Усова Е. Е. Определение естественной смертности половозрелых зеленых лягушек (Pelophylax esculentus complex; Amphibia, Ranidae) с использованием скелетохронологии // Вісник Харківського національного університету імені В.Н.Каразіна. Серія: біологія. – 2010. – Вип.12 (№920). – С. 104–110.
УДК (57.022+57.032):591.84:591.158.1:597.851
ОПРЕДЕЛЕНИЕ ЕСТЕСТВЕННОЙ СМЕРТНОСТИ ПОЛОВОЗРЕЛЫХ ЗЕЛЕНЫХ ЛЯГУШЕК (PELOPHYLAX ESCULENTUS COMPLEX; AMPHIBIA, RANIDAE) С ИСПОЛЬЗОВАНИЕМ СКЕЛЕТОХРОНОЛОГИИ
Е.Е.Усова
Биологический ф-т Харьковского национального университета имени В.Н.Каразина (Харьков, Украина)
Изучен характер смертности зеленых лягушек (Pelophylax esculentus и Pelophylax ridibundus) из Харьковской области по скелетохронологическим данным (146 особей). На основании распределения по возрасту половозрелых лягушек, пойманных на местах нереста, сделано предположение, что большинство самок нерестится впервыев возрасте 4-х, а самцов — в возрасте 5-и лет. Для половозрелых зеленых лягушек характерен II тип кривой выживания по Р. Перлю(смертность не зависит от возраста).
Ключевые слова: Pelophylax esculentus(=Ranaesculenta)complex, скелетохронология, половозрелость, смертность.
О. Є. Усова. Визначення природної смертності статевозрілих зелених жаб (Pelophylax esculentus complex; Amphibia, Ranidae) із застосуванням скелетохронології. Вивчено характер смертності зелених жаб (Pelophylax esculentusі Pelophylaxridibundus) з Харківської області за скелетохронологічними даними про вік 146 особин. На підставі розподілу за віком жаб, спійманих на місцях нересту, зроблене припущення, що більшість самок нереститься вперше у віці 4+, а самців —у віці 5+ років. Для статевозрілих зелених жаб характерний II тип кривої виживання за Р. Перлем (смертність не залежить від віку).
Ключові слова: Pelophylax esculentus (=Ranae sculenta) complex, скелетохронологія, статевозрілість, смертність.
E. Е. Usova. Determining the level of mature water frogs (Pelophylax esculentus complex;Amphibia, Ranidae) natural mortality by use of skeletochronology.The rate of mortality of water frogs (Pelophylax esculentus and Pelophylax ridibundus) from the Kharkiv region was estimated on the basis of skeletochronology data on the age of 146 individuals. Taking into account the age distribution of the frogs caught in the spawning grounds, we suggest that females come on the first spawning by age 4+, and males – by age 5+. R. Pearl type II survival curve is characteristic for adult water frogs.
Keywords: Pelophylax esculentus (=Rana esculenta) complex, skeletochronology, maturity, mortality.
Введение
В гибридогенный комплекс зеленых лягушек, Pelophylax esculentus complex (=Rana esculenta complex), входят два родительских вида, Pelophylax lessonae (Camerano, 1882) и Pelophylax ridibundus (Pallas, 1771), а также различные ди‑, три‑ и тетраплоидные формы их гибридов (Plötner, 2005), которые в данной работе рассматриваются под именем Pelophylax esculentus (Linnaeus, 1758). Совместное обитание гибридов с родительскими видами поддерживается благодаря гемиклональному наследованию (передаче гибридами определенных геномов клонально, без рекомбинации), что представляет значительный теоретический интерес. Одни из наиболее сложных гемиклональных популяций зеленых лягушек зарегистрированы в бассейне верхнего течения р. Северский Донец в пределах Харьковской области (Шабанов, Литвинчук, 2010). Для определения механизмов поддержания столь сложных биосистем следует изучить протекающие в них демографические процессы.
Задача данной работы — определить присущий половозрелым лягушкам характер естественной смертности, используя результаты скелетохронологического определения возраста.
Изученный материал
Нами определен возраст 163 половозрелых лягушек, 59 самок и 104 самцов (табл. 1). Отлов лягушек проводился в Харьковской области с 2004 по 2009 г. Почти половина из них (73) собрана в окрестностях биостанции ХНУ (с. Гайдары Змиевского района Харьковской области) и Национального природного парка «Гомольшанские леса». Видовая принадлежность и состав геномов в генотипе для большинства изученных особей определялась методом проточной ДНК-цитометрии в ИНЦ РАН (Санкт-Петербург) С. Н. Литвинчуком и Ю. М. Розановым. Часть особей (представители P. lessonae) определены морфологически по комплексу признаков (Лада, 1995; Шабанов и др., 2006).
Таблица 1. Состав изученной выборки лягушек (L — геном P. lessonae; R — геном P. ridibundus)
|
Генотип |
Самок |
Самцов |
Из них из окрестностей биостанции ХНУ и НПП «Гомольшанские леса» |
|
LL |
8 |
9 |
- |
|
LLR |
1 |
– |
- |
|
LR |
18 |
73 |
47 |
|
LRR |
5 |
8 |
11 |
|
RR |
27 |
14 |
15 |
Методика скелетохронологического определения возраста
Для определения продолжительности жизни мы использовали метод скелетохронологии (Смирина, 1983 и др. работы; Усова 2010). Третью фалангу длиннейшего пальца задней ноги лягушек очищали от мягких тканей, декальцинировали, получали на замораживающем микротоме срезы толщиной 20-22 мкм средней части диафиза, окрашивали гематоксилином Эрлиха, фотографировали временные препараты, а затем проводили измерения и подсчеты линий склеивания по фотографиям. Эти линии образуются при зимовках; их подсчет позволяет определить возраст лягушек. Поскольку определенная часть онтогенеза лягушки проходит до ее первой зимовки, и, кроме того, определение возраста, как правило, проводится у лягушек, вышедших после зимовки, возраст лягушки, имеющей, к примеру, 3 линии склеивания, обозначается как «3+», т.е. «более 3-х лет».
Одна из главных сложностей при определении возраста связана с тем, что при добавлением новых линий склеивания с наружной, периостальной части кости происходит их разрушение (резорбция) изнутри, со стороны эндостальной полости. В ряде случаев периостальные кольца разрушаются не полностью, и о том, что они когда-то существовали, свидетельствуют их сохранившиеся фрагменты. Но даже в случае полного разрушения одной или нескольких линий можно с достаточной надежностью оценить их количество. Диаметр первых линий склеивания у разных особей отличается в меньшей степени, чем диаметр последующих линий; хотя на протяжении всей жизни отличия в размере тела и диаметре линий склеивания в фалангах у быстрорастущих и медленнорастущих лягушек увеличиваются, размеры молодых лягушек и диаметр их линий склеивания оказываются достаточно близкими. При изучении срезов фаланг пальцев неполовозрелых лягушек мы определили типичные размеры линий склеивания, образующихся на 1-м и 2-м году жизни. Измерив диаметр эндоста, можно определить, сколько линий склеивания должно было помещаться в занятой им полости. У большинства половозрелых лягушек резорбировались 2 линии склеивания, у некоторых особей — 1 или 3. В некоторых случаях точное определение возраста оказывается невозможным. В таких случаях мы указывали наиболее вероятный возраст, допуская, что действительный возраст может быть на один год меньше или больше.
Особенности статистической обработки зарегистрированных возрастных распределений
Результаты определения возраста у изученных особей показаны в табл. 2. Как видно, каждая из категорий лягушек (например, особи определенного пола, заданного генотипа, пойманный в конкретном местообитании в тот или иной год) представлена небольшим количеством особей, недостаточным для обоснованного суждения о характере их смертности. В то же время вся совокупность данных достаточна для ее анализа статистическими методами. Если распределения по возрасту, полученные для разных категорий особей, не имеют существенных отличий, их можно рассматривать совместно.
Таблица 2. Распределение по возрасту изученных особей лягушек
|
Генотип |
Возраст самок |
Возраст самцов |
|||||||||||
|
3+ |
4+ |
5+ |
6+ |
7+ |
8+ |
9+ |
10+ |
3+ |
4+ |
5+ |
6+ |
7+ |
|
|
LL |
– |
3 |
3 |
1 |
1 |
– |
– |
– |
3 |
5 |
1 |
– |
– |
|
LLR |
– |
– |
– |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
LR |
5 |
7 |
2 |
1 |
1 |
– |
1 |
1 |
8 |
13 |
29 |
16 |
6 |
|
LRR |
– |
2 |
1 |
2 |
– |
– |
– |
– |
– |
– |
4 |
4 |
– |
|
RR |
3 |
10 |
7 |
3 |
2 |
1 |
1 |
– |
– |
1 |
6 |
4 |
1 |
Прежде всего, для дальнейшего анализа следует определить, необходимо ли обрабатывать полученные данные по отдельности для каждого пола, или их можно обрабатывать всех вместе. Мы сравнили (с помощью программы Statistica for Windows 6.5) зарегистрированное распределение возрастов самок и самцов с ожидаемым, вычисленным при условии отсутствия связи между полом и возрастной структурой (табл. 3).
Таблица 3. Пример сравнения эмпирического и ожидаемого (основанного на предположении об отсутствии связи между признаками, отраженными в строках и столбцах таблицы) распределений. Показаны возрастные распределения самок и самцов лягушек. χ2= 16,07; к-во степеней свободы =7; p=0,0245; различие распределений значимо.
|
Распределения |
Пол |
Возраст |
Всего |
|||||||
|
3+ |
4+ |
5+ |
6+ |
7+ |
8+ |
9+ |
10+ |
|||
|
Эмпирическое |
Самки |
8 |
22 |
13 |
8 |
4 |
1 |
2 |
1 |
59 |
|
Самцы |
11 |
19 |
40 |
24 |
7 |
3 |
0 |
0 |
104 |
|
|
Всего |
19 |
41 |
53 |
32 |
11 |
4 |
2 |
1 |
163 |
|
|
Ожидаемое |
Самки |
6,88 |
14,84 |
19,18 |
11,58 |
3,98 |
1,45 |
0,72 |
0,36 |
59 |
|
Самцы |
12,12 |
26,16 |
33,82 |
20,42 |
7,02 |
2,55 |
1,28 |
0,64 |
104 |
|
|
Всего |
19 |
41 |
53 |
32 |
11 |
4 |
2 |
1 |
163 |
|
Сравнение эмпирического и ожидаемого распределения по критерию Пирсона χ2 позволяет оценить значимость наблюдаемого отклонения распределения от случайного. Как ясно из табл. 3, возрастные распределения самок и самцов значимо отличаются. Это означает, что данные по двум полам следует обрабатывать по отдельности.
Следующим шагом необходимо определить, можно ли совместно рассматривать данные по отдельным генотипам. Для самок разница между эмпирическим и ожидаемым распределением недостоверна (р=0,8). Для самцов такая разница значима (р=0,007), причем в первую очередь (см. табл. 2) она связана с отличием между распределением возрастов самцов P. lessonae и представителей других генотипов. При исключении из рассмотрения самцов P. lessonae разница между зарегистрированным и ожидаемым распределением для оставшихся генотипов оказывается незначимой (р=0,69). И это означает, что данные о распределении по возрасту P. ridibundus и различных представителей P. esculentus можно рассматривать совместно, а P. lessonae характеризуются иным распределением, по крайней мере, для самцов.
Далее необходимо определить, можно ли совместно рассматривать данные о распределении по возрасту, полученные от лягушек из разных популяций. Как показано в табл. 1, почти половина изученных особей (и более половины от количества изученных особей P. ridibundus и P. esculentus) получена из одной метапопуляции, населяющей окрестности биостанции ХНУ и НПП «Гомольшанские леса». Сравнивая распределение по возрасту особей из этой метапопуляции, а также остальных изученных нами представителей P. ridibundus и P. esculentus из Харьковской области, мы можем убедиться, что связь между возрастом и происхождением лягушек незначима (р=0,43 для самок и р=0,29 для самцов). Сравнение распределений лягушек, пойманных в разные годы, также дает основания для их совместного рассмотрения.
Следовательно, все полученные данные, относящиеся к P. ridibundus и P. esculentus из Харьковской области, можно рассматривать совместно. Безусловно, совместное рассмотрение данных для лягушек разных форм, пойманных в разных местообитаниях и в разное время, является упрощением действительной картины. Такое упрощение не является грубым искажением, но зато оно дает возможность оценить общий характер возрастного распределения.
Результаты и обсуждение: характер кривых выживания для половозрелых лягушек
Возрастные распределения для всех изученных форм зеленых лягушек, за исключением P. lessonae, показаны на рис. 1.
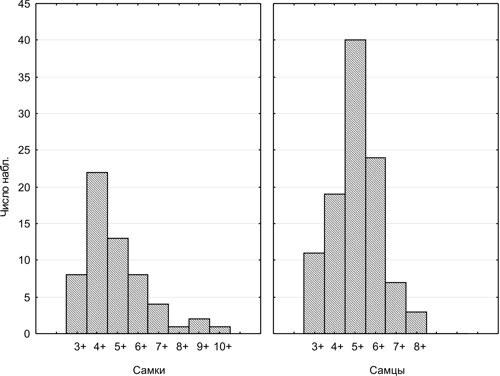
Рис. 1. Возрастное распределение, зарегистрированное для самок и самцов P. ridibundus и P. esculentus из Харьковской области (данные для разных генетических форм лягушек, разных местообитаний и разных лет сбора рассматриваются совместно)
При анализе данных, представленных на рис. 1, заслуживает внимание то, что для самок самым многочисленным возрастным классом оказывается 4+, а для самцов — 5+. Вероятно, это свидетельствует о том, что именно к такому возрасту большинство представителей указанных полов приходит на нерест в первый раз. К примеру, нет оснований предполагать, что в популяции в целом самок возраста 4+ больше, чем возраста 3+. Преобладание старших возрастных классов в изученной выборке можно объяснить тем, что она в первую очередь состоит из особей, пойманных на нересте. Вероятно, значительная часть трехлеток не приходит на нерестилище, поскольку не достигает половозрелости. Начиная от четырехлетнего возраста самки год от года регулярно приходят на нерест, и сокращение их численности отражает их естественную смертность. Поскольку нерестовые продукты образуются у бесхвостых осенью, приход на нерест в возрасте 4+ означает, что половозрелость была достигнута в возрасте 3+.
Аналогично, можно предположить, что первый нерест для большинства самцов происходит в возрасте 5+. Значит, из-за отсутствия полных данных об относительной численности более младших возрастных групп, смертность самок по нашим результатам можно анализировать начиная с 4-х летнего возраста, а самцов — с 5-летнего.
Для интерпретации полученных результатов можно воспользоваться классификацией типов кривых выживания, принадлежащей американскому классику популяционной экологии Раймонду Перлю (Pearl, 1937). Преимущественная смертность в зрелом возрасте (рис. 2) характерна для популяций с I типом кривых выживания по Перлю, например, для людей в цивилизованном обществе. Этот тип кривых называют типом дрозофилы, имея в виду имаго дрозофил в лабораторной культуре. III тип, тип устрицы, со смертностью в раннем возрасте, характерен для многих животных с высокой плодовитостью. Не зависящая от возраста смертность на графике с логарифмической шкалой численности характерна для II типа кривых выживания, типа гидры. Этот тип кривых выживания регистрируется у организмов, уже прошедших самую уязвимую стадию своего онтогенеза (Шабанов, Кравченко, 2009).
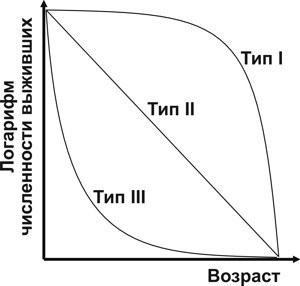
Рис. 2. Типы кривых выживания по Р. Перлю (источник иллюстрации: Шабанов, Кравченко, 2009)
Чтобы определить тип смертности, характерный для половозрелых лягушек, данные о численности их возрастных классов следует перевести в логарифмическую форму (табл. 4).
Таблица 4. Итоговые данные для построения кривых выживания. В двух последних столбцах указаны только точки, использованные для построения кривых
|
Возраст |
Численность самок (FemalesN) |
Численность самцов (MalesN) |
Логарифм численности самок Log2(FemalesN) |
Логарифм численности самцов Log2(MalesN) |
|
3+ |
8 |
11 |
|
|
|
4+ |
22 |
19 |
4,46 |
|
|
5+ |
13 |
40 |
3,70 |
5,32 |
|
6+ |
8 |
24 |
3,00 |
4,58 |
|
7+ |
4 |
7 |
2,00 |
2,81 |
|
8+ |
1 |
3 |
0,00 |
1,58 |
|
9+ |
2 |
0 |
1,00 |
|
|
10+ |
1 |
0 |
0,00 |
|
Кривые выживания, построенные на основании приведенных в табл. 4 данных, показаны на рис. 3.
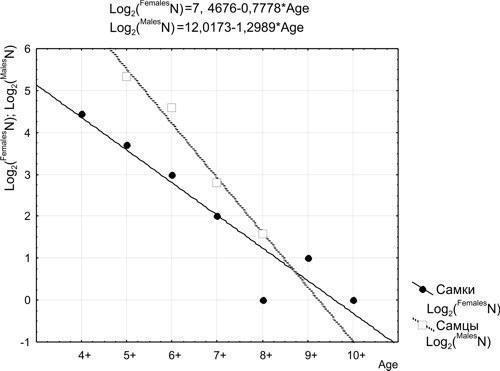
Рис. 3. Кривые выживания для самок и самцов зеленых лягушек
Как видно из рис. 3, кривые выживания для половозрелых особей обоих полов соответствуют II типу по Перлю (за исключением «вылетающей» точки, соответствующей восьмилетним самкам). В верхней части графика приведены уравнения линий регрессии, аппроксимирующих зарегистрированное распределение возрастов.
В полученных уравнениях регрессии первое слагаемое определяет уровень, на котором проходит кривая (численность первого из рассматриваемых возрастных классов, т.е. фактически — численность изученной выборки), а множитель во втором слагаемом определяет наклон кривой (уровень ежегодной смертности). В логарифмическом масштабе численность каждого следующего возрастного класса меньше на определенную величину. Обозначив как Log2(FemalesNA) двоичный логарифм численности самок в возрасте A (при условии A>4), и, соответственно, Log2(FemalesNA+1) — двоичный логарифм их численности в возрасте A+1, мы на основании показанных на рис. 3 данных можем утверждать, что Log2(FemalesNA+1) = Log2(FemalesNA) – 0,7778. Отсюда FemalesNA+1 = FemalesNA × 2–0,7778 и, наконец, FemalesNA+1 = 0,58 × FemalesNA.
Аналогично, Log2(MalesNA+1) = Log2(MalesNA) – 1,2989, откуда MalesNA+1 = MalesNA × 2–1,2989 и, окончательно, N(Male)A+1 = 0,406 × MalesNA.
Упрощая, можно сказать, что из 10 половозрелых самок до следующего года доживут 6, а из 10 половозрелых самцов — 4.
Следует подчеркнуть, что независимая от возраста смертность лягушек характерна не для всего их онтогенеза, а только для его последних стадий.
Полученный результат относится к обобщенной выборке, содержащей P. ridibundus и P. esculentus. К сожалению, имеющихся в нашем распоряжении данных о возрастной структуре P. lessonae недостаточно для анализа структуры их смертности.
Несмотря на то, что полученные кривые носят усредненный характер, нивелируя разницу между местообитаниями, различными лягушек и годами, они являются первой оценкой смертности зеленых лягушек в естественных местообитаниях. Такая оценка, кроме прочего, необходима для математического моделирования процессов в гемиклональных популяционных системах зеленых лягушек (Кравченко, 2010). Дальнейшие исследования могут уточнить выясненные закономерности и показать специфику демографических процессов, характерных для отдельных форм этих животных.
Благодарности
Автор выражает искреннюю благодарность С. Н. Литвинчуку, Ю. М. Розанову и А. В. Коршунову за помощь в идентификации исследуемого материала, а также Д. А. Шабанову за участие в интерпретации и статистической обработке результатов.
Работа выполнена при поддержке Фонда фундаментальных исследований ХНУ им. В. Н. Каразина и совместного гранта ГФФИ Украины и РФФИ (Россия).
Список литературы
Кравченко М. А. Математическое моделирование динамики различных кариогенетических форм зеленых лягушек в гемиклональных популяционных системах // Мат. ХI Междунар. научно-практич. экологической конф. — Белгород, 2010. —С. 210.
Лада Г. А. Среднеевропейские зеленые лягушки (гибридогенный комплекс Rana esculenta): введение в проблему // Флора и фауна Черноземья. — Тамбов, 1995. — С. 88–109.
Смирина Э. М. Прижизненное определение возраста и ретроспективная оценка размеров тела серой жабы (Bufo bufo) // Зоол. журн. — 1983. — т. 63, № 3. — С. 437–444.
Усова Е. Е. Возрастная структура представителей гибридогенного комплекса зеленых лягушек в Харьковской области // Мат. ХI Междунар. научно-практич. экологической конф. — Белгород, 2010. — С. 131.
Шабанов Д. А., Зиненко А. И., Коршунов А. В., Кравченко М. А., Мазепа Г. А. Изучение популяционных систем зеленых лягушек (Rana esculenta complex) в Харьковской области: история, современное состояние и перспективы // Вісник Харківського національного університету імені В.Н.Каразіна. Серія: біологія. — 2006. — Випуск 3 (№ 729). — С. 208–220.
Шабанов Д. А., Кравченко М. А. Материалы для изучения курса общей экологии с основами средоведения и экологии человека. — Х.: ХНУ имени В.Н. Каразина, 2009. — 292 с.
Шабанов Д. А., Литвинчук С. Н. Зеленые лягушки: жизнь без правил или особый способ эволюции? // Природа. —2010. — № 3. — С. 29–36.
Pearl R. On biological principles affecting populations: human and other // Amer. Natur. — 1937. — Vol. 71, N 1. — P. 50-68.
Plötner J. Die westpaläarktichen Wasserfrösche. Bielefeld: Laurenti-Verlag, 2005. – 161 S.
Коментарі
Можно ли на основании данной публикации
использовать в работе по оценке численности ГПС зелёных лягушек на Иськовом выражение не "половозрелые самцы и полувзрослые особи", а "самцы, старше 3(4) лет"? Или наши были ещё моложе?
Нужно говорить о том, что мы регистрировали
Если мы определяли фазу онтогенеза, нужно называть фазу онтогенеза (например, полувзрослые самцы). Если мы определяли возраст, нужно указывать возраст. Но я не помню, чтоб мы определяли возраст во время учета!
Раз
не брали сеголетков, значит минимум - двухлетние. По-моему это лучше воспринимается, чем "самцы и полувзрослые особи", как Вы помните, об этом непосредственно на защите встал вопрос. Хотя это, в принципе, детали.
В принципе, если на защите кто-то другой спросит, в чём различие между полувзрослыми особями и самцами, теперь можно будет назвать конкретные количества лет жизни.