Починаючи з античності, термін «монада» (з грецької monos ‒ одиниця, єдине) слугує для позначення основоположного і неподільного елементу. Френсіс Тейлор в 1982 році використав цей термін в загальноцитологічному сенсі, щоб показати відмінність між «монадами» (власне клітинами) і «химерами» (ендосимбіотичними системами). Клітина-монада є елементарною одиницею життя і характеризується набором структур, які необхідні в онтогенезі для розмноження:
- Цитоплазаматична мембрана, або мембранний бар’єр, що оточує цитоплазматичний компартмент;
- Геном або індивідуальна кодуюча система на основі компактизованої дволанцюгової ДНК;
- Популяція рибосом, або універсальних органел трансляції.
Всі вище перераховані особливості характерні для мітохондрій, ендосимбіотичних органел бактеріального походження, які виконують роль «енергетичних станцій» клітини. Ендосимбіотичні системи являють собою принципово інший тип міжклітинної асоціації. В такому випадку прокаріотна монада поселяється всередині еукаріотної, в результаті вони стають на стільки взаємоповязаними, що існування їх окремо неможливе. Саме таким шляхом в ході еволюції виникли мітохондрії (потомки хемотрофних прокаріотних монад) (рис.1). Згідно цієї гіпотези, одні древні клітини поглинули інші, але не стали їх перетравлювати. Поглинені клітини отримали постійне місце проживання і стабільні умови існування, однак, натомість, повинні були забезпечувати клітину господаря необхідною енергією. В результаті вивчення послідовності основ в мтДНК були отримані докази, що мітохондрії ‒ це нащадки аеробних бактерій, родинних рикетсіям, що колись в предковой еукаріотичній клітині «навчилися» жити в якості симбіонтів. Тепер мітохондрії є майже у всіх еукаріотичних клітинах, розмножуватися поза клітини вони вже не здатні. Спочатку ендосимбіотичні предки мітохондрій не могли ні імпортувати білки, ні експортувати АТФ. Ймовірно, вони отримували від клітини-хазяїна піруват, а вигода для господаря полягала в знешкодженні аеробними симбіонтами токсичного для нуклеоцитоплазми кисню [1,6].
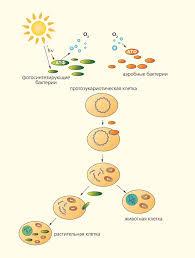
Рис.1 Симбіотична теорія походження мітохондрій (і пластид) (по Margulis L., 1975)
Але, як виявилось в травні 2016 року, мітохондрії і будь-які мітохондріальні гени, які повязані з їх роботою не є обовязковими компонентами еукаріотичної клітини. Вченим із Чехії та Канади вперше вдалося виявити і дослідити еукаріотичний організм, який вторинно втратив мітохондрії і всі гени, необхідні для їх функціонування. Даний організм використовує гени, запозичені у бактерій в ході горизонтального перенесення генів (а це вже демонструє, що еукаріоти не такі вже й недоступні для даного процесу як вважлося раніше), які дозволяють отримувати енергію за рахунок окиснення речовин в цитоплазмі. Мова йде про представника оксимонад ‒ Monocercomonoides globus. Результати дослідження опубліковані в Current Biology.
Monocercomonoides ‒ рід джгутикових паразитичних найпростіших, що належать до роду Oxymonadida. Види Monocercomonoides були знайдені в кишківнику дрібних ссавців, змій і комах і описані як перші еукаріотичні організми, що втратили мітохондрії при переході на безкисневе дихання та паразитичний спосіб життя. Мають овальну або грушоподібну фору, довжину ‒ 5–15 мкм, 4 джгутики, зібрані у пари по два. Живляться шляхом піноцитозу. Загалом геном Monocercomonoides має розмір 75 Mb і включає 16629 білок-кодуючих генів. Геном не містить мітохондріальних (mtDNA) або будь-яких генів для синтезу кардіоліпіну. Представник Monocercomonoides sp. PA 203, який був виділений з кишківника шиншили, запозичив шляхом горизонтального перенесення генів у бактерій гени, що відповідають за систему цитозольної мобілізації сірки, необхідної для побудови залізо-сірчастих кластерів (Рис.2а,b) [2] .

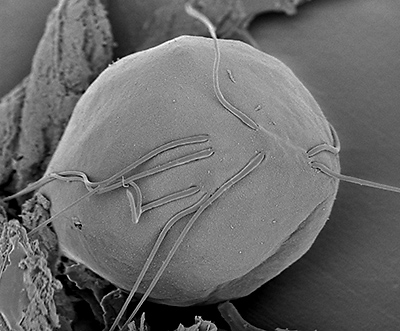
Рис.2а Monocercomonoides (джерело) Рис.2b Monocercomonoides globus — 15 мкм (джерело)
Відомо, що мітохондрії безпосередньо відіграли роль в еволюції еукаріот, тому слід ще раз переглянути археозойну і симбіотичну теорії походження мітохондрій [3].
У 1980-х існувало безліч претендентів на звання сучасного архезоя, але в наступні роки у всіх них знайшли органели, подібні мітохондріям (мітосоми і пероксисоми) і гени-маркери мітохондріального минулого: гени складання Fe-S-білків, мітохондріальних транспортерів і шаперонів, синтетази кардіоліпіну.
Геном Monocercomonoides sp. містить 16629 генів, звичайного комплексу генів, пов'язаного з мітохондріями у еукаріотів (зокрема, генів ферментів, що здійснюють транспорт і сортування пептидів через мітохондріальні мембрани, так званих «механізмів сортування і складання» (sorting and assembly machinery)) не було виявлено. Також відсутні гени мембранних транспортерів АТФ, які є в метамонад. У них вони перекачують АТФ з гідрогеносом і мітосом назовні. Не було генів і білків, що забезпечують контакт ендоплазматичного ретикулуму з мітохондріями або їх похідними.
Яким же чином Monocercomonoides sp. отримують енергію, якщо в них відсутні мітохондрії? У них є гени ферменти, запозичені від бактерій, що відповідають за обмін речовин. Енергію вони отримують за рахунок анаеробного гліколізу ‒ розкладання глюкози в анаеробних умовах, далі пірувату до водню або етанолу і оцтової кислоти . Також вони мають і ще один, більш ефективний спосіб акумуляції енергії ‒розщеплення амінокислоти аргініну безпосередньо в цитоплазмі (аналогічно лямбліям і трихомонодам).
Для отримання енергії важливу роль відіграють провідники електронів, функцію яких виконують залізо-сірчасті кластери (Fe—S-кластери). Вони можуть приймати участь в перенесенні електронів за рахунок рухливих зв’язків. У еукаріотів ці кластери локалізуються в мітохондріях або пластидах. Для синтезу енергії Monocercomonoides sp. використовують бактеріальний набір ферментів, як і близький родич Paratrimastix pyriformis (рис.3).

Рис.3 Синтез енергії у організмів (джерело)
Вгорі: еукаріотичний організм, в якому функціонує комплекс ферментів зборки Fe—S- кластерів. Всередині: анаеробні еукаріотичні організми з редукованими мітохондріями (гідрогеносомами, мітосомами), в яких відсутнєі окисне фосфорилювання продуктів гліколізу, синтезуються Fe—S- кластери. Внизу: заміна мітохондріального комплексу ISC на бактеріальний SUF (Sulfur mobilization) у безмітохондріальних організмів, який відповідає за продукцію залізо-сірчастих кластерів.
Як же еволюціонували мітохондрії метамонад? (рис.4) Мітохондрії можуть редукуватися до мітосом або інших подібних органел, але при цьому функціонують компоненти зборки Fe—S-кластерів (ISC). Предки Monocercomonoides і Paratrimastix отримують бактеріальний комплекс для синтезу Fe—S-кластерів (SUF), що дозволяє їм відкинути ферменти ISC-комплексу. У Monocercomonoides зникають і мітосоми. Паралельно в інших метамонад, не отримали додатковий цитоплазматичний комплекс для синтезу Fe—S-кластерів, залишаються мітосоми або їх аналоги з початковим мітохондріальним комплексом ISC.

Рис.4 Еволюція мітохондрій метамонад (джерело)
Автори відкриття вважають, що Monocercomonoides в своєму еволюційному минулому мали мітохондрії. Доказом цього є наявність у близькородинного виду Paratrimastix pyriformis мітосом. Можливо, у цих найпростіших ще не виявлені мітосоми, які деградували настільки, що в геномі не залишилося будь-яких ознак їх присутності. А успішно захоплені бактеріальні гени дають їм можливість анаеробному, або в середовищі з низькою концентрацією кисню за наявності високих концентрацій органічних речовин, заліза, сірки [4].
Література
- А.В. Пиневич. Микробиология. Биология прокариотов: Учебник в 3 т. Том 1.‒ СПб.: Изд. С.-Петепрб. ун-та. ‒2006. ‒ 352с.
- http://eol.org/data_objects/27476785
- http://biomolecula.ru/content/1955
- Karnkowska et al. A Eukaryote without a Mitochondrial Organelle // Current Biology. – 2016. – 1-11 р.
- http://elementy.ru/novosti_nauki/432755/
- И.О. Мазунин, Володько Н.В. Митохондрии: жизнь в клетке и ее последствия // Природа. № 10. – 201. – 3с.