і напівводних тварин, які відомі з пізнього девону до кінця пермі. Їх назва пов'язана з емболомерним типом хребців, характерним для цієї групи. Цікава особливість емболомеров (як і всіх антракозавров) — п'ятипалі і передніх, і задні кінцівки (за винятком тулерпетона, що на передніх кінцівках мав шість пальців, а на задніх — й того більше). Це яскраво відрізняє рептиліоморфів від батрахоморфів, для яких характерна чотирипалість передньої кінцівки. Для емболомерів були характерні лабіринтодонтні зуби, успадковані від ріпідістій.
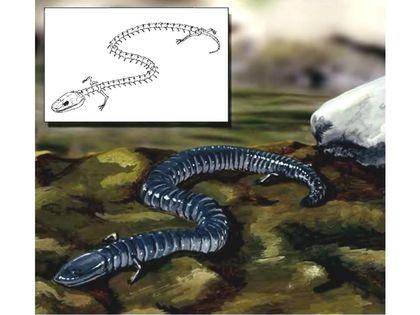
Перший представник, якого включають (не всі автори!) в число антракозаврів, — девонський тулерпетон (Tulerpeton), якого ми вже згадували, розглядаючи походження чотириногих.
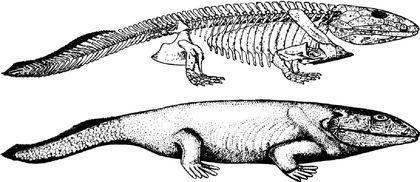
Судячи з того, що плечовий пояс тулерпетона зберігав зв'язок з його черепом, тулерпетон залишався ще первинноводною твариною. Ймовірно, він міг переміщатися суходолом, але в цілому його будова була пов'язана з пристосуванням до життя на мілководді. Багато інших емболомерів також вели водний спосіб життя. Наприклад, таким був показаний нижче Eogyrinus з нижнього карбону. Це велика, до 4,5 м в довжину, саламандроподібна або крокодилоподібна тварина, яка вела водний спосіб життя.

Вище був показаний скелет еогірінуса, а нижче розміщена його художня реконструкція.

Деякі з емболомерів спеціалізувалися саме як водні хижаки. Їх яскравим прикладом є ранньокарбоновий Crassigyrinus, що сягав довжини приблизно в 2 метри (його реконструкцію показано вище). Його редуковані передні кінцівки зберігають анатомічний зв’язок з черепом.
Далі — фрагмент фантастичного фільму BBC «Доісторичний парк». У ньому відомий англійський ведучий фільмів про тварин Найджел Марвін (нібито в ході подорожей в минуле) спочатку ловить гігантську комаху меганевру, а потім випадково виявляє крассігірінуса.
А це — Archeria, дрібний пермський водний емболомер.

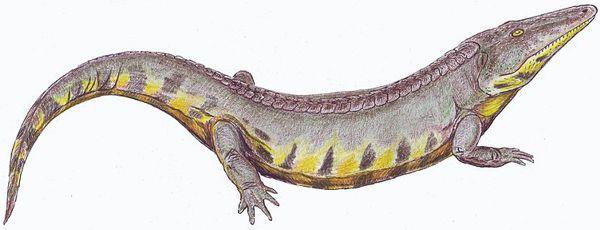
Хроніозухії — Chroniosuchia, крокодилоподібні рептиліоморфи, деякі представники яких пережили перм-тріасове вимирання і існували ще в ранньому тріасі. Йдеться про представників роду Bystrowiana, ймовірно, наземних хижаків, що сягали довжини декількох метрів (череп завдовжки 40 см):
Значний інтерес (і великі суперечки) викликають сеймуріаморфи. Самі сеймурії (Seymouria) — ймовірно, наземні тварини. Однак для їх найближчих родичів були описані водні личинки із зовнішніми зябрами. На підставі цього можна припустити, що і сеймурії теж знаходилися на амфібійному рівні розвитку. Тут показаний скелет сеймурії та художня реконструкція двох особин.


До сеймуріаморфів належать також досить різноманітні водні тварини, деякі з яких, мабуть, були неотенічними формами, що взагалі втратили наземну стадію. Одним з найбільш характерних прикладів тісно пов'язаних з водою сеймуріаморфів можуть бути пермські діскозавріски (Discosauriscus). На наступних зображеннях показаний відбиток діскозавріска і реконструкція його личинки.

До діскозаврісків близькі карпінскіозавріски (Karpinskiosauridae), які також відносяться до сеймуріаморфів. Нижче показана реконструкція Karpinskiosaurus.

З якими рептиліоморфами пов'язано походження амніот? Остаточної відповіді на це питання ще немає. Більшість відомих антракозаврів жили вже після того, як з'явилися перші рептилії. Ймовірно, кандидатами на роль наших предків є гефіростегіди.
Гефіростега (Gephyrostegus bohemicus) відома з пізнього карбону, тобто жила занадто пізно, щоб підходити на роль предка рептилій. У той же час за своїми порівняльноанатомічними особливостям буквально «проситься» на цю роль. Далі наведено зображення скелета гефіростегі і дві різні реконструкції її зовнішнього вигляду.
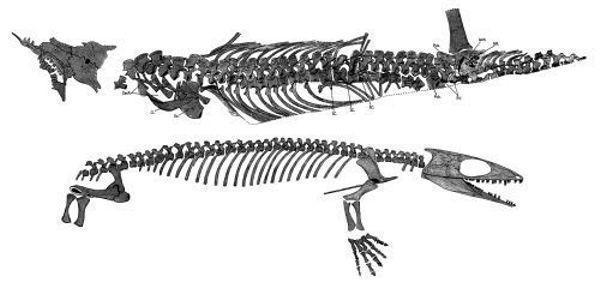
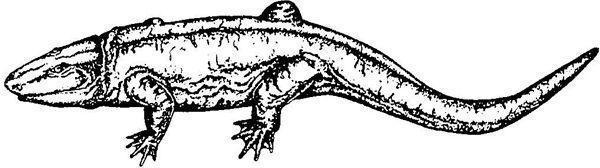
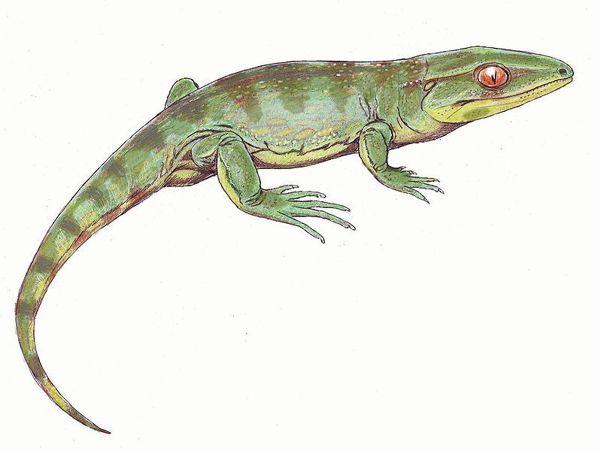
Можливо, гефіростегіди могли існувати і раніше кінця карбону. До того ж сімейства, що гефіростега, відносять ранньокарбонового бруктерерпетона (Bruktererpeton fiebigi), який теж підходить якщо не на роль ймовірного предка рептилій, то, в усякому разі, на роль близького родича такого предка.
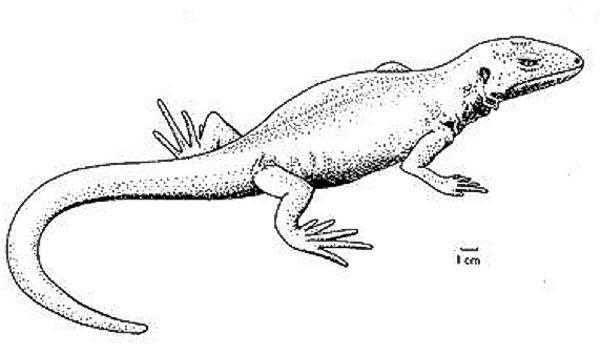
Ймовірно, в еволюції рептиліоморфів проявився сценарій, що неодноразово спостерігався в ході еволюції. При освоєнні нової екологічної зони в неї спочатку проникають групи, що еволюціонували швидко і першими придбали окремі пристосування, необхідні для нового способу життя. Таких першопроходців, використовуючи термін, введений В.О. Ковалевським, можна назвати інадаптованими. Згодом, однак, перевага може перейти до груп, які довше еволюціонували в перехідних умовах і придбали, в силу цього, більш глибокі, комплексні адаптації — евадаптованим групам. Зрештою, евадаптовані групи повністю або частково витісняють представників інадаптованих (колись більш успішних) груп. Вище в цьому розділі описаний какопс. Він був представником перших, інадаптованих зістрибувачів у воду. Какопс зник, а евадаптовані безхвості є цілком успішною групою.
Рептиліоморфи виявилися більш успішними, евадаптованими при освоєнні суходолу, ніж батрахоморфи (хоча в карбоні, як здавалося, перевагу отримали саме батрахоморфи). Одна з ключових переваг рептиліоморфів — вентиляція легенів за допомогою зміни обсягу грудної клітини. Для батрахоморфов типово, що, якщо у них взагалі є легкі, повітря в них закачується за допомогою гулярного насосу — під'язикового апарату. Це вимагає, щоб череп був широким (платібазальним), що забезпечує достатню площу горлового насоса. Мускулатура, що закриває щелепи, розташовується при цьому зверху (можливо, обмежуючи можливості для збільшення обсягу мозку); сила закривання щелеп виявляється в більшості випадків сильно обмеженою (через несприятливе співвідношення плечей важеля). Отже, механізм вентиляції легень у батрахоморфів обмежує і інтенсивність газообміну, і ефективність харчування.
Вентиляція легенів за допомогою зміни обсягу грудної клітини позбавлена цих недоліків. Мускулатура, що закриває щелепи, може перебувати збоку від них; череп стає вузьким (тропібазальним), зберігаються кращі умови для збільшення розмірів мозку. Ефективна вентиляція легенів забезпечує можливість збільшення рівня метаболізму. Перед рептиліоморфами і їх нащадками відкрилися широкі еволюційні перспективи!
Коментарі
Интересно было бы узнать
Интересно было бы узнать информация о всех вариантах преобразования рыб в тетраподов и их точность. И возможные источники информации по этому вопросу...
Вся проблема в том что ты веришь будто ты существуешь....