Систематический список земноводных Украины
Класс Amphibia
Отряд бесхвостые — Anura
Семейство квакши — Hylidae
Род Квакша — Hyla
Квакша обыкновенная — Hyla arborea
Квакша восточная — Hyla orientalis
Семейство жерлянки — Bombinatoridae
Род Жерлянка — Bombina
Краснобрюхая жерлянка — Bombina bombina
Желтобрюхая жерлянка — Bombina variegata
Семейство чесночницы — Pelobatidae
Род Чесночница — Pelobates
Обыкновенная чесночница — Pelobates fuscus
Чесночница Палласа — Pelobates vespertinus
Семейство жабы — Bufonidae
Род Жаба — Bufo
Серая или обыкновенная жаба — Bufo bufo
Зеленая жаба — Bufo viridis
Жаба камышовая — Bufo calamita
Семейство лягушки — Ranidae
Род Бурая лягушка — Rana
Остромордая лягушка — Rana arvalis
Прыткая лягушка — Rana dalmatina
Травяная лягушка — Rana temporaria
Род Зеленая (водная) лягушка — Pelophylax
Озерная лягушка — Pelophylax ridibundus
Съедобная лягушка — Pelophylax esculentus (гемиклональный межвидовой гибрид)
Прудовая лягушка — Pelophylax lessonae
Отряд хвостатые — Caudata
Семейство саламандровые — Salamandridae
Подсемейство Salamandrinae
Род Саламандра — Salamandra
Пятнистая саламандра — Salamandra salamandra
Подсемейство Pleurodelinae
Род Малый (гладкий) тритон — Lissotriton
Карпатский тритон — Lissotriton montandoni
Обыкновенный тритон — Lissotriton vulgaris
Род Средний тритон — Mesotriton
Альпийский тритон — Mesotriton alpestris
Род Крупный тритон — Triturus
Тритон гребенчатый — Triturus cristatus
Дунайский тритон — Triturus dobrogicus
Тритон Карелина — Triturus karelinii
Источники:
- Писанец Е. М. Амфибии Украины (справочник - определитель земноводных Украины и сопредельных территорий).- Киев: Зоологический музей ННПМ НАН Украины, 2007. — 312 с.
Отряд: Бесхвостые — Anura
Семейство: Квакши — Hylidae
Род: Квакша — Hyla
Квакша обыкновенная — Hyla arborea (Linnaeus, 1758)
 Описание. Животные небольших размеров, длина тела половозрелых особей обычно до 50–60 мм. Пальцы задних конечностей соединены перепонкой. Кожа на спине гладкая, на брюшной стороне крупнозернистая. В спинной окраске чаще всего преобладают однотонные ярко зеленые цвета, хотя встречаются светло серые, коричневатые или серо голубоватые особи. Брюшная сторона белого цвета. Окраска верхней (спинной) и нижней (брюшной) сторон тела разделена двумя контрастными темными полосами с желтобелыми краями, каждая из которых тянется по обоим бокам тела от глаз до паховой области, образуя здесь петлю. Темные полосы есть также и на морде, где они соединяют ноздри и верхне передние края глаз.
Описание. Животные небольших размеров, длина тела половозрелых особей обычно до 50–60 мм. Пальцы задних конечностей соединены перепонкой. Кожа на спине гладкая, на брюшной стороне крупнозернистая. В спинной окраске чаще всего преобладают однотонные ярко зеленые цвета, хотя встречаются светло серые, коричневатые или серо голубоватые особи. Брюшная сторона белого цвета. Окраска верхней (спинной) и нижней (брюшной) сторон тела разделена двумя контрастными темными полосами с желтобелыми краями, каждая из которых тянется по обоим бокам тела от глаз до паховой области, образуя здесь петлю. Темные полосы есть также и на морде, где они соединяют ноздри и верхне передние края глаз.
Распространение. Обитают на большей части Центральной и Западной Европы (отсутствуют в южной Испании и южной Франции), на север граница доходит до Англии (здесь интродуцирована), северо западной части Нидерландов, Норвегии. Квакши обитают на большей части Украины . поднимаясь в горы на высоту до 1250 м; в юго восточном направлении доходят примерно до востока Харьковской, Днепропетровской (долины рек Орель и Самара — Булахов, 2001; Мисюра, Марченковская, 2001) и севера Запорожской областей (есть на о. Хортица — Петроченко, 1990). В южной и юго западной Украине встречается спорадически, начиная от низовий Днепра (Херсонская обл.) и далее на запад; здесь в Одесской области доходит до устья Дуная (в южных регионах степной зоны распространение связано с островными лесными и плавневыми участками). В Крыму встречается в горной части.
Образ жизни. Большинство квакш ведет древесный образ жизни, и это обусловило особое строение конечностей: фаланги пальцев на концах имеют диски – присоски. Половой диморфизм выражен слабо: у самцов имеется не парный горловой резонатор и брачные мозоли на первом пальце передних конечностей. Поющий самец раздувает крупный горловой резонатор.
Днем обычно сидят неподвижно на растениях, охотятся по ночам. Летом можно встретить в влажных местах в пойменном лесу или в нагорной дубраве, в зарослях кустарников в сырых местах. Весной (в апреле-мае) нерест происходит почти во всех непроточных водоемах. Это могут быть мелкие водоемы на просеках или опушках леса, лужи, болотца, мелиоративные канавы, мелководная прибрежная часть озер. В реках и других проточных водоемах квакши икру не откладывают. Интенсивные ночные концерты, устраиваемые самцами, могут продолжаться до конца мая.
Нерест происходит при температуре воды 13°С. Самка откладывает около 690—1870 икринок несколькими порциями в виде небольших комочков (в Молдавии 15—21 комочек по 21—56 икринок). Кладки лежат на дне водоема или прикреплены к растениям. Период икрометания растянут и длится с начала апреля до конца июля, на Кавказе с конца марта до мая. Диаметр икринки с оболочкой 3—4,5 мм, яйцеклетки 1,0—1,6 мм. Эмбриональное развитие длится около 8—14 суток. Размер личинок после выклева 8—9 мм. Личиночное развитие продолжается 45—90 суток. Перед метаморфозом головастики достигают длины 46—49 мм. В Карпатах отмечены случаи зимовки личинок. Сеголетки длиной 10—17 мм и более выходят на сушу в дневное время в июле — начале сентября. В отличие от взрослых они весьма активны днем и держатся в основном на траве у водоемов.
Источники:
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
- Банников А. Г., Даревский И. С., Ищенко В. Г., Рустамов А. К., Щербак Н. Н. Определитель земноводных и пресмыкающихся фауны СССР. — М.: "Просвещение",
Отряд: Бесхвостые — Anura
Семейство: Квакши — Hylidae
Род: Квакша — Hyla
Квакша восточная — Hyla orientalis Bedriaga, 1890
 Систематическое положение. Hyla orientalis имеет высокое генетическое разнообразие в отличие от H. arborea – генетически однообразного вида. Распознается пять географических групп по мтДНК: на Кавказе и прилежащих территориях; в Малой Азии; на западном побережье Черного моря; в Крыму, на северо-западном побережье Черного моря, Украине (кроме Закарпатья), Беларуси, России и Польше Морфологическая изменчивость высока (в частности, у ряда особей из Закавказья нет паховой петли). Между Hyla orientalis и H. arborea, найдены зоны контакта в северо-восточной Греции и в Польше. Крупные реки Карпат дают редкие возможности для вторичных контактов этих видов (в одной точке найдены обе мтДНК), но вторичный контакт и гибридизация документированы лишь на равнине р. Висла. Эти данные свидетельствуют в пользу того, что H. arborea и H. orientalis – разные виды. Кариотип Кариология. 2n=24, NF=48, средний размер генома определен как 10,31–12,17 пг.
Систематическое положение. Hyla orientalis имеет высокое генетическое разнообразие в отличие от H. arborea – генетически однообразного вида. Распознается пять географических групп по мтДНК: на Кавказе и прилежащих территориях; в Малой Азии; на западном побережье Черного моря; в Крыму, на северо-западном побережье Черного моря, Украине (кроме Закарпатья), Беларуси, России и Польше Морфологическая изменчивость высока (в частности, у ряда особей из Закавказья нет паховой петли). Между Hyla orientalis и H. arborea, найдены зоны контакта в северо-восточной Греции и в Польше. Крупные реки Карпат дают редкие возможности для вторичных контактов этих видов (в одной точке найдены обе мтДНК), но вторичный контакт и гибридизация документированы лишь на равнине р. Висла. Эти данные свидетельствуют в пользу того, что H. arborea и H. orientalis – разные виды. Кариотип Кариология. 2n=24, NF=48, средний размер генома определен как 10,31–12,17 пг.
Hyla orientalis, в отличие от H. arborea, генетически весьма гетерогенна. Распознается пять географических групп по мтДНК: на Кавказе и прилежащих территориях; в Малой Азии; на западном побережье Черного моря; в Крыму, на северо-западном побережье Черного моря, Украине (кроме Закарпатья), Беларуси, России и Польше Морфологическая изменчивость высока (в частности, у ряда особей из Закавказья нет паховой петли). Однако значение морфологических признаков для таксономии не установлено.
Описание. Морфологические различия Hyla orientalis и H. arborea не выражены, в частности, не найдено существенных различий по мерным признакам.
Животные небольших размеров, длина тела половозрелых особей обычно до 50–60 мм. Пальцы задних конечностей соединены перепонкой. Кожа на спине гладкая, на брюшной стороне крупнозернистая. В спинной окраске чаще всего преобладают однотонные ярко зеленые цвета, хотя встречаются светло серые, коричневатые или серо голубоватые особи. Брюшная сторона белого цвета. Окраска верхней (спинной) и нижней (брюшной) сторон тела разделена двумя контрастными темными полосами с желтобелыми краями, каждая из которых тянется по обоим бокам тела от глаз до паховой области, образуя здесь петлю. Темные полосы есть также и на морде, где они соединяют ноздри и верхне передние края глаз.
Распространение. Встречается на Кавказе и прилежащих территориях; в Малой Азии; на западном побережье Черного моря; в Крыму, на северо-западном побережье Черного моря, Украине (кроме Закарпатья), Беларуси, России и Польше. В Крыму встречается только в горно-лесной части.
Образ жизни. Населяет хорошо освещенные участки широколиственных и смешанных лесов, кустарники, сады, виноградники, парки, берега водоемов и потоков. Темных и густых лесов избегает. Луга использует в основном при размножении. В лесостепи населяет островные леса и плавни. Встречается в антропогенных ландшафтах, в том числе в крупных городах (например, Киеве и Львове). Размножается обычно около древесной растительности, в стоячих водоемах.
Предпочитает высокие температуры. Спаривание при температуре воздуха +11°С, воды 15°C , почвы +9°C. Днем обычно сидит на растении, активна в основном вечером и ночью, когда спускается на землю. После ночного восполнения потерь воды вес особи увеличивается на 21–43%. Зимовка с октября – ноября по февраль – начало мая, обычно по апрель. Зимует в земле, под листьями и мхом, в норах, кучах камней и дуплах деревьев. В сухие бесснежные зимы происходит массовая гибель.
Размножается в апреле – мае, иногда в марте, июне – июле Самцы держатся в водоемах дольше самок. Спаривание иногда начинается на суше по дороге к воде. Крик самца состоит из громких коротких звуков, напоминающих карканье.
Полностью развитые личинки ведут в основном пелагический образ жизни. Метаморфоз со второй половины июня по сентябрь. После метаморфоза молодые остаются на берегах водоема, забираясь на траву и небольшие кустарники, иногда большом количестве. Половая зрелость у закавказских особей достигается обычно после 3–4 зимовок, максимальная продолжительность жизни оценена в 12 лет
Питается в основном насекомыми. В период размножения периодически питается на берегах и высоких стеблях растений, выступающих из воды. Отмечен каннибализм в форме личинка–яйцо При опасности H. orientalis в редких случаях принимает защитную позу: выгибает тело, закидывает на спину вывернутые задние конечности, и прикрывает голову вывернутыми передними .
Состояние популяции. В настоящее время очень редка там, где сохранились северные популяции. Сокращение численности происходит не только на севере, но и на юге.
Источники:
- Кузьмин С.Л. Земноводные бывшего СССР. — М.: Товарищество научных изданий КМК, 1999
Отряд: Бесхвостые — Anura
Семейство: Жерлянки — Bombinatoridae
Род: Жерлянка — Bombina
Краснобрюхая жерлянка — Bombina bombina (Linnaeus, 1761)
 Описание. Небольшие животные, размеры туловища чаще всего не превышают 50–60 мм, спинная поверхность обычно серо коричневатых (иногда зеленых) тонов с нечетко выраженными более темными пятнами неправильной формы. Бугорки на спинной стороне округлые. У самцов краснобрюхих жерлянок имеется внутренний резонатор, и в сезон размножения на первом и втором пальцах передних конечностей развиваются брачные мозоли. Плавательные перепонки развиты незначительно. Окраска брюшной стороны обычно представлена красными или красно оранжевыми пятнами неправильной формы на темном фоне.
Описание. Небольшие животные, размеры туловища чаще всего не превышают 50–60 мм, спинная поверхность обычно серо коричневатых (иногда зеленых) тонов с нечетко выраженными более темными пятнами неправильной формы. Бугорки на спинной стороне округлые. У самцов краснобрюхих жерлянок имеется внутренний резонатор, и в сезон размножения на первом и втором пальцах передних конечностей развиваются брачные мозоли. Плавательные перепонки развиты незначительно. Окраска брюшной стороны обычно представлена красными или красно оранжевыми пятнами неправильной формы на темном фоне.
Достаточно сильное токсичное действие кожных секретов нередко отмечается в литературе как потенциальное защитное средство жерлянок. Специальные лабораторные исследования показали, что введение этого секрета в дозе 3000, 4000 и 5000 мг/кг приводит к гибели мышей на 4, 5 и 7 сутки в 20, 50 и 70% случаев соответственно (Федярова, 1973). В качестве другого специфического защитного способа также указывается на своеобразное поведение жерлянок, когда они демонстрируют апосематическую окраску — ярко окрашенное пестрое брюхо (животные выгибают вверх переднюю и заднюю части тела и конечности).
При сравнении числа аномалий в строении конечностей краснобрюхих жерлянок в условно чистых и в загрязненных водоемах (Днепропетровская обл.) установлено, что в чистых водоемах они проявляются в 16,1–21,6% случаев, а в загрязненных в 39,1–44,2%. И хотя точных экспериментальных доказательств того, что причиной аномалий является загрязнение, нет, все же можно предположить, что их появление связано с действием антропогенных факторов (Flax, Borkin, 1997, 2004).
Распространение. Ареал охватывает территорию Центральной и Восточной Европы (от Дании и Западной Германии) до Уральских гор на востоке, на юге — до Кавказских гор, на севере — до Финского залива (в Швеции, вероятно, интродуцирована). В Турции встречается на небольшом участке крайнего северо запада Анатолии, откуда, в основном по особенностям окраски, и был описан подвид B. bombina arifiyensi O zeti & Yilmaz, 1987. Однако специфика рисунка укладывается, скорее всего, в рамки изменчивости этого вида в европейской части, что дало основание для сомнения в валидности подвидового названия (Gollmann et al., 1997).
Вид распространен почти на всей территории Украины, исключая Крым, где краснобрюхая жерлянка была обнаружена лишь однажды только в его северной части (Красноперекопский р н, с. Новопавловка). Места обитания краснобрюхих жерлянок на юге Украины чаще всего связаны с водоемами в долинах степных рек. В связи с расширением сети оросительных систем в этом регионе (особенно в связи с существованием Северо Крымского канала, берущего свое начало на материке), можно предположить возможность дальнейшей экспансии этого вида в Крыму. В горы краснобрюхие жерлянки обычно не поднимаются выше 200–350 м. В Прикарпатье распространение этого вида практически совпадает с границей собственно Карпат, а в Закарпатье — с территорией Закарпатской равнины. На юго западе доходит до Николаевской (Врадиевский р н) и Одесской (Килийский р н) областей, на юге — до Херсонской (Цюрупинский р н) и Днепропетровской (Никопольский р н) областей, на юго востоке — до Харьковской (Изюмский р н) области.
Образ жизни. Животные встречаются в большинстве типов биотопов на территории Украины, при этом характеризуются достаточно сильной привязанностью к стоячим или слабопроточным водоемам, в которых, как правило, предпочитают мелководье (глубина до 0,5 м или немного более). В Карпатах их отмечали в более чистых водоемах, чем те, в которых живет желтобрюхая жерлянка, хотя в южной части своего ареала B. (B.) bombina может обитать в участках, загрязненных промышленными и бытовыми отходами. Не исключено, что краснобрюхая жерлянка отдает предпочтение водоемам с бoльшим развитием водной растительности, избегая при этом водоемов с песчаными берегами.
Весеннее появление приходится на март — апрель и может смещаться в зависимости от погодных условий и широты местности. В среднем течении Южного Буга весеннее пробуждение отмечали в первой половине апреля при температуре воздуха 14,5°C и воды 9°C. Примерно через две–три недели после пробуждения животные переходят в водоемы в связи началом сезона размножения. В этот период жерлянки могут встречаться как в ночные, так и в дневные часы, хотя иногда отмечали некоторое повышение активности в промежуток от 10 до 12 и от 18 до 20 часов. Максимальная же вокализация самцов в период нереста приходится на сумеречно ночное время.
Численность яиц в кладке невелика, и по данным, полученным в Карпатах, составляет примерно 250–300 икринок. Икра откладывается поодиночке или порционно в виде комков по 15–30 штук. Для популяций средней части Южного Буга отмечено наличие в каждом комке по 5–8 икринок, при общем количестве икры в кладке 300–350 штук (Гончаренко и др., 1978). По сведениям из других участков ареала (Россия, Окский заповедник), размеры кладок колеблются от 181 до 926 яиц (в среднем 455). Продолжительность жизни в естественных условиях составляет около 12 лет, хотя для Западной Европы приведены несколько большие показатели: до 16 лет у самцов и до 14 у самок (Guarino et al., 1985), однако для большей части популяций эти сроки все же короче (около 8 лет). При этом было выяснено, что возраст самцов (напр. в южно итальянских популяциях) соответствует в среднем 8,0 ± 0,24 годам, а самок 8,3 ± 1,8 (Guarino et al., 1995). Согласно некоторым исследованиям за 10 лет популяция обновляется на 85–88% (Панченко, 1989).
Считается, что краснобрюхие жерлянки относительно теплолюбивые животные и оптимальные температуры, при которых они наиболее активны, лежат в пределах 18–25°C.
Сезонная активность продолжается до конца сентября — октября, после чего животные уходят на зимовку. Зимовать они могут как в водоемах (на дне в иле), так и на суше в полостях почвы, норах грызунов и пр. Есть указания о том, что в зимовочных скоплениях 26–33% могут составлять сеголетки и 67–74% взрослые (Гончаренко и др., 1978). Зимовочные укрытия на суше обычно расположены в районе водоемов, в радиусе 400–600 м. На зимовках отмечались скопления до 70 особей.
Личинки краснобрюхих жерлянок питаются в основном высшими растениями и водорослями, хотя при этом нередко поедают и беспозвоночных (Protozoa, Rotatoria, Microcrustacea и др.). В состав пищи взрослых животных входят различные виды наземных и водных беспозвоночных (Chironomidae, Chrysomelidae, Lepidoptera и др.), доля которых в составе питания может колебаться от 21 до 56%. При сравнении рациона жерлянок и обитающей в этих же биотопах озерной лягушки (Р. ridibundus) в России (Поволжье) установлено, что оба вида являются в той или иной степени пищевыми конкурентами, и перекрывание их трофических ниш составляет от 34,1 до 63,8%.
Состояние популяции. В подходящих для обитания биотопах численность краснобрюхих жерлянок довольно высока и может достигать 20–200 ос. /га. При исследовании в бассейне Южного Буга установлено, что весной численность этих животных на хорошо прогреваемых участках водоемов может достигать 8,1 ос. /м2, а в летний период — обычно 30–40 ос. /га (Гончаренко и др., 1978). По данным маршрутных учетов численность жерлянок на 100 м береговой линии в Карпатах составляет 15–40 особей (Щербак, Щербань, 1980), на юго западе Украины (Дунайский биосферный заповедник) — до 50 особей.
Несмотря на относительную пластичность вида, некоторые антропогенные факторы могут существенно влиять на его численность. Так, в деструктивных биогеоценозах (рудно угольные карьерные ландшафты в степном Приднепровье) численность краснобрюхой жерлянки составляет 2–10,2 ос. /га, а в местах, не подверженных техногенному прессу, — от 45,1 до 58,0 ос./га.
Вместе с тем следует отметить, что в некоторых случаях действие антропогенных факторов может приводить к повышению численности этих амфибий. Так, в отстойниках сахарных заводов в Белгородской области России, несмотря на то, что вода отличается здесь повышенной жесткостью и высоким содержанием нитритов, плотность жерлянок в благоприятные годы достигает 41 тыс. ос./га, а в засушливые годы эти водоемы являются убежищами, и численность жерлянок падет до 400–600 ос./га.
Среди антропогенных факторов, влияющих на снижение численности этого вида, в первую очередь указывают на уменьшение площади водоемов и их загрязнение промышленными и бытовыми отходами.
Вид внесен в списки конвенции об охране дикой фауны и флоры и естественной среды обитания в Европе (Берн, 1979; приложение II — «виды, подлежащие особой охране»), а также в Красную книгу Международного Союза Охраны Природы (категория «низкий риск исчезновения / зависит от охраны»).
Источники:
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
Отряд: Бесхвостые — Anura
Семейство: Жерлянки — Bombinatoridae
Род: Жерлянка — Bombina
Желтобрюхая жерлянка — Bombina variegata (Linnaeus 1758)
 Систематическое положение. В настоящее время различают 2–4 подвидa желтобрюхой жерлянки. При этом не исключено, что апеннинский подвид B. (B.) variegata pachypus (Bonaparte, 1838) и балканский подвид B. (B.) variegata scabra (Kuster, 1843), скорее всего, являются отдельными видами (Gollmann et al., 1997; Vukov et al., 2006). Украинские популяции относятся к номинативному подвиду.
Систематическое положение. В настоящее время различают 2–4 подвидa желтобрюхой жерлянки. При этом не исключено, что апеннинский подвид B. (B.) variegata pachypus (Bonaparte, 1838) и балканский подвид B. (B.) variegata scabra (Kuster, 1843), скорее всего, являются отдельными видами (Gollmann et al., 1997; Vukov et al., 2006). Украинские популяции относятся к номинативному подвиду.
Описание. Амфибии небольших размеров, длина туловища 27–50 мм, окраска спинной стороны тела обычно оливково зеленоватых тонов с нечетко выраженными мелкими темными пятнами. Брюшная часть яркой раскраски, представлена желтым или оранжевым фоном с крупными темными пятнами, площадь желтооранжевого фона обычно больше площади темных пятен. Желтооранжевый фон брюшной части переходит на нижнюю часть задних конечностей. На спинной стороне хорошо развиты бугорки, которые заканчиваются острыми шипиками. У самцов резонаторы отсутствуют. У них же в брачный сезон на первых трех пальцах передних конечностей и на нижней стороне предплечий развиваются брачные мозоли. Иногда встречаются морфы с зеленой окраской спины, при этом подчеркивается генетическая детерминированность этого признака и его адаптивный характер.
Распространение. Вид распространен в Европе, где приурочен к горным районам, однако отсутствует на Иберийском полуострове и на севере Италии (долина р. По), на востоке его распространение ограничено Украинскими Карпатами; граница ареала на севере не доходит до прибрежных районов Франции, Германии, Дании (в Англии интродуцирован). Обитает на большей части Апеннинского и Балканского полуостровов.
В Украине желтобрюхаямжерлянка обитает в горных и предгорных районах Карпат, где поднимается до высоты почти 2000 м. В Закарпатской области встречается почти повсеместно, за исключением крайних юго западных равнинных участков. Во Львовской области вид связан с горно предгорной юго западной частью, доходя на север до Перемышлянского и Старосамборского районов. В Ивано Франковской области обитает почти на всей территории (хотя не обнаружен в ее северной части). Распространение в Черновицкой области приурочено к ее западным районам, на восток доходит до г. Черновцы. Известно несколько находок желтобрюхой жерлянки в западной части Тернопольской области (Бучакский и Гусятинский р ны), что требует проверки.
Образ жизни. Желтобрюхая жерлянка характеризуется достаточно высокой экологической пластичностью в выборе биотопов, ее можно найти в разнообразных местообитаниях: в хвойных, лиственных и смешанных лесах, на полонинах, в кустарниках, на лугах и в полях. Здесь она встречается не только в стоячих или слабопроточных водоемах (озера, пруды, лужи, каналы, водохранилища), но иногда и в водоемах с относительно быстрым течением (по берегам ручьев и рек). Есть данные, что в Карпатах желтобрюхие жерлянки менее требовательны к уровню загрязненности воды, чем B. (B.) bombina, и могут занимать водоемы с довольно высоким насыщением органикой, с высокой соленостью или даже в минеральных источниках.
Желтобрюхие жерлянки менее привязаны к водоемам, чем B. (B.) bombina. Однако для них также характерно увеличение миграционной активности в периоды дождей и при сырой погоде. При сравнении суточной активности желтобрюхой и краснобрюхой жерлянок особых отличий не отмечено, они примерно сходны . Уход на зимовку часто зависит от погодных условий и высоты обитания, но чаще всего приходится на конец сентября — начало ноября. Обычно зимовка проходит на суше в полостях под камнями, упавшими деревьями, норах грызунов и пр., хотя активных желтобрюхих жерлянок иногда отмечали в воде термальных источников и зимой. На зимовках отмечена вместе с другими видами не только амфибий (тритоны, жабы и лягушки), но и рептилий — живородящие ящерицы, обыкновенные ужи (Щербак, Щербань, 1980).
Состав пищи желтобрюхих жерлянок несколько отличается от питания B. (B.) bombina за счет большего числа беспозвоночных, живущих на суше (иногда до 81%). Так, в их рационе отмечены Formicidae (до 15–36,8%), наземные формы Diptera (13,2–16,8%); существенна доля различных видов моллюсков, жуков (иногда до 78,3%) и пр.
Начало сезонной активности в Закарпатье обычно приходится на первую половину марта, а в Прикарпатье — на конец марта. Почти сразу же после пробуждения (примерно через 5–10 суток) при прогревании воды до 12°C животные приступают к спариванию и икрометанию. Период икрометания растянут и может длиться до августа, при этом в одном и том же водоеме могут быть не только особи в амплексусе, но также и головастики с разной степенью развития конечностей, или проходящие метаморфоз.
Обычно самка откладывает около 100 или немногим более икринок в виде одной или порционных кладок (по 10–30), часто прикрепляя их к различным подводным предметам: растениям, веткам, камням. Продолжительность эмбриогенеза в значительной степени зависит от температуры воды и составляет примерно 5–12 суток.
Среди естественных врагов B. (B.) variegаtа, также как и красно брюхой жерлянки, указаны многие околоводные позвоночные: их личинок поедают тритоны (Triturus cristatus), взрослых животных — ужи (водяной и обыкновенный), некоторые виды птиц (аист и др.). Для желтобрюхих жерлянок также характерно своеобразное защитное поведение в виде демонстрации апосематической окраски — ярко окрашенного брюха и нижней стороны конечностей.
Состояние популяции. Численность B. (B.) variegаtа на большей части ее ареала в пределах Украины довольно высока (часто превышает 1 особь на 0,02–20 м2).
Источники:
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
Отряд: Бесхвостые — Anura
Семейство: Чесночницы — Pelobatidae
Род: Чесночница — Pelobates
Обыкновенная чесночница — Pelobates fuscus (Laurenti, 1768)
 Описание. Тело коренастое, голова крупная, задние конечности короткие. Внутренний пяточный бугор очень большой, коричневато-желтый. Сверху серовато-желтая, серовато-коричневая или коричневая с темно-коричневыми пятнами и мелкими красноватыми точками. Брюхо серовато-белое, без рисунка или с редкими серыми точками. Лоб между глазами более-менее выпуклый. Самцы отличаются от самок выступающей овальной железой на плече, меньшим размером тела, более темной окраской, более контрастным рисунком на спине. В период размножения у самца есть мелкие бугорки на ладонях и предплечьях.
Описание. Тело коренастое, голова крупная, задние конечности короткие. Внутренний пяточный бугор очень большой, коричневато-желтый. Сверху серовато-желтая, серовато-коричневая или коричневая с темно-коричневыми пятнами и мелкими красноватыми точками. Брюхо серовато-белое, без рисунка или с редкими серыми точками. Лоб между глазами более-менее выпуклый. Самцы отличаются от самок выступающей овальной железой на плече, меньшим размером тела, более темной окраской, более контрастным рисунком на спине. В период размножения у самца есть мелкие бугорки на ладонях и предплечьях.
Распространение. Распространена от востока и севера Италии, европейской части Турции, юга Центральной Европы на север до Эстонии, Швеции, центральной России, среднего Предуралья и Западной Сибири (в горных частях Урала и Карпат отсутствует). В Крыму распространена спорадически. В Черном море, на крупных островах в реках и водохранилищах в умеренном поясе европейской части и т.д. Диапазон населяемых высот 0–500 м над ур. моря.
Образ жизни. Населяет хвойные, лиственные и смешанные леса и их опушки, рощи, степи, лесостепь, поля, луга, парки и сады. В лесах выбирает открытые участки. На суше ведет полуроющий образ жизни, что ведет к предпочтению мягких почв и избеганию плотных и каменистых. Плотность населения максимальна на песчаных и распаханных почвах. В подходящих биотопах Украины обилие достигает от нескольких десятков до нескольких тысяч особей на гектар.
Икрометание и раннее развитие проходят обычно в постоянных, стоячих, более-менее глубоких (до 20–130 см) водоемах, покрытых густой травянистой растительностью: в прудах, озерах,затопленных карьерах, канавах и т.д. Берега таких водоемов, как правило, пологие. Обычно в период размножения не образует больших скоплений. В то же время, в Предкавказье на 14 м2 водоема отмечено 27 кладок. Предпочитает теплую, довольно чистую, слабокислую воду с низкой минерализацией и отсутствием нитратов, но иногда размножается и в сильно загрязненной воде. В качестве убежищ иногда использует норы других животных и полости под камнями. Но чаще сама роет норки в почве. Эти норки выкапывает роющими движениями задних конечностей, используя большой внутренний пяточный бугор. Закапываясь, использует задние конечности по очереди и постепенно погружается в почву задом вперед. Погрузившись в почву, выталкивает передними конечностями грунт вперед и засыпает выход из норки. В дождливую погоду закапывается на 5–35 см, а во время засух – на 70–200 см. Молодые головастики обычно держатся на дне, незадолго до метаморфоза в основном среди растений в толще воды и часто появляются у поверхности. Недавно завершившие метаморфоз молодые закапываются на берегах и часто зимуют там же. В других случаях головастики зимуют в незамерзающих водоемах и метаморфизируют в конце мая – начале июня.
Чувствительна к качеству воды и структуре почвы. Загрязнение водоемов промышленными отходами, пестицидами, минеральными удобрениями, скотом, домашними отходами и т.п. опасно для головастиков. Другие типы деятельности человека (разрушение лугов, особенно скотом, урбанизация, рекреация, открытые колодцы в местах концентрации чесночниц и т.д.) также имеют отрицательные последствия.
Состояние популяции. Статус в Красном списке МСОП – LC. На периферии ареала редкий и/или сокращающийся вид.
Источники:
- Кузьмин С.Л. Земноводные бывшего СССР. — М.: Товарищество научных изданий КМК, 1999
Отряд: Бесхвостые — Anura
Семейство: Чесночницы — Pelobatidae
Род: Чесночница — Pelobates
Чесночница Палласа — Pelobates vespertinus (Pallas, 1771)
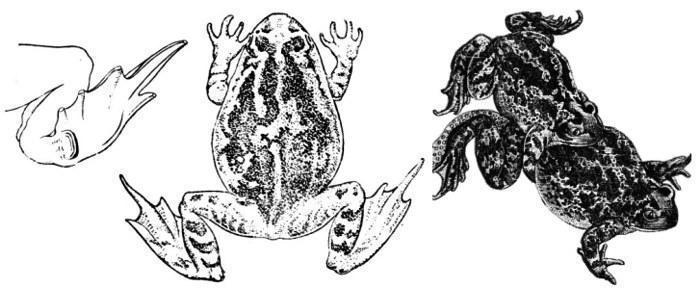
Pelobates fuscus — Обыкновенная чесночница (внешне весьма похожа на Pelobates vespertinus)и ее лопатообразный пяточный бугор (слева); ингвинальный амплексус чесночниц (справа)
Систематическое положение. В начале XIX века при изучении изменчивости обыкновенной чесночницы, Pelobates fuscus (Laurenti, 1768), было выяснено, что ее ареал населяют две формы, отличающиеся по размеру ядерного генома. По их результатам для восточной формы, отличающейся более крупным геномом, восстановлено название Pelobates vespertinus (Pallas, 1771).
Описание. Дальнейшее описание относится к Pelobates fuscus, но, вероятно, относится и к Pelobates vespertinus.
Напоминает некрупную жабу, отличаясь от нее вертикальным зрачком, отсутствием крупных ядовитых желез и лопатообразным ороговевшим пяточным бугром. Череп с обширными кожными окостенениями, формирующими характерный бугор на затылке. Длина тела до 71 мм. Головастики могут быть очень крупными (в некоторых случаях до 22 см).
Распространение. Граница между восточной и западной (собственно P. fuscus) формами проходит по западной части Харьковской области. Возможно, в фауну Харьковской области входит и Pelobates fuscus. Внешние отличия между двумя формами невелики; сравнение экологических особенностей двух форм до настоящего времени должным образом не выполнено.
Образ жизни. Населяет степи, лесостепь и смешанные леса в Центральной и Восточной Европе и Западной Азии. На территории Украины представлена двумя формами: западной и восточной. В Харьковской области обитает восточная форма.
Сухолюбивый вид, замечательно приспособленный к закапыванию (даже в сухую почву). Голос самца — негромкое похрюкивание. Кладка икры — толстый шнур с несколькими рядами икринок. Самец при амплексусе удерживает самку впереди бедер («за талию», ингвинальный амплексус), а не позади плечей («под мышками», аксиллярный амплексус). Головастики чесночницы крупные, и иногда достигают длины 175 мм. Сеголеток чесночницы намного меньше головастика (хотя и крупнее сеголетков других наших бесхвостых).
Состояние популяции. Многочисленный вид в окрестностях биостанции. Нерестится в стоячих водоемах — во всех прудах, временных водоемах в пойме, старицах. Наземные биотопы чесночницы вне периода размножения довольно разнообразные, но, как правило, наибольшей численности она достигает на открытых местах и на песчаных почвах: пойменных лугах, вырубках в бору и т. п.
Источники:
- /Pelobates_vespertinus
- Атемасова Т. А., Влащенко А. С., Зиненко А. И., Токарский В. А., Шабанов Д. А., Шандиков Г. А. Учебно-полевая практика по зоологии позвоночных. — Х.: ХНУ имени В. Н. Каразина, 2008. — 180 с.
Отряд: Бесхвостые — Anura
Семейство: Жабы — Bufonidae
Род: Жаба — Bufo
Серая или обыкновенная жаба — Bufo bufo (Linnaeus, 1758)
 Описание. Животные средних и крупных размеров, длина тела половозрелых особей около 60–90 мм, хотя самки могут быть значительно крупнее (более 100 мм). Паротиды выражены хорошо. Размер барабанной перепонки от 2–4 мм и более, у представителей карпатских популяций она не была выражена у 53,1% животных Внутренняя кожистая складка на предплюсне отсутствует, с нижней стороны самого длинного пальца задней ноги развиты парные сочленовные бугорки. На спинной стороне имеются многочисленные округлые бородавки, которые могут заканчиваться острыми шипиками (у самцов обычно выражены сильнее). Общий окрас тела коричнево серых тонов, при этом на спинной стороне часто имеются нечетко выраженные округлые темные пятна. Брюшная сторона серо белого цвета с размытыми мелкими пятнами неправильной формы. На нижней части паротид имеется черно коричневая полоса, которая обычно не переходит на бока туловища. Внешне половой диморфизм выражен в более мелких размерах самцов, в наличии у них брачных мозолей и, кроме этого, их окраска может быть не столь контрастной, как у самок В окраске сеголеток могут преобладать кирпично красноватые тона.
Описание. Животные средних и крупных размеров, длина тела половозрелых особей около 60–90 мм, хотя самки могут быть значительно крупнее (более 100 мм). Паротиды выражены хорошо. Размер барабанной перепонки от 2–4 мм и более, у представителей карпатских популяций она не была выражена у 53,1% животных Внутренняя кожистая складка на предплюсне отсутствует, с нижней стороны самого длинного пальца задней ноги развиты парные сочленовные бугорки. На спинной стороне имеются многочисленные округлые бородавки, которые могут заканчиваться острыми шипиками (у самцов обычно выражены сильнее). Общий окрас тела коричнево серых тонов, при этом на спинной стороне часто имеются нечетко выраженные округлые темные пятна. Брюшная сторона серо белого цвета с размытыми мелкими пятнами неправильной формы. На нижней части паротид имеется черно коричневая полоса, которая обычно не переходит на бока туловища. Внешне половой диморфизм выражен в более мелких размерах самцов, в наличии у них брачных мозолей и, кроме этого, их окраска может быть не столь контрастной, как у самок В окраске сеголеток могут преобладать кирпично красноватые тона.
Распространение. Обыкновенные жабы встречаются почти по всей Европе, за исключением Ирландии, севера Скандинавии и некоторых средиземноморских островов, на восток распространены до Забайкалья, имеются в Северно западной Африке.
В Украине эти животные распространены на большей части ее территории. На юге граница проходит примерно по среднему и нижнему течению Днепра — Днепропетровской и северу Запорожской областей, Кировоградской, северу Николаевской и Одесской областей; известны находки из окрестностей Херсона и низовий Дуная. На востоке ареал частично включает Харьковскую,Донецкую и Луганскую области. В Карпатах поднимается на высотудо 2000 м.
Образ жизни. Cерые жабы обитают в регионах с лесными ландшафтами, где их можно встретить в достаточно разнообразных биотопах, начиная от различных типов лесов (хвойные, широколиственные, смешанные) и заканчивая антропогенными насаждениями: рощи, парки, садовые участки городские скверы, изредка — лесопосадки. Открытых участков избегает, хотя и отмечалась на пахотных землях и пастбищах. Исследование серых жаб в Закарпатье показало их определенную привязанность к местам нереста. Так, из 62 помеченных в 1972 г. животных на следующий год на этом же месте (урочище Дубки, окр. Ужгорода) обнаружено 56% животных, еще через год — 35,2%. При имплантации датчиков и наблюдении с их помощью за миграциями этих животных в долине Рейна (Германия) установлено, что некоторые из них, хотя и мигрировали поздней осенью и зимой, но, в общем, в течение последнего сезона оставались в нижней части пойменной равнины.
Активные взрослые и сеголетки встречаются обычно при температурах воздуха +10–20°C, иногда при +24–26°C. Взрослые в период размножения активны круглосуточно, а потом лишь в сумерках, но в облачную и дождливую погоду также днем. Сеголетки активны днем и вечером. После зимовки появляется при температуре воздуха +5–6°C. Предельное расстояние миграций к местам размножения – 2,5 км.
Размножение в марте – июне, обычно в конце апреля – мае. Голос самца низкий и хриплый. Амплексус подмышечный. Одну самку часто обхватывают несколько самцов, а иногда самцы обхватывают даже рыб, тритонов, мертвых жаб. Часто несколько самцов пытаются обхватить одну самку, и образуются большие «шары», состоящие из многих жаб – до 20 особей.

Самец Bufo bufo в амплексусе с мертвой самкой
Охотится только на суше, в основном на ползающие формы беспозвоночных. Характерна мирмекофагия – поедание муравьев (как и у других видов Bufo). Иногда в одном желудке встречается несколько десятков муравьев. С этим согласуется тактика подстерегающего хищника, которую используют жабы.
Состояние популяции. Разрушение лесов и лугов, осушение водоемов составляют наиболее серьезную опасность. Эти факторы привели к вымиранию некоторых популяций. Загрязнение среды минеральными удобрениями и промышленными отходами, урбанизация, рекреация, гибель на автодорогах, бессмысленное уничтожение людьми и т.д. ведет к постепенному сокращению популяций в поселках и городах.
Статус в Красном списке МСОП – LC. Широко распространенный и нередкий вид на значительной части ареала, редкий в основном на периферии.
Источники:
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
Отряд: Бесхвостые — Anura
Семейство: Жабы — Bufonidae
Род: Жаба — Bufo
Зеленая жаба — Bufo viridis (Laurenti, 1768)
 Описание. Длина тела обычно 50–80 мм, окраска очень изменчива, но чаще всего верх тела оливково зеленых тонов с округлыми темно зелеными пятнами, выраженность и размеры которых часто варьируют в зависимости от пола (у самок пятна обычно крупнее и более контрастные) и географического распространения (самцы южных популяций в восточной части ареала характеризуются меньшей пятнистостью, вплоть до ее полного отсутствия). Характер расположения пятен и их конфигурация в течение всей жизни не изменяются (Brauer, 1988). Брюшная сторона однотонного беловато серого цвета с более или менее выраженными мелкими пятнами часто неправильной формы. На спинной стороне имеются бородавки, степень выраженности и шиповатость которых увеличивается в популяциях в юго восточном направлении, что более характерно для самцов. На боках туловища имеются иногда мелкие бородавки красноватых оттенков. На предплечье и голени обычно по 3 поперечных пятна полосы. На внутренней стороне предплюсны развита кожистая складка, самый длинный палец задней конечности, как правило, имеет одинарные сочленовные бугорки, пальцы задних конечностей характеризуются развитием плавательных перепонок разной степени выраженности. Зрачок горизонтальный, барабанная перепонка имеется, позади глаз расположены округлые железы паротиды. Половой диморфизм кроме особенностей окраски и бородавчатости, выражен в более мелких размерах самцов, наличии у них внутреннего непарного горлового резонатора и развитии брачных мозолей на первых двух пальцах передних конечностей
Описание. Длина тела обычно 50–80 мм, окраска очень изменчива, но чаще всего верх тела оливково зеленых тонов с округлыми темно зелеными пятнами, выраженность и размеры которых часто варьируют в зависимости от пола (у самок пятна обычно крупнее и более контрастные) и географического распространения (самцы южных популяций в восточной части ареала характеризуются меньшей пятнистостью, вплоть до ее полного отсутствия). Характер расположения пятен и их конфигурация в течение всей жизни не изменяются (Brauer, 1988). Брюшная сторона однотонного беловато серого цвета с более или менее выраженными мелкими пятнами часто неправильной формы. На спинной стороне имеются бородавки, степень выраженности и шиповатость которых увеличивается в популяциях в юго восточном направлении, что более характерно для самцов. На боках туловища имеются иногда мелкие бородавки красноватых оттенков. На предплечье и голени обычно по 3 поперечных пятна полосы. На внутренней стороне предплюсны развита кожистая складка, самый длинный палец задней конечности, как правило, имеет одинарные сочленовные бугорки, пальцы задних конечностей характеризуются развитием плавательных перепонок разной степени выраженности. Зрачок горизонтальный, барабанная перепонка имеется, позади глаз расположены округлые железы паротиды. Половой диморфизм кроме особенностей окраски и бородавчатости, выражен в более мелких размерах самцов, наличии у них внутреннего непарного горлового резонатора и развитии брачных мозолей на первых двух пальцах передних конечностей
Распространение. От Ближнего Востока через Европу до Сибири и Средней Азии. Северная граница ареала проходит с запада на восток от Балтийского моря до Алтая, в европейской части достигает 58° с.ш., восточнее спускается к югу. Южнее этой линии населяет почти всю территорию бывшего СССР. Зеленая жаба в Украиневстречается почти повсеместно . В горных районах Карпат и Крыма поднимается до 1000 и более метров.
Образ жизни. Обитает в зонах леса, лесостепи, степи, полупустыни и пустыни. В зоне лесов населяет обычно открытые места и заросли кустарников, зачастую далеко от водоемов. В аридных районах встречается в основном во влажных местах: в оазисах, наберегах арыков и озер. Здесь использует арыки и каналы как коридоры для расселения. В горах обитает в основном ниже верхней границы леса и избегает каменистых склонов и альпийских лугов. Размножается в очень разнообразных водоемах: болотах, прудах, озерах, заводях ручьев и рек, водохранилищах, канавах, лужах и т.д. Как правило, эти водоемы не глубже 50 см. Икра откладывается как в пресные, так и соленые водоемы (например, с минерализацией 8–12о/оо.
Переносит очень высокие температуры. Верхний предел, по-видимому, лежит ок. +40°C. Наибольшая подвижность особей отмечена при температуре тела +16–22°С, воздуха +21–22°C и почвы22°С.
Взрослые активны в основном в сумерках и ночью. Днем скрываются под бревнами, камнями, в норах и т.д. В мягкой почве выкапывают норки длиной до 20–30 см.
В период размножения активны и днем. Однако и вне его взрослые нередко проявляют дневную активность, даже на открытых местах и в солнечную погоду. Днем активных B. viridis легче встретить в сухих южных районах и в горах, чем в средней полосе России и Украины. В жаркие дни жабы нередко держатся в мелких водоемах. Совершают миграции на расстояния до 2–5 км от мест размножения. Головастики активны днем; они скапливаются в хорошо прогреваемых, открытых, мелких местах. Ночью они мигрируют на глубину в связи с охлаждением мелководий. Размножение в феврале – июле, в зависимости от широты и высоты над уровнем моря, но на большей части ареала в апреле – мае.
Головастики питаются в основном детритом и водорослями. еголетки сразу после метаморфоза питаются Collembola, Coleoptera, Acarina и Diptera, взрослые – в основном ползающими формами беспозвоночных, в том числе Aranei, Coleoptera и т.д. У части жаб, направляющихся к водоему, в желудках встречаются насекомые и пауки. Однако у большинства особей имеется «брачный пост» – по крайней мере, во время миграций. Характерна мирмекофагия: муравьи зачастую составляют значительную часть диеты взрослых (но не сеголеток). Следовательно, эта черта развивается у особей после первой зимовки – по-видимому, в связи с возрастными изменениями пищедобывательной стратегии.
Состояние популяции. Разрушение лугов, осушение водоемов, урбанизация и рекреация могут вести к сокращению популяций. С другой стороны, разрушение лесов может быть благоприятно для этого вида, адаптированного к отрытым пространствам. Известно также отрицательное влияние загрязнения среды пестицидами, минеральными удобрениями и промышленными отходами, а также отлова и бессмысленного уничтожения людьми. Зеленые жабы часто выходят на дороги вечером и/или после дождей, что ведет к гибели под колесами автотранспорта. Например, в Киевской обл. встречалось до 5 раздавленных B. viridis на 1,5 км шоссе.
Статус в Красном списке МСОП – LC. Широко распространенный, на большей части ареала обычный или многочисленный вид, редкий на периферии.
Источники:
- Кузьмин С.Л. Земноводные бывшего СССР. — М.: Товарищество научных изданий КМК, 1999
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
Отряд: Бесхвостые — Anura
Семейство: Жабы — Bufonidae
Род: Жаба — Bufo
Жаба камышовая — Bufo calamita (Laurenti, 1768)
 Описание. По внешнему виду отчасти напоминает зеленых жаб, но размеры тела существенно меньше и чаще всего составляют 50–70 мм. В окраске спинной стороны тела преобладают зелено оливковые тона с более темными округлыми пятнами, при этом обычно присутствует светлая или желтая продольная спинная полоса. Брюшная сторона светло серого или серо белого цвета с мелкими темными пятнами, часто неправильной формы. Одним из диагностических признаков камышовых жаб, является развитие у них пигментации также и на горле, в отличие от зеленой жабы. Зрачок горизонтальный, барабанная перепонка развита. Позади глаз расположены околоушные железы — паротиды, на всей спинной стороне беспорядочно разбросаны бородавки, которые у некоторых особей могут заканчиваться шипиками. На самом длинном пальце задних конечностей развиты парные сочленовные бугорки, внутренний край предплюсны несет кожистую складку. Половой диморфизм проявляется в несколько более мелких размерах самцов и частично выраженной у них шиповатости кожи, в наличии непарного внутреннего резонатора, развитии брачных мозолей на первых двух трех пальцах передних конечностей, часто меньшей контрастности рисунка.
Описание. По внешнему виду отчасти напоминает зеленых жаб, но размеры тела существенно меньше и чаще всего составляют 50–70 мм. В окраске спинной стороны тела преобладают зелено оливковые тона с более темными округлыми пятнами, при этом обычно присутствует светлая или желтая продольная спинная полоса. Брюшная сторона светло серого или серо белого цвета с мелкими темными пятнами, часто неправильной формы. Одним из диагностических признаков камышовых жаб, является развитие у них пигментации также и на горле, в отличие от зеленой жабы. Зрачок горизонтальный, барабанная перепонка развита. Позади глаз расположены околоушные железы — паротиды, на всей спинной стороне беспорядочно разбросаны бородавки, которые у некоторых особей могут заканчиваться шипиками. На самом длинном пальце задних конечностей развиты парные сочленовные бугорки, внутренний край предплюсны несет кожистую складку. Половой диморфизм проявляется в несколько более мелких размерах самцов и частично выраженной у них шиповатости кожи, в наличии непарного внутреннего резонатора, развитии брачных мозолей на первых двух трех пальцах передних конечностей, часто меньшей контрастности рисунка.
Распространение. В Европе камышовые жабы встречаются в ее западной и центральной частях к северу от Альп и Балкан, доходя до южной Швеции и Англии, на восток до стран Прибалтики и Калининградской области России, Белоруссии и Западной Украины, на юг — до северной Италии, Австрии, Чехии. В Украине встречается в западных областях — Волынской и частично во Львовской и Ровенской. Самой южной точкой ее находки являются окрестности с. Голоско Львовской обл. (49° 51' с. ш., 23° 56' в. д.), самой восточной — с. Дубровица Ровенской обл. (51° 34' с. ш., 26° 34' в. д.). Судя по очертанию восточной границы ареала и его частичному совпадению с январской изотермой –8°C, не исключено, что температурный фактор, как и длительность безморозного периода в 230 дней, могут быть определяющими распространение камышовых жаб на востоке их ареала.
Образ жизни. Bстречаются B. calamita в залесненных или открытых биотопах с легкими песчаными грунтами, которые обычно имеются на дюноподобных участках, окраинах и полянах сосновых лесов. Они также нередки в песчаных карьерах, полях, огородах, при этом условием их присутствия является, как правило, наличие соответствующей почвы, неглубоких водоемов и открытых участков в лесу или на месте бывших лесных насаждений. Украине камышовые жабы обитают в облесненных или открытых биотопах с легкими песчаными почвами. Обычно это дюнные участки, окраины и поляны сосновых лесов, парки, поля, огороды, песчаные карьеры.
Несмотря на достаточно обширный ареал, для вида характерна определенная ограниченность экологической пластичности. Так, хотя в спектр выбора ее биотопов, как уже отмечалось входят, среди прочего, участки, связанные с деятельностью человека (населенные пункты, огороды, картофельные поля, сады и пр.), но при этом обязательными фактором является сочетание легких супесчаных почв, неглубоких водоемов и открытых участков лесной зоны или мест сведенных лесов.
Размножение с апреля по июль. Интенсивность брачных хоров максимальна вечером. Пары в амплексусе активны в темноте, а днем прячутся под водой. Амплексус подмышечный. Иногда два самца обхватывают одну самку. Кладки икры обвиваются вокруг подводной травы и ветвей на мелководьях.
Питается в основном ползающими формами беспозвоночных.
Состояние популяции. Отмечена гибель от автотранспорта на дорогах. По-видимому, ряд популяций сокращается под влиянием антропогенных воздействий, в основном изменений биотопов. Подобно B. viridis, часто встречается в антропогенных ландшафтах – особенно на полях сельскохозяйственных культур, где обилие может возрастать. Создание карьеров с небольшими водоемами тоже оказывает положительное влияние.
Статус в Красном списке МСОП – LC. В целом на рассматриваемой территории это уязвимый узкоареальный вид, многие популяции которого сокращаются.
Источники:
- Кузьмин С.Л. Земноводные бывшего СССР. — М.: Товарищество научных изданий КМК, 1999
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
Отряд: Бесхвостые — Anura
Семейство: Лягушки — Ranidae
Род: Бурая лягушка — Rana
Остромордая лягушка — Rana arvalis Nilsson, 1842
 Описание. Животные средних размеров, длина тела половозрелых особей 40–64 мм. Материалы исследования внешнеморфологических признаков животных из Закарпатья показали, что длина тела самцов составляет около 52 мм, самок — 53 мм. Внутренний пяточный бугор высокий. Голова заострена, наспине имеются две продольные спинно боковые складки, в окраске преобладают коричневато светлые или оливково бурые тона, позади головы хорошо выражено темное образное пятно, брюшная сторона без пятен. По бокам головы расположены темные полосы, которые доходят до основания передних конечностей . Задние конечности развиты хорошо и, если их вытянуть вдоль тела (по направлению к голове), то голеностопное сочленение может только доходить до глаз (представители номинативного подвида), а у других — слегка заходить за конец морды (лягушки длинноногого подвида wolterstorffi). На задних лапах часто имеется рисунок из нескольких поперечных пятен полос, пальцы здесь соединяются плавательной перепонкой. Позади глаз расположена хорошо выраженная барабанная перепонка. У самцов есть внутренние резонаторы и развиты брачные мозоли на первых пальцах передних конечностей. Половой диморфизм в окраске особенно хорошо выражен в сезон нереста и проявляется в том, что для самцов характерно появление общего голубоватого фона.
Описание. Животные средних размеров, длина тела половозрелых особей 40–64 мм. Материалы исследования внешнеморфологических признаков животных из Закарпатья показали, что длина тела самцов составляет около 52 мм, самок — 53 мм. Внутренний пяточный бугор высокий. Голова заострена, наспине имеются две продольные спинно боковые складки, в окраске преобладают коричневато светлые или оливково бурые тона, позади головы хорошо выражено темное образное пятно, брюшная сторона без пятен. По бокам головы расположены темные полосы, которые доходят до основания передних конечностей . Задние конечности развиты хорошо и, если их вытянуть вдоль тела (по направлению к голове), то голеностопное сочленение может только доходить до глаз (представители номинативного подвида), а у других — слегка заходить за конец морды (лягушки длинноногого подвида wolterstorffi). На задних лапах часто имеется рисунок из нескольких поперечных пятен полос, пальцы здесь соединяются плавательной перепонкой. Позади глаз расположена хорошо выраженная барабанная перепонка. У самцов есть внутренние резонаторы и развиты брачные мозоли на первых пальцах передних конечностей. Половой диморфизм в окраске особенно хорошо выражен в сезон нереста и проявляется в том, что для самцов характерно появление общего голубоватого фона.
Распространение. Ареал занимает пространство от северо западной Франции, Бельгии, Нидерландов, Германии, Швеции и прилежащих районов Норвегии, Финляндии на юг до Альп, Хорватии, Словении, частично Италии, северной Румынии и на восток до Якутии, Алтая (Россия), доходит до северного Китая.
Современные исследования подтверждают более ранние данные о том, что в Украине остромордая лягушка обитает на ее большей части и приурочена к лесной и отчасти лесостепной зонам; в степной регион заходит только по долинам крупных рек — до низовий Днепра (окр. Голой Пристани), Южного Буга (среднее течение р. Чичиклия), имеется в дельте Дуная. В Крыму отсутствует. В Карпатах встречается только на равнинах и в предгорьях, в горы же может проникать по речным долинам, но обычно не выше 1000 м.
Образ жизни. Обитает в зонах тундры, лесотундры, леса, лесостепи и степи.
В Европе населяет в целом более сухие и открытые места, чем R. temporaria, в том числе опушки леса и поляны, болота, луга, поля, заросли кустарников, сады и т. д.
Размножение и раннее развитие происходят на мелководье стоячих водоемов (озер, прудов, болот, в лужах, канавах) площадью от нескольких до тысяч квадратных метров и глубиной до 2 м. В зоне симпатрии с R. temporaria размножается обычно в более открытых и мелких (т.е. лучше освещенных и прогреваемых) водоемах.
Размножение в марте – июне, обычно через несколько дней после окончания зимовки.
Питание. Головастики потребляют Chlorophyta, Diatomeae и другие водоросли, высшие растения, детрит и в небольшом количестве водных беспозвоночных. В период метаморфоза питание прекращается на короткое время и возобновляется еще до конца метаморфического климакса. Недавно метаморфизировавшие сеголетки охотятся на Acarina, Collembola и других мелких членистоногих. Взрослые потребляют в основном сухопутную добычу. Водные беспозвоночные (Gastropoda, Dytiscidae и т.д.) поедаются меньше. Имеется «Брачный пост».
Состояние популяции. Некоторые популяции сокращаются в результате разрушения биотопов, особенно вследствие урбанизации, рекреации и перевыпаса скота. Промышленное загрязнение тоже оказывает отрицательное воздействие. Загрязнение влияет и на городские популяции.
Статус в Красном списке МСОП – LC. Широко распространенный вид с высокой численностью, за исключением некоторых периферических популяций.
Источники:
- Кузьмин С.Л. Земноводные бывшего СССР. — М.: Товарищество научных изданий КМК, 1999
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
Отряд: Бесхвостые — Anura
Семейство: Лягушки — Ranidae
Род: Бурая лягушка — Rana
Прыткая лягушка — Rana dalmatina Bonaparte, 1840
 Описание. Окраска тела сходна с другими видами бурых лягушек и характеризуется преобладанием светло коричневых или бурооливковых тонов, на границе головы и туловища развито темное образное пятно, имеются спинно боковые складки. Начиная от ноздрей и далее с обеих сторон через область глаз и барабанных перепонок тянутся коричневые височные полосы. На задних конечностях обычно развиты поперечные темные полосы. Брюхо однотонно светлое. Барабанная перепонка крупная, по величине примерно соответствует размерам глаза . Резонаторы у самцов отсутствуют; в сезон нереста у них на первых пальцах передних конечностей развиваются брачные мозоли серо желтого цвета.
Описание. Окраска тела сходна с другими видами бурых лягушек и характеризуется преобладанием светло коричневых или бурооливковых тонов, на границе головы и туловища развито темное образное пятно, имеются спинно боковые складки. Начиная от ноздрей и далее с обеих сторон через область глаз и барабанных перепонок тянутся коричневые височные полосы. На задних конечностях обычно развиты поперечные темные полосы. Брюхо однотонно светлое. Барабанная перепонка крупная, по величине примерно соответствует размерам глаза . Резонаторы у самцов отсутствуют; в сезон нереста у них на первых пальцах передних конечностей развиваются брачные мозоли серо желтого цвета.
Распространение. Ареал этого европейского вида охватывает пространство от северной Франции и юга Швеции на запад до северо восточной Испании, на юге захватывает практически весь Балканский и большую часть Апеннинского полуостровов (исключая Сардинию и Сицилию), на востоке ареал ограничен Карпатами. Встречается на крайнем северо западе Турции.
Образ жизни. В выборе биотопов прыткая лягушка предпочитает разреженные широколиственные леса, включая заливные островные дубовые массивы, поляны, вырубки. Хвойных лесонасаждений и окультуренных ландшафтов (пастбища,поля и пр.) обычно избегает. Размножается в конце марта – мае.
Размножающиеся особи не образуют больших групп. Брачный крик самца слабый, напоминает квохтанье кур. Икра откладывается ночью на погруженную растительность в открытых частях водоема. Лягушки задерживаются в водоемах лишь на короткое время.
Состояние популяции. Данных о влиянии антропогенных факторов нет. Отрицательное воздействие могут оказывать разрушение широколиственных лесов и загрязнение водоемов. В больших количествах не отлавливалась. Статус в Красном списке МСОП – LC. Внесена в Красную книгу Украины как редкий и узкоареальный вид.
Источники:
- Кузьмин С.Л. Земноводные бывшего СССР. — М.: Товарищество научных изданий КМК, 1999
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
Отряд: Бесхвостые — Anura
Семейство: Лягушки — Ranidae
Род: Бурая лягушка — Rana
Травяная лягушка — Rana temporaria Linnaeus, 1758
 Описание. Лягушки средних размеров (максимальная длина тела 100–110 мм). Общий фон тела обычно характеризуется преобладанием светло коричневых тонов с наличием пятнистости. Как и для других представителей этой группы, достаточно типичным является развитие в затылочной области темного образного (лямбдовидного) пятна и темных боковых «височных» полос, идущих от кончика морды часто почти до основания передних конечностей. На спинной стороне тела обычно выражены темные пятна неправильной формы, которые у многих особей могут совпадать с двумя спинно боковыми складками кожи. У некоторых животных вдоль середины спины проходит светлая спинная полоса, которая чаще всего имеет нечеткие контуры. На задних конечностях развиты, как правило, поперечные полосы. Кожа гладкая с незначительным развитием мелких бугоров.
Описание. Лягушки средних размеров (максимальная длина тела 100–110 мм). Общий фон тела обычно характеризуется преобладанием светло коричневых тонов с наличием пятнистости. Как и для других представителей этой группы, достаточно типичным является развитие в затылочной области темного образного (лямбдовидного) пятна и темных боковых «височных» полос, идущих от кончика морды часто почти до основания передних конечностей. На спинной стороне тела обычно выражены темные пятна неправильной формы, которые у многих особей могут совпадать с двумя спинно боковыми складками кожи. У некоторых животных вдоль середины спины проходит светлая спинная полоса, которая чаще всего имеет нечеткие контуры. На задних конечностях развиты, как правило, поперечные полосы. Кожа гладкая с незначительным развитием мелких бугоров.
Окрас брюха представлен небольшими, прикасающимися друг к другу темными пятнами неправильной формы на грязно сером или желтоватом фоне. В брачный период горло самцов имеет голубоватый оттенок.
По сравнению с другими бурыми лягушками фауны Украины задние конечности относительно короче и, если их вытянуть к переди вдоль средней оси тела (по направлению к голове), то голеностопный сустав доходит лишь до области глаз или незначительно заходит за них. Размеры (высота) внутреннего пяточного бугра невелики, во всяком случае, он отчетливо меньше, чем у остромордой лягушки. На внутренней стороне первых пальцев передних лап самцов развиты брачные мозоли, разделенные бороздками на четыре более или менее выраженные доли. У них же имеются парные внутренние. Горловые резонаторы, а окраска горла в брачный сезон часто приобретает голубоватый оттенок.
Распространение. Вид распространен почти во всей Европе (исключая центральную и южную части Иберийского полуострова,южную Италию и южные Балканы), на восток встречается до Урала, интродуцирован в Ирландии. Из 4–6 европейских подвидов травяной лягушки в Украине обитает номинативный .В ископаемом состоянии травяная лягушка обнаружена в позднеплейстоценовых отложениях в Тернопольской области и в среднем плиоцене —среднем неоплейстоцене Одесской и Полтавской областей.
Украине травяная лягушка распространена в большей части северных, западных и восточных регионов страны, почти полностью отсутствует в степных районах . Здесь на юге граница распространения проходит по северу Одесской и Николаевской (с. Николаевка, долина р. Чичеклия) областей, Кировоградской области (р. Громоклея), северу Запорожской области, центральным районам Харьковской и Днепропетровской областей. В последней R. temporaria почти полностью исчезла и сейчас изредка встречается только в Днепровско Орельском заповеднике. Не исключено, что распространение этого вида может ограничиваться сильными весенними паводками, как, например, ее отсутствие в пойме реки Припять.
Образ жизни. Появление травяных лягушек в весенний период зависит от погодных особенностей года, но все же оно приходится на один из самых ранних сроков по сравнению с другими земноводными. Так, в Закарпатье (окр. Ужгорода), их отмечали уже в начале — середине марта, когда в водоемах еще были остатки льда (Щербак, Щербань, 1980). С увеличением высоты местности над уровнем моря сроки весеннего появления сдвигаются на более поздние сроки. Выбор подходящих водоемов для нереста часто обусловлен ландшафтными особенностями, но чаще всего это слабопроточные или стоячие водоемы в биотопах с древесной или древесно кустарниковой растительностью. Имеются сведения о том, что R. temporaria для откладывания икры может использовать также водоемы в трансформированных биотопах с примыкающими полями.

Икра Hyla arborea
Спаривание начинается практически сразу же после весеннего пробуждения, в среднем через 2–6 суток (Боркин, 1998). В это время самцы могут пытаться обхватить любой предмет, который двигается в пределах их видимости, из за чего возникает амплексус с особями других видов бесхвостых амфибий. Длительность нереста достаточно кратковременна и составляет в среднем 3–7 дней, при этом самцы в водоемах пребывают на несколько дней дольше самок. Травяные лягушки, как и многие другие виды бесхвостых амфибий Украины, характеризуются групповой кладкой, что проявляется в сосредоточении на местах размножения многих десятков пар особей. Обычно пик размножения травяных лягушек наступает на несколько дней раньше, чем у обитающих в этом же регионе R. Аrvalis.
Для травяных лягушек характерна сумеречно ночная активность. Днем скрываются в полостях под камнями, пнями, в листовой подстилке. При активной охоте и температуре приземного слоя воздуха 11–21°C, температура тела животного на 1–2,5 градуса выше.
Состояние популяции. Вид внесен в списки конвенции об охране дикой фауны и флоры и естественной среды обитания в Европе (Берн, 1979; приложение III — «виды животных, подлежащие охране»).
Источники:
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
Семейство: Саламандровые - Salamandridae
Подсемейство: Salamandrinae
Род: Саламандра — Salamandra
Пятнистая саламандра — Salamandra salamandra
 Описание. Животные характеризуются массивной округлой головой, широким туловищем, округлым в сечении хвостом; хвост короче тела. Глаза большие, выпуклые, позади них расположены выступающие удлиненные околоушные железы — паротиды. На передних конечностях по 4 пальца, на задних — по 5. Плавательные перепонки отсутствуют. Тело сегментировано 10–12 костальными бороздами, которые имеются и на хвосте . Небные зубы расположены в виде двух S образно изогнутых рядов, образуя бокаловидную фигуру. Окраска тела сверху интенсивно черного цвета с яркими желтыми или желто оранжевыми пятнами, брюхо однотонно черных или коричневатых тонов. Форма и расположение пятен достаточно изменчивы. Общие размеры тела (длина туловища с хвостом) саламандр могут составлять 140–180 мм. Половой диморфизм выражен в более выпуклой форме клоаки самцов (более четко проявляется в брачный период), в некоторых пропорциях тела и в несколько более крупных размерах самок. Так, длина тела вместе с хвостом самцов карпатських популяций в бреднем составляет около 155 мм, а самок — 164 мм.
Описание. Животные характеризуются массивной округлой головой, широким туловищем, округлым в сечении хвостом; хвост короче тела. Глаза большие, выпуклые, позади них расположены выступающие удлиненные околоушные железы — паротиды. На передних конечностях по 4 пальца, на задних — по 5. Плавательные перепонки отсутствуют. Тело сегментировано 10–12 костальными бороздами, которые имеются и на хвосте . Небные зубы расположены в виде двух S образно изогнутых рядов, образуя бокаловидную фигуру. Окраска тела сверху интенсивно черного цвета с яркими желтыми или желто оранжевыми пятнами, брюхо однотонно черных или коричневатых тонов. Форма и расположение пятен достаточно изменчивы. Общие размеры тела (длина туловища с хвостом) саламандр могут составлять 140–180 мм. Половой диморфизм выражен в более выпуклой форме клоаки самцов (более четко проявляется в брачный период), в некоторых пропорциях тела и в несколько более крупных размерах самок. Так, длина тела вместе с хвостом самцов карпатських популяций в бреднем составляет около 155 мм, а самок — 164 мм.
Распространение. В Украине встречается в Закарпатье и в некоторых районах прикарпатских областей (Львовской, Черновицкой, Ивано Франковской). Сведения об отдельных находках этих животных в более восточных регионах Украины (Волынская, Житомирская, Киевская, Днепропетровская обл.), скорее всего касаются животных, выпущенных на волю или убежавших из террариумов .
Образ жизни. Саламандры обитают в высотном поясе от 200 до 1500 м, обачно связаны с биотопами достаточно увлажненных лесных участков, чаще всего лиственных, смешанных и, реже, хвойных . Известны находки в горах и за пределами лесной зоны (на полонинах), однако, учитывая то обстоятельство, что эти находки были сделаны обычно не далее 200–400 м от границы леса, наличие саламандр свидетельствует, скорее всего, о существовании здесь леса в прошлом.
Начало активности весной в значительной степени определяется погодными условиями года: в общем, животные в Карпатах пробуждаются после зимней спячки достаточно рано — в марте, хотя иногда в теплые зимы в феврале отмечены случаи передвижения саламандр по еще сохранившимся участкам снега. Сезон размножения растянут почти на весь период активности, но чаще всего спаривание приходится на июнь и июль, личинки появляются спустя 9–10 месяцев. По наблюдениям за саламандрами карпатских популяций, во время спаривания, которому предшествует сложное видоспецифичное брачное поведение, самец откладывает на почву сперматофор, а затем самка захватывает его клоакой. Имеются сведения о том, что беременность и рождение личинок может протекать не только на протяжении двух сезонов (уход беременных самок на зимовку в ноябре и рождение личинок после окончания зимовки — примерно в апреле), но и происходить в течение одного года.
Яд кожных желез действует как нейротоксин и довольно токсичен для млекопитающих, его средняя летальная доза составляет 1,2 мг на 1 кг веса жертвы. Не исключено, что защитная функция кожних выделений также связана с их антигрибковым и антибактериальным действием.
Состояние популяции. Способность к обитанию в урбанизированных биотопах, по видимому, ограничена. Вместе с тем, специальные исследования в г. Овьедо (Испания) позволили обнаружить здесь более 40 местообитаний этого вида, которые находились вдоль железной дороги, в развалинах, среди куч мусора, камней и деревьев. Среди причин снижения численности саламандр указывается их отлов в природе (раньше для учебных целей, в настоящее время —чаще всего для коммерции), а также замена после сплошных вырубок бука искусственными елово-пихтовыми насаждениями.
Вид внесен в Красную книгу Украины (категория «уязвимые виды»), а также в списки конвенции об охране дикой фауны и флоры и естественной среды обитания в Европе (Берн, 1979; приложе ние III — «виды животных, подлежащие охране»).
Источники:
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
Отряд: Хвостатые - Caudata
Семейство: Саламандровые - Salamandridae
Подсемейство: Pleurodelinae
Род: Малый (гладкий) тритон — Lissotriton
Карпатский тритон - Lissotriton montandoni (Boulenger, 1860)

Систематическое положение. Наиболее близок к обыкновенному тритону, с которым иногда может гибридизировать в природе. Отличается от него отсутствием спинного гребня, формой тела (в сечении) и окраской головы и брюха (нет полосок и пятен). От гребенчатого и альпийского тритонов отличается отсутствием спинного гребня и окраской тела. От дунайского и других видов тритонов изолирован географически. От пятнистой саламандры отличается формой и окраской тела.
Описание. Некрупные тритоны, максимальная общая длина 96 мм (длина тела 47 мм). Форма тела в сечении, в отличие от других тритонов, не округло-овальная, а скорее четырехугольная. Голова небольшая, уплощенная сверху, с ребристыми краями спереди и с боков, с тремя продольными бороздками, соединяющимися около ноздрей. Небные зубы в виде двух продольных рядов, сначала параллельных, а затем резко расходящихся. Спинной гребень у самцов отсутствует. Есть хвостовая оторочка, более широкая по нижней стороне хвоста. У самцов черные подошвы задних ног и короткий, до 1 миллиметра, нитевидный вырост на кончике хвоста, в брачный период удлиняющийся до 5— 8 миллиметров, который пропадает при переходе к жизни на суше.
Тело бурого или буровато-коричневатого цвета с неясно выраженными темными пятнами. Посередине спины проходит более светлая полоса с неровными краями. У самок верхняя часть тела бывает желтоватой. Брюхо оранжевое, без пятен.
Распространение. Вид характерен для Карпатского региона, встречаясь на территории Украины, Румынии, Словакии, Чехии и Польши. В Украинских Карпатах обитает на высоте 150-2000 м над уровнем моря. Изолированная популяция известна в Львовской области (Ополье). Это самый многочисленный вид тритонов в Карпатах.
Образ жизни. Предпочитает ландшафты с лиственным или хвойным лесом, но населяет также и полонины. В период размножения обнаруживаются в самых разнообразных водоемах, как временных, так и постоянных — от луж и канав до крупных озер. Заселяют водоемы как с чистой родниковой, талой или дождевой водой, так и мутные водоемы с большим количеством разлагающихся органических остатков. Наибольшая численность вида зарегистрирована в канавах со слабопроточной родниковой водой, на склоне с выходами скал и смешанным лесом.
В водоемах и во время миграций тритоны активны днем. После выхода на сушу активны в сумерки и ночью, иногда в дождливую погоду днем. Зимуют на суше под камнями, в трухлявых пнях, норах грызунов. Весной пробуждаются в апреле-мае, но известны находки особей уже в конце марта, а на полонинах (безлесный участок верхнего пояса Украинских Карпат) в начале июня. Карпатский тритон — холодостойкий вид; активные особи наблюдались при температуре воды 2-4°С. После перехода в водоем происходит линька, причем кожа сходит целиком, начиная с головы.
В июле — начале августа взрослые особи покидают водоемы. На суше они прячутся под камнями, в лиственную подстилку, под коряги, гнилые стволы деревьев, обычно около озер и ручьев в тенистых лесных местах. Уход на зимовку отмечен в конце октября.
Брачные игры такие же, как у обыкновенного тритона, хотя частота отдельных элементов иная. Икрометание в мае-июне в зависимости от высоты местности и погоды. Самки откладывают икринки около берега, прикрепляя их к подводным растениям по одной, реже по 2-5 штук. Икринки размером 2,2-2,8 x 3,4-3,6 мм (диаметр яйцеклетки 1,7-1,8 мм), имеют овальную форму и пигментированы (коричневатые) сверху. Количество икринок на одну самку колеблется от 35 до 80 штук.
Выклев личинок длиной 6-7 мм обычно наступает через 10-15 суток, но может задерживаться до 30 суток. Личиночное развитие длится около 3 месяцев, и личинки длиной 26-35 мм большей частью успевают пройти метаморфоз до наступления зимы. Однако некоторые из них зимуют: известна находка личинок с развитыми жабрами и длиной 30-41 мм в конце февраля.
В водоемах карпатские тритоны кормятся личинками комаров-звонцов и других насекомых, ракообразными (дафний и т.д.), иногда поедают икру тритонов и жерлянок. На суше ловят различных насекомых (двукрылых, жуков, муравьев).
Весьма чувствительны тритоны к загрязнению воды, в том числе ядохимикатами, которыми обрабатывают лесные массивы. Тритоны чутко отзываются на вырубку лесов, и при их сведении исчезают.
Состояние популяции. Численность карпатского тритона стала сокращаться в начале двадцатого столетия. Из Германии он исчез еще до второй мировой войны. Сейчас основная зона его обитания находится на территории Украины, в том числе он встречается в Карпатском заповеднике. Как эндемик Карпат, обитающий на краю ареала, был внесен в Красные книгу и Украины. Охраняется Бернской Конвенцией.
Источники:
- Ананьева Н.Б., Боркин Л.Я., Даревский И.С., Орлов Н.Л. Земноводные и пресмыкающиеся. Энциклопедия природы России. М.: ABF, 1998
Отряд: Хвостатые - Caudata
Семейство: Саламандровые - Salamandridae
Подсемейство: Pleurodelinae
Род: Малый (гладкий) тритон — Lissotriton
Обыкновенный тритон — Lissotriton vulgaris (Linnaeus, 1758)
 Систематическое положение. По новой классификации этот вид относят к роду малых тритонов (Lissotriton). Тритон обыкновенный впервые был описан в 1758 году шведским натуралистом Карлом Линнеем.
Систематическое положение. По новой классификации этот вид относят к роду малых тритонов (Lissotriton). Тритон обыкновенный впервые был описан в 1758 году шведским натуралистом Карлом Линнеем.
Описание. Тритон обыкновенный (Triturus vulgaris или Lissotriton vulgaris) - самый распространенный вид тритонов.
Один из самых мелких тритонов в фауне Украины. Общая длина тела взрослых особей - около 55-105 мм. Небные зубы в виде двух линий, сближающихся друг с другом в своей дистальной части. Кожа гладкая, или малозернистая. Спина окрашена в оливково-бурые или серые тона, бока - в оливково-желтые с серебристые тона. Сверху и по бокам тела развиты относительно большие темные пятна (у самок не так выразительны). Окраска брюха желто-оранжевых тонов с хорошо развитыми небольшими темными пятнами. На голове заметны продольные темные полосы, две из которых выглядят контрастнее и проходят через глаза. Самцы в сезон размножения окрашены ярче, у них на теле и по бокам хвоста развиваются голубоватые полосы-пятна. Одновременно на спинной стороне у них развивается гребень, который в отличие от гребенчатого тритона (Triturus cristatus), при переходе на спинную часть не образует перерыва у основания хвоста. Не исключено, что в это время он может использоваться как дополнительный дыхательный орган. Также в брачный период на пальцах задних конечностей появляются перепонки. Половой диморфизм проявляется в больших размерах самцов. Так, средняя длина тела вместе с хвостом у особей мужского пола в среднем колеблется в пределах 72,01-72,06 мм, а у самок от 70,52 до 72,01 мм.
Распространение. Вид широко распространен на большей части Европы (за исключением всего Пиренейского полуострова, юга Апеннинского полуострова, южной части Франции и северной части Скандинавского полуострова). На востоке ареал захватывает часть Азии в Алтайский гор.
В Украине обыкновенный тритон водится в западных, центральных, северных и восточных областях. На юге его граница проходит по Одесской (Беляевский и Белгород-Днестровский районы), Николаевской (Врадиевский район) и Херсонской областями, далее в восточном направлении она смещается несколько севернее и идет Запорожской (остров Хортица), Харьковской и Луганской (Станично-Луганский район) областями.
Образ жизни. Обыкновенный тритон чаще всего связан с лесными регионами, хотя встречается и в других типах биотопов, в том числе в кустарниковых зарослях, парках, садах, лугах и даже населенных пунктах.
L. vulgaris проникает глубоко в степную зону лесистыми долинами рек. В горах (Карпаты) поднимается до высоты 600-1000 м, хотя в других участках ареала (Кавказ) его обнаруживали на высотах до 2700 м. Находят L. vulgaris и во временных водоемах (лужи на полях, в кюветах дорог).
На Закарпатье среднее количество тритонов на 100 м береговой линии колеблется от 7 до 60 особей. В восточных регионах (долина Днепра, Днепропетровская область) при отлове этих животных с помощью ловчих цилиндров учтено от 2,1 до 6,8 особей/10 суток, но при этом их численность зависела от типа и структуры лесных насаждений, близости водоема , степени увлажненности почвы и других факторов.
Питается тритон обыкновенный многоножками, насекомыми, гусеницами, разнообразными личинками, паукообразными, червями, моллюсками, ракообразными, водяными клопами, личинками жуков-плавунцов, однодневками, равнокрылых, икрой рыб и лягушек в водный период жизни.
В конце марта - в апреле тритоны мигрируют к водоемам. Через 5-9 дней приступают к размножению. Оплодотворению и отложению яиц предшествуют брачные игры. Самка откладывает 60-700 икринок (чаще около 150). Каждую икринки самка откладывает на отдельный листок подводного растения и заворачивает ее внутри листочка. Личинки появляются на 14-20 день после отложения икры.
В апреле 2000 г. среди украинских популяций обыкновенного тритона в Одесской области (Измаил) были впервые зарегистрированы два педоморфные( неотенические) особи. Эти животные характеризовались примерно такими же размерами, как и остальные тритоны, собранных в том же месте. Одна из личинок вскоре завершила метаморфоз в лабораторных условиях.
Явление педоморфоза у амфибий этого вида отмечено в Германии, Румынии, Венгрии, Швеции, Англии, бывшей Югославии. Так, с 14 популяций в Черногории в 4 из них обнаружены педоморфные особи, причем их количество колебалось в течение сезона от 44% в октябре до 80% в августе. Количество педоморфних особей было больше, чем нормальных (58,8% против 41,2%). Среди этих педоморфных личинок преобладали самки (42,22% против 14,57%).
Не исключено, что причиной такой структуры и динамики популяций может быть нестабильность биотопов и низкая миграционная активность этих животных, а явление педоморфоза при этом имеет адаптационный характер, существенно увеличивая изменчивость популяции в целом.
Обобщение сведений о причинах педоморфоза позволило сделать вывод, что они обычно связаны с низкими температурами при метаморфозе. Другими факторами могут быть голодание личинок, химический состав воды и корма (недостаток йода и витаминов).
Состояние популяции. Тритон обыкновенный занесен в Приложение III "Конвенции по охране дикой флоры и фауны и природной среды обитания в Европе" (категория "виды, подлежащие охране") и в "Красного Списка позвоночных Международного союза охраны природы (МСОП)".
Создание просек в лесах и ирригационных каналов способствует повышению численности популяций и расселению вида.
Источники:
- Писанец Е. Земноводные Украины (руководство для определения амфибий Украины и сопредельных стран).- М.: Издательство Раевского, 2007
Отряд: Хвостатые - Caudata
Семейство: Саламандровые - Salamandridae
Подсемейство: Pleurodelinae
Род: Средний тритон — Mesotriton
Альпийский тритон - Mesotriton alpestris (Laurenti, 1768)
 Систематическое положение. Вид включает 8-9 подвидов, из которых в нашей стране известен один — номинативный (M. alpestris alpestris).
Систематическое положение. Вид включает 8-9 подвидов, из которых в нашей стране известен один — номинативный (M. alpestris alpestris).
Описание. Редкий и один из наиболее красивых, ярко окрашенных тритонов. Средних размеров, длина обычно около 90 мм, половина которой приходится на хвост. Поверхность тела гладкая. Как и другие представители этого рода, имеет веретеновидное тело. Самцы сверху темные, серовато-коричневые с фиолетово-синим оттенком. С двух сторон тела, на конечностях и щеках ряд темно-синих пятен неправильной формы. Брюхо и горло огненно-оранжевые. Спинной гребень, переходящий в плавниковую складку хвоста, кажется клетчатым из-за чередующихся светлых и темных прямо-угольных пятен. Небные зубы хорошо развиты. Хвост в верхней части голубовато-серый, в нижней — оливково-серый с синими пятнами. Самки окрашены менее ярко, на спине они обычно имеют мраморный узор, спинной гребень у них не развит.
Распространение. Западная и Центральная Европа от Испании, Италии и Греции на север до Дании и на восток до Карпат включительно. В Украине встречается в горных и предгорных районах Карпат, включая Тернопольскую область и Буковину (Черновицкая обл.)
Образ жизни. Встречается от предгорий до вершин гор в тенистых и сырых местах, на высотах от 500 до 1500 м над уровнем моря. В северных и северо-восточных районах Карпат редок, а на Буковине в лиственных и смешанных лесах, напротив, самый многочисленный вид среди хвостатых земноводных. На зимовку уходит в сентябре. Зимует в кучах камней и лесной подстилке, валежнике, под упавшими деревьями. Взрослые от мест зимовок перемещаются в апреле-мае к воде - в различные водоемы, в канавы и ямы, дорожные колеи, заполненные водой и часто загрязненные. Покидают воду в конце июля - в августе. На суше тритоны кормятся дождевыми червями, насекомыми, паукообразными, голыми слизнями. В воде ловят мелких ракообразных (дафний и др.), личинок насекомых (комаров, ручейников, поденок и др.), моллюсков.
Спаривание, сопровождаемое свадебными играми, происходит в воде. В мае — начале июня самки откладывают 100-190 яиц (икринок) по одному или группами (по 3-5 штук) среди листьев подводных растений. Высоко в горах личинки остаются зимовать в воде. Известны случаи, когда личинки оставались в воде по нескольку лет и достигали длины 70—80 мм (явление неполной неотении).
Источник: Куриленко В.Є., Вервес Ю.Г. Земноводні та плазуни фауни України: Довідник-визначник . К.: Генеза, 1998.
Отряд: Хвостатые - Caudata
Семейство: Саламандровые - Salamandridae
Подсемейство: Pleurodelinae
Род: Крупный тритон — Triturus
Тритон гребенчатый - Triturus cristatus (Laurenti, 1768)

Описание. Кожа крупнозернистая. Окраска сверху черная или коричнево- черная, брюхо оранжевое с крупными черными пятнами. Гребень самца в брачном наряде зубчатый, прерывается (или резко снижается) у основания хвоста; по бокам хвоста голубовато-белая полоса. Самки всегда без гребня, но часто с тонкой желтой линией вдоль спины.
Распространение. Это европейский вид, распространенный от Франции до Урала.
На юге Украины обитают близкородственные виды, относительно недавно получившие самостоятельность: дунайский тритон (T. dobrogicus) – Закарпатская, Одесская и Херсонская области (низовья Дуная и Днепра), и тритон Карелина (T. karelinii) – Крым.
Образ жизни. Весну и первую половину лета гребенчатые тритоны проводят в водоемах, во второй половине лета живут в лесу. В водоемах тритоны активны преимущественно днем, на суше в сумерки и ночью, скрываясь в остальное время под пнями, гнилыми стволами деревьев, в ямах с песком и опавшими листьями, в густом дерне, иногда даже в кротовых ходах. В этих убежищах обычно и зимуют, иногда вместе с другими видами амфибий. Зиму проводят также под мхом, в корневых пустотах, даже в подвалах и погребах. Хотя зимуют обычно на суше, но могут зимовать и в непромерзающих до дна водоемах.
Из зимовок появляются в марте (Закарпатье), в апреле-мае (средняя Россия) в период вскрытия водоемов при температуре воздуха 9-10°С и температуре воды 6°С. Через 3-6 суток тритоны перемещаются к водоемам.
В отличие от обыкновенного тритона в качестве нерестилищ предпочитают более глубокие водоемы (до 0,5-0,7 м). Размножение начинается при температуре воздуха 14°С. Самцы к этому времени уже находятся в брачном наряде с высоким спинным гребнем, давшим название виду, и красивой голубой полосой вдоль хвоста. Этот гребень, богатый сосудами, по-видимому, усиливает также газообмен животного. После ритуального ухаживания самка откладывает от 80 до 600 икринок (чаще 150-200). Процесс икрометания занимает от двух недель до двух месяцев. Икринки имеют овальную форму размером 2,0-2,5 x 4,0-4,5 мм (размер яйцеклетки около 1,5 мм) и сверху не пигментированы. Икринки заворачиваются в листочки, причем обычно прикрепляются к их нижней стороне (по 1, реже 2-3 штуки); при этом выбираются более крупные листья, часто лежащие на поверхности воды.

Самка Triturus cristatus
Питается в водоемах несколько более крупными водными беспозвоночными, чем обыкновенный тритон. Среди кормов преобладают моллюски, водяные жуки, личинки насекомых, иногда даже тритоны поедают икру и головастиков амфибий. На суше в рацион входят, главным образом, дождевые черви, слизни, а также разнообразные насекомые (имаго и личинки).
Источники:
- Банников А. Г., Даревский И. С., Ищенко В. Г., Рустамов А. К., Щербак Н. Н. Определитель земноводных и пресмыкающихся фауны СССР. — М.: "Просвещение",
- Атемасова Т. А., Влащенко А. С., Зиненко А. И., Токарский В. А., Шабанов Д. А., Шандиков Г. А. Учебно-полевая практика по зоологии позвоночных. — Х.: ХНУ имени В. Н. Каразина
Отряд: Хвостатые - Caudata
Семейство: Саламандровые - Salamandridae
Подсемейство: Pleurodelinae
Род: Крупный тритон — Triturus
Дунайский тритон - Triturus dobrogicus (Kiritzescu, 1903)

Систематическое положение. В списках батрахофауны бывшего СССР (Терентьев, Чернов, 1949; Банников и др., 1977) дунайский тритон (ранее рассматривался как подвид гребенчатого тритона Triturus cristatus dobrogicus) не приводился. Однако изучение внешнеморфологических признаков этих хвостатых амфибий из Одесской обл., Молдавии и Карпат (Осташко, 1977) послужило основанием для утверждения об обитании здесь переходных форм между номинативным подвидом гребенчатого тритона и T. cristatus dobrogicus. Еще позже (Щербак, Щербань, 1980), изучение батрахофауны Карпат позволило сделать заключение о распространении в Закарпатье наряду с тритонами номинативного подвида гребенчатого тритона и гибридных с T. cristatus dobrogicus популяций (но, как отмечалось выше, сейчас установлено, что это два разных вида: T. сristatus и T. Dobrogicus). Современные электрофоретические исследования белков и определение количества ядерной ДНК подтвердили вывод о видовой самостоятельности Triturus dobrogicus и Triturus cristatus, сделанный ранее при их изучении в других участках ареала (Litvinchuk et al., 1994; Litvinchuk et al., 1995). Сравнение этих двух видов в Чехии (материалы по аллозимам), также не показали интрогрессии генов вследствие естественой гибридизации (Pialek et al., 1997). Вместе с тем следует указать на доказательства того, что в генотипе дунайских тритонов могут встречаться «чужеродные» аллели, свойственные генным комплексам других видов. Так, биохимический анализ T. dobrogicus из окр. с. Балево (Закарпатье), показал наличие «чужеродного» аллеля эстеразы (ESTA 2), характерного для гребенчатого тритона. Такие «чужеродные» аллели, свойственны не только Triturus cristatus, но и тритону Карелина, были обнаружены у дунайского тритона из Сербии. Не исключено, что эти материалы могут свидетельствовать о существовании в прошлом генетических контактов T. dobrogicus с двумя другими видами тритонов (Litvinchuk et al., 1999). Более того, недавние исследования в Закарпатье (Морозов Леонов и др., 2003) позволили обнаружить гибридную особь между дунайским и гребенчатым тритонами, что подтверждает (отдельные случаи) скрещивание тритонов обоих таксонов в природе.
Описание. Животные средних размеров (длина тела с хвостом 115–120 мм), внешне похожи на гребенчатых тритонов, но отличаются более удлиненным туловищем и некоторыми другими размерными характеристиками. Общий фон тела коричнево темных тонов, с нечетко выраженными небольшими округлыми черными пятнами, на нижней стороне туловища обычно имеются мелкие светлые пятна неправильной формы, которые переходят на нижнюю часть головы, по бокам средней части хвоста проходит бело голубая полоса, брюхо красно оранжевого цвета с темными пятнами. Половой диморфизм проявляется в более крупной клоаке самцов и в развитии у них в сезон размножения на верхней части фестончатого гребня, который перерывается в конце тела и далее вновь продолжается на хвосте. Половой диморфизм проявляется также во вздутости горла самцов, особенно хорошо видным в сезон размножения.
Распространение связано с Дунайской и Паннонской равнинами, ареал покрывает территорию от устья Дуная вверх по его течению до Австрии, Чехии и Хорватии.
По современным представлениям дунайский тритон в Украине представлен номинативным подвидом Triturus d. dobrogicus (Kiritzescu, 1903) в Одесской, Херсонской и, вероятно, в Николаевской областях (в двух последних регионах необходимы дополнительные исследования по уточнению таксономической принадлежности и границ распространения), а также паннонским подвидом Triturus d. macrosomus (Boulenger, 1908) в Закарпатье. Представители последнего подвида отличаются от номинативного оранжево желтыми тонами окраски брюха против более ярко красных оттенков у Triturus d. dobrogicus (Litvinchuk, Borkin, 2000), меньшим значением индекса Lt.c./L. (на 13,8% у самцов, и на 13,3% у самок), меньшим значением индекса IW (у самцов этого подвида менее 54,0 у T. d. dobrogicus — более), числом туловищных позвонков, которое у них в среднем составляет 17, а у тритонов номинативного подвида 16 (по другим данным это число у T. dobrogicus в общем несколько больше — в среднем 17,5; Arntzen, Wallis, 1999).
В пределах Украины распространен в Одесской области (дельта Дуная) и в Закарпатье. Особый интерес вызывает находка одной неполовозрелой особи T. dobrogicus за пределами основного ареала в окрестностях Голой Пристани (Херсонская обл.), выявленная при анализе коллекций Зоологического музея ННПМ НАНУ. Позже, в 2004 г., в окрестностях Херсона было добыто еще две личинки этого вида тритонов (Litvinchuk, 2005), что дало основание полагать, что дунайские тритоны дельты Днепра представляют собой изолированную популяцию как от остальной части ареала T. dobrogicus, так и от T. cristatus. Специальный анализ питания не проводился, однако обычно упоминается, что это беспозвоночные, в том числе ракообразные, насекомые, черви, моллюски (Литвинчук, Боркин, 2002). Учитывая близость этого вида с T. cristatus, можно также ожидать совпадения спектров питания обоих видов. Следует отметить, что если для T. cristatus отмечали отрицательную связь в его распространении с водоемами, заселенными рыбами, то дунайский тритон нередко заселяет такие водоемы.
Состояние популяции. На современное состояние популяций, очевидно, оказывает негативное воздействие и антропогенный пресс. Сделано заключение о необходимости внесения этого вида в третье издание Красной книги Украины (категория «уязвимые виды»); вид включен в списки конвенции об охране дикой фауны и флоры и естественной среды обитания в Европе (Берн, 1979; приложение II — «виды, подлежащие особой охране»), а также включен в Красную книгу Международного Союза Охраны Природы (вид, которому угрожает опасность исчезновения, категория «недостаточно изученные»).
Источники:
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007
Отряд: Хвостатые - Caudata
Семейство: Саламандровые - Salamandridae
Подсемейство: Pleurodelinae
Род: Крупный тритон — Triturus
Тритон Карелина - Triturus karelinii (Strauch, 1870)
 Систематическое положение. Ранее этот тритон рассматривался как подвид гребенчатого тритона — Triturus cristatus karelinii (Терентьев, Чернов, 1949; Щербак, 1966; Банников и др., 1977), однако использование кариологических и биохимических методов исследования этих животных из разных участков их ареала, а также сравнение с другими родственными таксонами, позволило прийти к выводу о существенных отличиях и придать им статус самостоятельного вида (Macgregor et al., 1990;Litvinchuk et al., 1994; Межжерін та ін., 1999). По последним сведениям вид представлен двумя подвидами — Triturus karelinii karelinii и Triturus karelinii arntzeni (Litvinchuk et al., 1999).
Систематическое положение. Ранее этот тритон рассматривался как подвид гребенчатого тритона — Triturus cristatus karelinii (Терентьев, Чернов, 1949; Щербак, 1966; Банников и др., 1977), однако использование кариологических и биохимических методов исследования этих животных из разных участков их ареала, а также сравнение с другими родственными таксонами, позволило прийти к выводу о существенных отличиях и придать им статус самостоятельного вида (Macgregor et al., 1990;Litvinchuk et al., 1994; Межжерін та ін., 1999). По последним сведениям вид представлен двумя подвидами — Triturus karelinii karelinii и Triturus karelinii arntzeni (Litvinchuk et al., 1999).
Описание. Тритон средних размеров, длина тела (с хвостом) около 130 мм, внешне похож на гребенчатого тритона. Два ряда небных зубов, начинающихся впереди хоан, изогнуты и расходятся в своей задней части. Хвост с боков уплощен. Спинной гребень самцов в брачный период не такой высокий, как у гребенчатого тритона и его перерыв при переходе на хвостовую часть обычно выражен не так четко. Окраска тела от темно коричневого до светло желтого фона с нечеткими темными пятнами, на оранжевом или оранжево желтом брюхе — черные пятна. Цвет горловой части обычно ярко желтый (иногда до красноватых оттенков) с темными пятнами. Кожа крупнозернистая . Половой диморфизм выражен в развитии спинного гребня у самцов в сезон размножения, более крупных, по сравнению с самками, размерах и темном цвете клоаки (у самок клоака светлая).
По сравнению с другими тритонами этой группы, туловищных позвонков с ребрами у тритона Карелина обычно меньше — в среднем 14,2 (у T. dobrogicus и T. cristatus 17,5 и 16,0 соответственно — Arntzen et al., 1999).
Распространение. Обитает на востоке Балканского полуострова, в горной части Крыма, в Турции, далее на восток встречается в Предкавказье и на северном Кавказе, есть в северном Иране. В Украине обитают тритоны номинативного подвида (Triturus k. Karelinii). В пределах Украины встречается только в горно лесной части Крымского п-ова, от Севастополя на западе до Алушты на востоке, на север — примерно до Симферополя.
Образ жизни. Большую часть периода активности (от начала весеннего пробуждения и до конца сезона размножения — примерно до конца мая — июля) обычно предпочитают временные или постоянные стоячие и слабопроточные водоемы. Так, в окрестностях горы Демерджи в окрестностях Алушты отмечалось несколько особей этого вида в одном небольшом искусственном водоеме диаметром 15–20 м и глубиной до 70–80 см, в другом — до 100 м в диаметре и до 1,5 м глубиной. В Севастопольском районе (окр. с. Терновка) отмечены в искусственном озере площадью 800–1000 м2 на глубине 50–100 см.
По данным учета в этом месте весной 2003 г. численность этих животных на 50–70 метров береговой линии составляла 40–50 особей. В мае 2005 г. в окрестностях с. Лучистое Алуштинского горсовета (окр. горы Демерджи) в двух соседних прудах было отловлено 8 особей тритонов на расстоянии 50–70 м береговой линии и 11 животных были обнаружены неподалеку в двух искусственных ямах размером примерно 30 х 30 см и глубиной около 30 см.
Появления после зимовки зависит от ряда факторов, в том числе от высоты над уровнем моря, специфики погодных условий и может происходить уже в конце февраля. В середине — второй половине марта самцы приобретают брачный наряд и их в это время можно встретить в водоемах. Как правило, такие водоемы приурочены к горно лесной зоне, однако отмечены случаи, когда тритоны в сезон размножения занимали водоемы на участках с ксерофитной растительностью. Примерно до августа тритоны ведут водный образ жизни, проявляя при этом дневную активность, затем покидают водоемы. Известны факты, когда тритоны в конце августа снова возвращались в водоемы после того, как они вновь наполнялись водой после летних дождей. На суше убежищами служат щели под корой упавших деревьев и пней, полости под камнями и пр. Отмечены довольно дальние миграции, когда тритонов находили в 2 км от ближайших водоемов. Примерно в октябре тритоны уходят на зимовку, которая, скорее всего, происходит примерно в таких же убежищах. Не исключено, что тритоны Карелина могут зимовать в водоймах. Существует предположение о том, что эти амфибии, по сравнению с другими представителями данной группы являються наиболее водным видом.
По данным специальных исследований, проведенных в Крыму (Щербак, 1966), в составе питания наибольшую роль играют беспозвоночные, в той или иной степени связанные с водоемами: водяные ослики (более 50%), брюхоногие моллюски (ок. 17%), личинки плавунцов (более 12%) и др. Остальные группы присутствовали в незначительном числе (дождевые черви, бокоплавы, ручейники, паукообразные и др.). Личинки чаще всего питаются мелкими водными ракообразными (циклопы, дафнии), личинками двукрылых, моллюсками.
Состояние популяции. Сделано заключение о необходимости внесения этого вида в третье издание Красной книги Украины (категория «уязвимые виды»). Вид внесен в списки конвенции об охране дикой фауны и флоры и естественной среды обитания в Европе (Берн, 1979; приложение II — «виды, подлежащие особой охране»).
Источники:
- Писанец Е. М. Амфибии Украины (справочник определитель земноводных Украины и сопредельных территорий). — Киев: Зоологический музей ННПМ НАН Украины, 2007