 |
||||
|
|
Дмитро Шабанов, Марина Кравченко. |
|
||
|
I. Vertebrates Zoology |
Історія рептилій. Філа Synapsida |
IV. Amphibia sensu lato |
||
Здається, систематика ніякої іншої групи хребетних не зазнала таких змін, не стикається з такими складнощами, як систематика рептилій. Один з авторів цього підручника багато років викладав цілком логічну систематику рептилій, яка була побудована на морфологічних та палеонтологічних даних і здавалася зрозумілою. І тут, завдяки молекулярній філогенетиці, біологія отримала величезний пласт нових даних, і стало зрозуміло, що філогенію рептилій ми розуміли явно недостатньо.
Проблемою, що зруйнувала усю систему, стало розташування у системі черепах. Багато років головним критерієм у розділі рептилій (у широкому сенсі) на групи була будова скроневих відділів їхніх черепів. Вихідний стан — цільний, анапсидний череп, зі скроневою дільницею, яку утворює суцільний щит покривних кісток, унаслідований ще від лопатеперих риб. Такий стан був характерним для предків рептилій, їх найпримітивніших представників, окремої групи, що отримала назви парарептилій, та, у дещо зміненому вигляді, у сучасних черепах. Завдяки цьому черепах включали в число парарептилій. В інших групах, для того, щоб мускулатура, яка забезпечує рух щелеп, мала простір, куди вона може розширятися при скороченні (а також просто для полегшення голови) у скроневій дільніці черепа виникали отвори — скроневі вікна. Виділяли три типи фенестрації (утворення отворів) у черепі. Для лінії, що з часом дала ссавців, є характерним утворення одного скроневого вікна. Такий череп називається синапсидним.
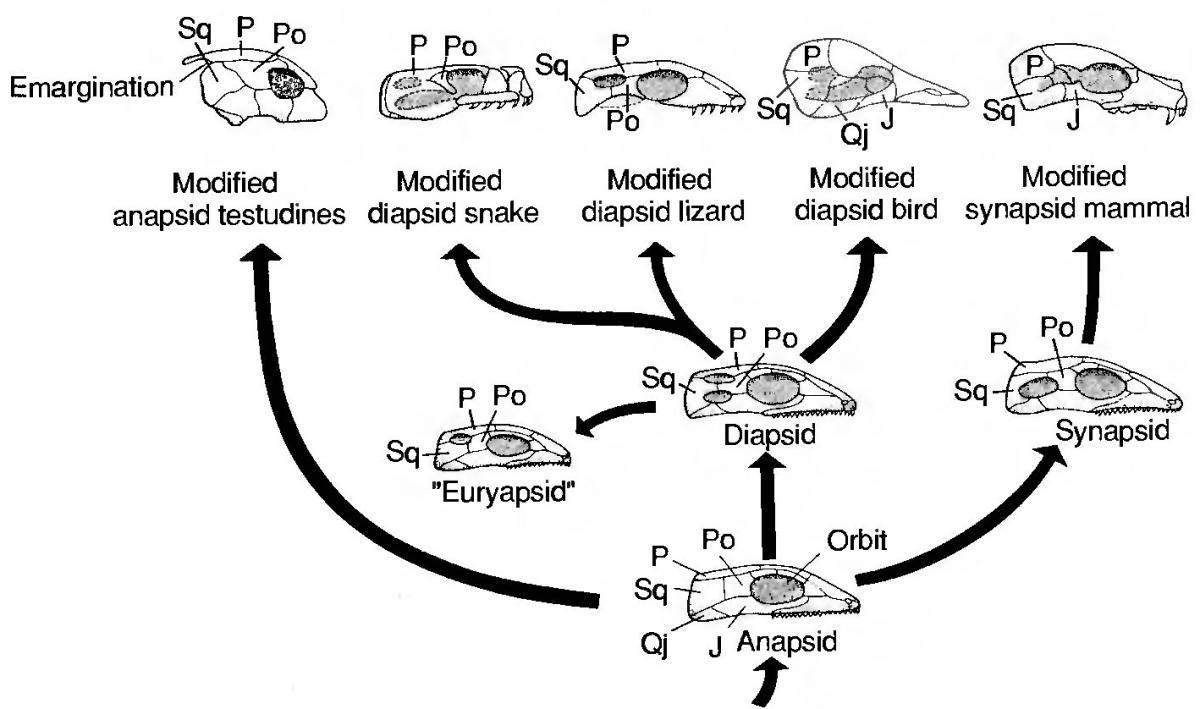
Переважна більшість рептилій належала до груп, у яких утворювалися два скроневих вікна (такий череп має назву діапсидного). Крокодили та гатерії зберегли діапсидний череп; ящірки втратили нижню вилічну дугу, що замикала ніжнє вікно, а змії загалом втратили дуги, що замикали вікна. Птахи, що також належать до групи завропсид, що характеризується типовим або зміненим діапсидним черепом, навпаки, втратили дуги, що закривали скроневі отвори зверху.
Для двох морських груп рептилій, іхтіозаврів та синаптозаврів (плезіозаврів, спільно з іншими близькими до них групами) був характерним евріапсидний череп. Він мав один отвір, але не такий, як у синапсід, а подібний до верхнього отвору діапсид. На підставі цього було прийнято вважати, що евріапсиди є бічною гілкою диапсид.
На підставі описаної тут схеми фенестрації скроневої частини черепу було прийнято розділяти рептилій на підкласи. І тут відкрилися можливості для молекулярної філогенетики.
Яких відносин між черепахами, гатеріями, ящірками (та іншими лускатими) та крокодилами можна було очікувати на підставі таких поглядів? Гатерії, лускаті та крокодили є діапсидами. Вони мали бути відносно близькими. Черепахи та ссавці мали бути філогенетично більше далекими від діапсид, причому набагато.
Як не дивно, черепахи опинилися у гілці між різними діапсидами. Скоріше за все, вони відділилися від спільної гілки з крокодилами вже після того, як розішлися лінії гатерій та лускатих... Як таке пояснити? В усякому разі, будова скроневої ділянки черепа перестала бути надійним дороговказом.
Загальноприйнятої систематики рептилій зараз не існує. Для цілей навчання далі наведено дуже спрощену тимчасову систему. Черепахи в ній поєднані (на підставі палеонтологічних даних) з синаптозаврами. Ще одна "проблемна" група — іхтіозаври — розглядається у складі окремого інфракласа еурептилій.
Як і у інших випадках, ті групи, які мають уявляти собі студенти, виділено цегловим кольором.
Клас "Reptilia" (sensu lato) — Рептилії (у широкому сенсі, включно з синапсидами)
Філа "Synapsida" — Синапсиди
† Ряд Caseasauria — Казеазаври
† Ряд Pelycosauria — Пелікозаври
Ряд "Therapsida" — Терапсиди
† Підряд Biarmosuchia — Біармозухи
† Підряд Dinocephalia — Діноцефали
† Підряд Anomodontia — Аномодонти
† Підряд Gorgonopsia — Горгонопси
† Підряд Therocephalia — Тероцефали
Підряд "Cynodontia" — Цинодонти
Філа Sauropsida — Завропсиди
† Підклас Parareptilia — Парарептилії
† Ряд Pareiasauromorpha — Парейязавроморфи
Підклас "Eureptilia" — Еурептилії (Справжні рептилії)
† Інфраклас Ichthyosaursa — Іхтіозаври
Інфраклас Pantestudines — Пантестудіни
† Ряд Placodontia — Плакодонти
† Ряд Eosauropterygia — Еозавроптерігії
† Ряд Plesiosauria — Плезіозаври
Ряд Testudines — Черепахи
Інфраклас Lepidosauria — Лепідозаври
Ряд Rhynchocephalia — Клювоголові
Ряд Squamata — Лускаті
Інфраклас "Archosauria" — Архозаври
Ряд Crocodylia — Крокодили
† Ряд Pterosauria — Птерозаври
† Підряд Rhamphorhynchoidea — Рамфоринхоідеї
† Підряд Pterodactyloidea — Птеродактилоідеї
Надряд "Dinosauria" — Динозаври
Ряд "Saurischia" — Ящеротазові
Підряд "Theropoda" — Тероподи
† Підряд Sauropodomorpha — Зауроподоморфи
† Ряд Ornithischia — Птахотазові
† Підряд Thyreophora — Тиреофори, Щитоносні
† Інфраряд Stegosauria — Стегозаври
† Інфраряд Ankylosauria — Анкілозаври
† Підряд Cerapoda — Цераподи
† Інфраряд Pachycephalosauria — Пахіцефалозаври
† Інфраряд Ceratopsia — Цератопси
† Інфраряд Ornithopoda — Орнітоподи
Майте на увазі: наведена систематика є дуже спрощеною! Скоріше за все, в ній відсутня чимала кількість невеликих груп, які зіграли величезну роль у еволюції і походженні важливих груп.
В побудові цій систематики використано позначку †, що маркує групи, усі представники і нащадки яких вимерли. Лапки використані для тих груп, що включають не усіх своїх нащадків (тобто є парафілетичними"). Так, у склад "Synapsida" не входять ссавці, а у склад "Eureptilia" — птахи. Зрозуміло, що та група, яку ми позначаємо "Theropoda" (яка, скоріше за все, є предковою для птахів), у тому складі, у якому ми її зазначаємо в цій систематиці, є вимерлою. Втім, деякі нащадки цієї групи, скоріше за все, існують.
Філа "Synapsida" — Синапсиди
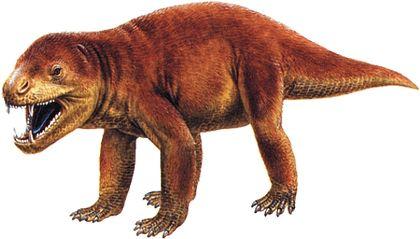
Ми розглянемо три групи синапсидних тварин. Перша, ряд Caseasauria — Казеазаври, є найпримітивнішою. За деякими поглядами, вони не перебувають у тісній спорідненості з іншими двома групами; можливо, в цій групі розвиток єдиного синапсидного вікна, проходив інакше, ніш у інших груп. Представники цього ряду відомі з пізнього карбону та пермі. До казеазаврів, безумовно, належить Cotylorhynchus, якого зображено на наступному рисунку на задньому тлі. Це були пермські, доволі крупні рослиноїдні тварини. Котилоринх сягав 6 м в довжину і був найкрупнішою відомою нам твариною свого часу. Перед котилоринхом розташовані Ophiacodon і Varanops — представники груп, яких частіше включають у наступний ряд, але у деяких випадках зближують з казеазаврами.
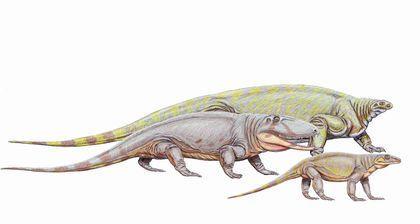
Ophiacodon — це пізнекарбоновий і ранньопермський напівводяний хижак. Найбільші представники роду перевищували в довжину 3 м. У деяких представників роду череп міг бути і діапсідним; у одного примірника череп з одного боку був діапсідним, а з іншого — сінапсідним (!). Така асиметрія може бути свідченням нестійкості в розвитку ознак, які в ході подальшої еволюції групи виявилися набагато краще стабілізованими. Родину Varanopidae, до якої належав Varanops, утворюють невеликі (до 1 м) пермські тварини.

Ряд пелікозаври (Pelycosauria) відомий з пізнього карбону і ранньої перми. До сфенакодонтів (Sphenacodontia) належали м'ясоїдні пелікозаври, а до едафозаврів (Edaphosauria) — рослиноїдні. Для обох груп характерною є наявність крупних гребенів, що утворені остистими відростками хребців.
Найвідоміший представник сфенакодонтів — диметродон (Dimetrodon). Це ранньопермський рід з кількома видами.
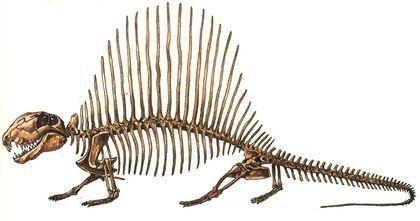

Найкрупніші диметродони сягали 4 м в довжину.
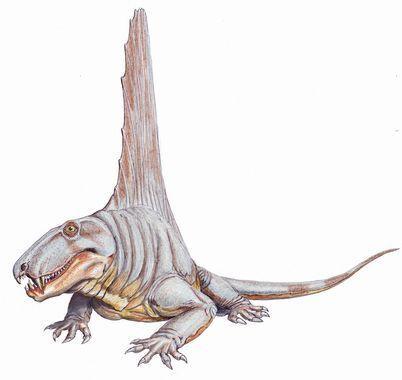
Найбільші едафозаври (Edaphosaurus) перевщували 3 м в довжину. На їхніх гребенях находилися й поперечні відростки, який перетворювали усе "вітрило" в складну тривимірну конструкцію.
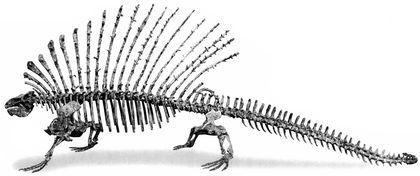

Считается, что именно сфенакодонты были предками последующих тероморф (и, соответственно, нашими предками тоже).
К диноморфам (Dinomorpha) относятся достаточно крупные пермские животные. Горгодонты (Gorgodontia) — плотоядные и растительноядные животные, которых разделяют на два подотряда: Дейноцефалы ("ужасноголовые", Dinocephalia) и Горгонопии (Gorgonopia). У обоих групп развивалась гетеродонтность — подразделение зубов на функциональные группы. У многих дейноцефалов это подразделение было иным, чем у млекопитающих. Это обстоятельство, а также развитие утолщение на черепе и обусловило их название.
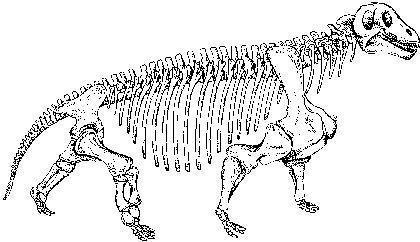
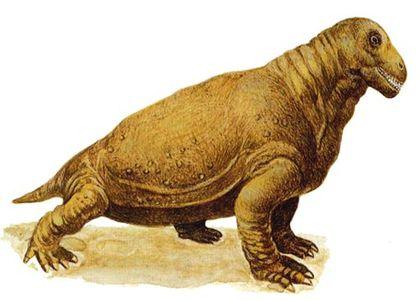
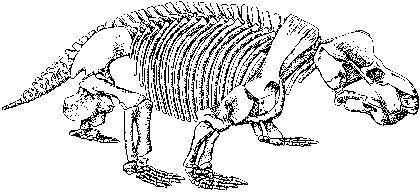
Примером плотоядных дейноцефалов может быть титанофонеус ("титанический убийца", Titanophoneus), скелет и череп которого показаны на двух следующих слайдах, а реконструкция - еще на следующем. Это животное достигало нескольких метров в длину.



Некоторые дейноцефалы были растительноядными. Например, это относится к тапиноцефалидам (Tapinocephalida). Эти крупные животные пережевывали растительную пищу передними зубами, отжимая ее языком кпереди от хоан. На лбу тапиноцефалов находилась губчатая костная ткань. Раньше предполагали, что тапиноцефалы могли бодаться, сейчас это подвергают сомнению (пахиостоз, разрастание кости, был у них достаточно хрупким).

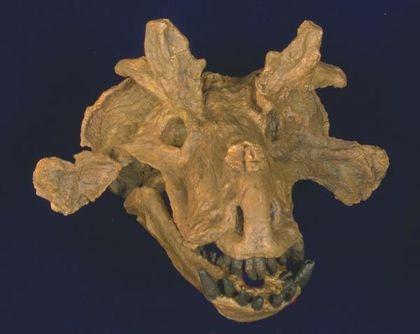
Также к дейноцефалам относят эстемменозухов (Estemmenosuchus) — крупных животных с потрясающим воображение черепом.
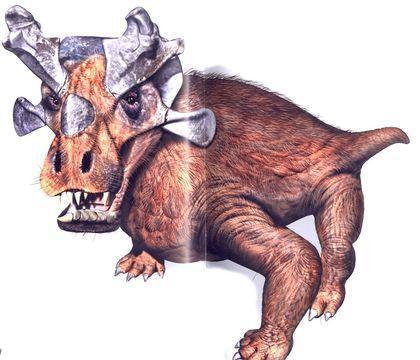

Для горгонопий свойственно такое же подразделение зубов на функциональные группы, какое было унаследовано млекопитающими. Жевание у них не могло быть эффективно (из-за отсутствия вторичного костного неба). Одна из известнейших горгонопий — иностранцевия (Inostrancevia), серьезный хищник, достигавший нескольких метров в длину.

Вероятно, иностранцевия была полуводной и могла охотиться на парейязавров (представителей парарептилий).

Кожа иностранцевии не была сухой и, вероятно, несла волосы, выполнявшие осязательные функции.
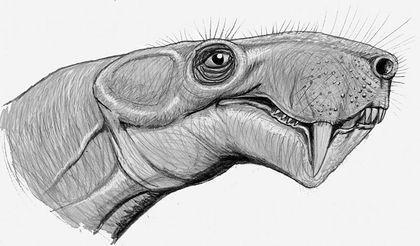
Все диноморфы были пермскими животными, а вот представители следующего отряда — аномодонты (Anomodontia) перешли из перми в триас. Аномодонты были чрезвычайно разнообразной группой. Самыми успешлыми из них оказались Дицинодонты (Dicynodontia), которые дожили до конца триаса. Нижняя челюсть дицинодонтов при закрывании рта "отъезжала" назад, что позволяло откусывать пищу (прежде всего — растительную). Многие дицинодонты теряли почти все или все зубы, и использовали для срывания пищи роговой клюв на верхней челюсти.
 (кликабельно)
(кликабельно)
Черепа дицинодонтов могут быть очень выразительными.
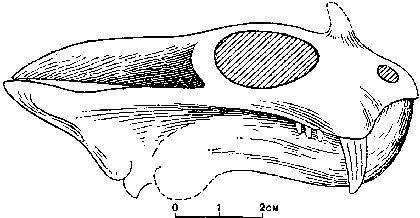
Наконец, к инфраклассу Терозавры (буквально — "Звероящеры", Therosauria) принадлежат отряды Тероцефалы ("Звероголовые", Therocephalia) и Цинодонты ("Собакозубые", Cynodontia). Эти существа имели вторичное костное небо и по многим признакам напоминали млекопитающих.
Говоря о тероцефалах, нельзя не упомянуть замечательный пример супружеской любви, проявленный Владимиром Прохоровичем Амалицким. Амалицкий был крупнейшим исследователем пермских позвоночных конца XIX-начала XX вв; например, именно он открыл иностранцевию, которую назвал в честь своего учителя А.А. Иностранцева. В честь своей жены он назвал намного более привлекательного тероцефала.

Тероцефалы — разнообразная группа пермских "рептилий", многие из которых были некрупными ночными животными. Их обоснованно реконструируют покрытыми волосами.
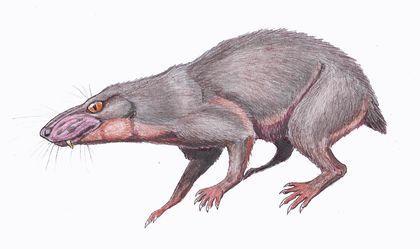
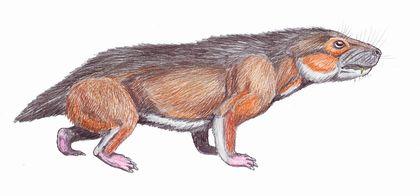
Наконец, цинодонты — группа, представителей которой, вероятно, было бы сложно отличить от млекопитающих. Их зубы не просто дифференцированы; они приобретают сложную форму, увеличивавшую эффективность их использвания.
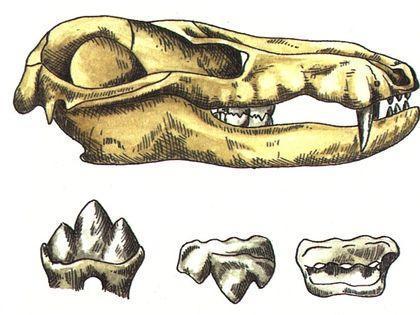
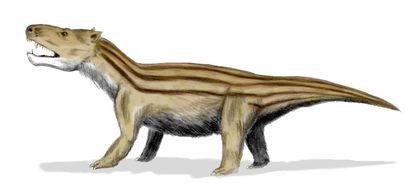
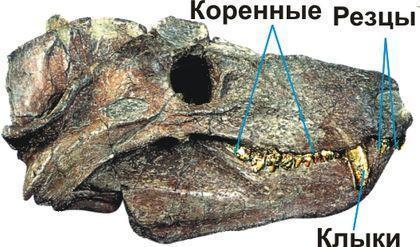
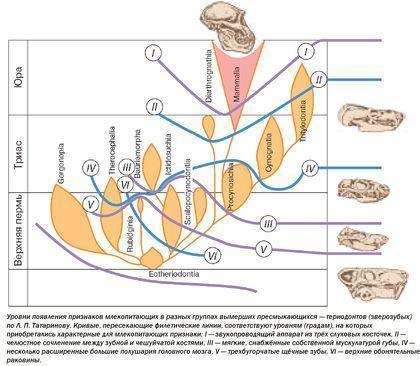
Важным выводом, который сделан на основании эволюции тероморфов, является то, что с течением времени в каждой из их ветвей возрастала доля "маммальных" признаков , характерных для млекопитающих.
Коментарі
Очепятки
1. Черепа дицинодонтов могуТ быть.
2. отряды Тероцефалы ("Звероголовые", Therocephalia) и [Ц]инодонты ("Собакозубые", Cynodontia).
3. Говоря о тероцефалах, нельзя не упомянуть замечательный пример супружеской любви...К примеру, Амалицкий открыл иностранцевию, которую назвал в честь своего учителя А.А. Иностранцева.
cмешались в кучу Annatherapsidus, Inostrancevia.
Спасибо!
Чуть разделил иностранцевию и Анну Петровну.
Так гораздо логичнее, но
иМенно он открыл.