Meleshko O. V., Korshunov O. V., Shabanov D. A. The study of three hemiclonal population systems Pelophylax esculentus complex from the Seversko-Donetskiy center of green frogs diversity // Вісник Харківського національного університету імені В. Н. Каразіна. Серія «Біологія». – 2014. – Вип.20, №1100. — С. 153-158.
УДК 597.851(477.54)
The study of three hemiclonal population systems Pelophylax esculentus complex from the Seversko-Donetskiy center of green frogs’ diversity
O. V. Meleshko, O. V. Korshunov, D. A. Shabanov
V.N. Karazin Kharkiv National University
meleshko.e.v@gmail.com
The structure of hemiclonal population systems (HPS) Pelophylax esculentus complex from the Iskov pond (Gaydary village, Zmiyiv District), the Lower Dobritskiy pond (the region of Gomolsha river floodplain, Dobritskiy yar, the territory of the National Park “Gomolshanski lisy”) and from the pond near the village Zhovtneve (Vovchanskiy District, Kharkiv Region) was studied. All studied HPS were defined as REt-type. We observed the triploid hybrids number increasing in the HPS from the Iskov pond. The large part of triploid hybrids was found in the HPS of the Lower Dobritskiy pond. It was observed that all triploid hybrids from the pond near the village Zhovtneve were females. There was significant bond between the diploid erythrocytes size and sex of the frogs. It seems that triploids in this HPS are originating in other way than compare to other HPS. There were two groups of females differs in body size which possibly corresponds to different ontogenetic strategies of green frog females. The males have a normal size distribution.
Key words: hemiclonal population systems, Seversko-Donetskiy center of green frogs diversity, Pelophylax esculentus complex, species composition, triploid hybrids, size distribution.
Дослідження трьох популяційних систем Pelophylax esculentus complex Сіверсько-Донецького центру різноманіття зелених жаб
О. В. Мелешко. О. В. Коршунов, Д. А. Шабанов
Харківський національний університет імені В. Н. Каразіна
meleshko.e.v@gmail.com
Вивчена структура геміклональних популяційних систем (ГПС) Pelophylax esculentus complex, що мешкають в Іськовому ставку (с. Гайдари, Зміївський район), Нижньому Добрицькому ставку (район заплави р. Гомольша, Добрицький яр, територія НПП «Гомільшанські ліси» у тому ж районі) та у ставку сел. Жовтневе (Вовчанський район Харківської області). Всі вивчені ГПС відносяться до RЕt-типу. В ГПС Іськового ставу в 2013 р. зареєстровано зростання частки триплоїдів порівняно з 2012 р. В ГПС Нижнього Добрицького ставку знайдена велика кількість триплоїдних гібридів. В ГПС сел. Жовтневе триплоїди представлені тільки самками. Серед диплоїдів цього ставка зареєстрований значущий зв'язок розмірів еритроцитів із статтю жаб. Ймовірно, механізм відтворення триплоїдів в ГПС сел. Жовтневе відрізняється від такого двох інших систем. Розмірний розподіл демонструє наявність двох груп самок, яке вірогідно відповідає двом онтогенетичним стратегіям. Розподіл самців має нормальний характер.
Ключові слова: геміклональні популяційні системи, Сіверсько-Донецький центр різноманіття, Pelophylax esculentus complex, видовий склад, триплоїдні гібриди, розмірний розподіл.
Исследование трех популяционных систем Pelophylax esculentus complex Северско-Донецкого центра разнообразия зеленых лягушек
Е. В. Мелешко, А.В. Коршунов, Д. А. Шабанов
Харьковский национальный университет имени В. Н. Каразина
meleshko.e.v@gmail.com
Изучена структура гемиклональных популяционных систем (ГПС) Pelophylax esculentus complex, обитающих в Иськовом пруду (с. Гайдары, Змиевской район), Нижнем Добрицком пруду (район поймы р. Гомольша, Добрицкий яр, территория Национального природного заповедника «Гомольшанские леса») и в пруду пос. Жовтневое (Волчанский р-н Харьковской области). Все исследованные ГПС относятся к RЕt-типу. В ГПС Иськова пруда зарегистрировано увеличение количества триплоидных гибридов в 2013 г. по сравнению с 2012 г. В ГПС Нижнего Добрицкого пруда обнаружено высокое содержание триплоидных гибридов. В ГПС пруда пос. Жовтневое триплоиды представлены только самками. Для диплоидов в этом пруду показана значимая связь размеров эритроцитов с полом лягушек. Видимо, возникновение триплоидов в данной ГПС связано с иным механизмом, чем в двух других изученных ГПС. В размерном распределении всех лягушек показано наличие двух групп самок, которое, возможно, соответствует различным онтогенетическим стратегиям. Распределение самцов носит нормальный характер.
Ключевые слова: гемиклональные популяционные системы, Северско-Донецкий центр разнообразия, Pelophylax esculentus complex, видовой состав, триплоидные гибриды, размерное распределение.
Introduction
There are three main forms of European green frogs (Pelophylax esculentus complex): the pool frog, Pelophylax lessonae (Camerano, 1882), the lake frog, Pelophylax ridibundus (Pallas, 1771), and the edible frog, Pelophylax esculentus (Linnaeus, 1758). In 1964, L. Berger discovered the green frogs’ phenomenon of interspecific hybridization (Berger, 1964) and found that P. esculentus is interspecies hybrid. The main distinctive feature of green frogs is a hemiclonal inheritance comprising the clonally transfer of one of the parental genomes to gametes, i.e. without recombination. In the case of crossing between hybrids frogs producing gametes of the same parental species, offspring are usually unviable. This phenomenon is called hybridolysis (Plötner, 2005).
Apart from that, different forms of the green frogs from Pelophylax esculentus complex are able to live together and breed in the same area despite of their ecological specificity. This property determines the existence of various types hemiclonal population systems (HPS), which are differs in the presence and dominance of the one or another form, as well as the nature of the gametes produced by hybrid forms (Shabanov et al., 2009; Shabanov, Litvinchuk, 2010).
There are several basic types of HPS (Uzzell, Berger, 1975):
L-type — ordinary populations of P. lessonae, the pool frog;
R-type — ordinary populations of P. ridibundus, the lake frog;
E-type — so called «pure» populations of hybrids, P. esculentus;
LR-type — population systems of P. lessonae and P. ridibundus; breeding produces no hybrids;
LE-type — population systems of P. lessonae and P. esculentus;
RE-type — population systems of P. ridibundus and P. esculentus;
LER-type — mixed systems, comprising both parental species and their hybrids.
It is common to indicate the presence of triploids in HPS by adding the letter “t” to mark the population system. For example, REt-type systems compose of P. ridibundus, as well as di- and triploid P. esculentus; Et-type systems consist of diploid and triploid hybrids only.
The features of Seversko-Donetskiy center of green frogs’ diversity. The region of the HPS’ high diversity is associated with the Seversky Donets river basin within Kharkiv and Donetsk Regions. Moreover, polyploid hybrids are often found in this region (Borkin et al., 2004), called Seversko-Donetskiy center of green frogs’ diversity (Shabanov et al., 2009). The population systems of R, RE, REt and Et-types with P. esculentus of both sexes have been found in the Seversky Donets river valley. Coexisting di-and triploid hybrids are typical for Seversky Donets floodplain and adjacent habitats (Korshunov, 2010).
Seversko-Donetskiy center of green frogs’ diversity is defined by the following: (i) P. ridibundus and various forms of P. esculentus inhabit the center; mature P. lessonae are absent; individual representatives of the parental species have been recorded only at the stage of fingerlings (froglets before the first hibernation). Subsequently all genomes of the P. lessonae are transmitted via diploid and triploid hybrids. (ii) Diploids of the both sexes are numerous among P. esculentus in the diversity center, as well as a significant number of triploids of the both sexes: LRR (more numerous) and LLR, and also single immature tetraploids LLRR were found. (iii) Both sexes’ specimens, producing either P. lessonae gametes or P. ridibundus gametes, or both P. lessonae and P. ridibundus gametes were observed among diploid P. esculentus. (iv) Genetic diversity of P. lessonae genomes transmitted via hybrid frogs in this center is not lower than that of F1 hybrids of those regions inhabited by both parental species (Mezhzherin et al., 2010). (v) The recombinant specimens have been registered among green frogs from this center i.e. P. ridibundus genome has fragments of the P. lessonae genome (Mezhzherin et al., 2005).
The history of previous researches
The most long lasting studies of the HPS structure was undertaken at the Iskov pond (hereinafter – IpHPS), situated near the biological station of V.N. Karazin Kharkiv National University (Gaidary village, Zmiyiv District, Kharkiv Region). Initially the E-type system was described there (Lada, 1995). This HPS is represented by almost exclusively diploid specimens of the P. esculentus of both sexes. However, because the pond was hauled down, significant change of the HPS structure occurs. After the pond restoring, spawning areas almost without females have been observed for a long time. Tadpoles haven’t been recorded in the pond since 2004 till 2011, when tadpoles and then fingerlings were observed in large quantities. Further observations have led us to conclusion that the frog reproduction in the pond is restoring. Suspected mechanisms of the HPS reproduction, considering females coming from other spawning habitats as a source of new clonal genomes, have been described (Meleshko et al., 2011). This allows us to assume that the dynamics of the IpHPS structure is of considerable interest to study.
HPS of Lower Dobritskiy Pond (hereinafter – DpHPS, near the Gomolsha river valley, the territory of National Park “Gomolshanski lisy”) was a subject of primary interest. The single specimen of the hybrid tetraploid frog was recorded close to the habitat of this HPS (Borkin et al., 2004). In the top of that, high proportion of triploid hybrids was recorded for this HPS.
The HPS of the Zhovtneve (hereinafter – ZhpHPS, Vovchanskiy District, Kharkiv Region) is located far away from the two latter. The high proportion of triploid hybrids is typical for the green frogs from this HPS.
In all HPS which was in focus of our studies, according to the data of Yu. Rozanov and S. Litvinchuk, RL genotypes were recorded. In addition to that, RRL triploid form was recorded from the IpHPS, and RL – in the DpHPS and ZhpHPS. Their numbers had not been estimated earlier, because separate specimens of particular interest were collected instead of the representative samples. The genotypes and ploidy level of specimens were determined by DNA flow cytometry. The details of the technique have been published previously (Borkin et al., 2001).
The study of these three HPS can provide valuable information for understanding the reproduction processes within Pelophylax esculentus complex in the Seversko-Donetskiy center of green frogs’ diversity.
Materials and methods
The research was undertaken in June-July, 2012-2013. The mature green frog individuals were collected from Iskov Pond (Gaydary village, Zmiyiv District), Lower Dobritskiy Pond (near the Gomolsha river valley, Dobritskiy tract, territory of the National Park “Gomolshanski lisy”) and from the pond in the Zhovtneve village (Vovchanskiy District, Kharkiv Region). Collection sites and the information about the samples are composed in Table. 1.
Table 1. The studied samples of green frogs
|
Locality |
Geographic coordinates |
2012, number |
2013, number |
Total number
|
|
|
Iskov pond |
N 49˚ 37′ 35″ E 36˚ 17′ 07″ |
161 |
93 |
254 |
433 |
|
Lower Dobritskiy Pond |
N 49˚ 33′ 22″ E 36˚ 18′ 39″ |
25 |
76 |
101 |
|
|
pond in the village Zhovtneve |
N 50˚ 08′ 04″ E 36˚ 46′ 01″ |
43 |
35 |
78 |
|
Field work was carried out using standard techniques. Animals were caught at night with a light tube or in the daytime manually or with a hand net. Captured frogs were placed in wet canvas or mesh bags or in plastic boxes. In some cases, during the temporary handling of the animals in the laboratory, they were placed in cans or plastic bottles, which lay on its side without water, once a day frogs were washed with running water.
Determination of the species and sex was carried out visually based on complex of the morphological characters (Korshunov, 2010; Lada, 1995). It is known that visual determination of green frogs based on morphological features does not allow making a precise assumption on their species status. In particular, the triploid hybrid individuals are harder to distinguish visually from individuals of the parental species due to the effect of gene dosage. However, determination of ploidy level by measuring the average length of erythrocytes, the results of which are regularly confirmed by karyological research, significantly reduces the probability errors in the determination.
All specimens from the ZhpHPS aliquot, 99 individuals from the DpHPS aliquot and 183 individuals from the IpHPS aliquot were analysed cytologically, i.e. erythrocytes size was determined. The air dried blood smears of each specimen were made according to the standard technique: a drop of blood obtained from a frog finger was coated on a slide glass and smeared with a thin layer of the second slide edge (Bondarieva et al., 2012). Smears were dried up and photographed under high magnification (13 of a light microscope using a USB-eyepieces camera. The object-micrometer was photographed under the same conditions. Major axis length measurement of 15-20 erythrocytes was performed on photos using the PDF XChange Viewer software. All measurements are given in µ. Sex of adults, determined by dissection, and the age composition (juveniles or adults) of samples are composed in Table 1.
Results and discussion
The composition of the samples studied is represented in Tables 2, 3. Only data for specimens with cytologically determined ploidy level are given in Table 2, Table 3 includes all sample data. The diploid specimens of the P. esculentus dominated in number green frogs taxocenes of the IpHPS (Table 2). It should be noted that we have registered a significant (p <0.02) increasing in the proportion of triploid P. esculentus in 2013 in comparison with 2012.
Table 2. Determination of ploidy of individuals from studied HPS
|
Sample |
Year |
P. ridibundus 2 n, number (%) |
P. esculentus, 2n, number (%) |
P. esculentus, 3 n, number (%) |
Total number |
|
Iskov pond |
2012 |
3 (3%) |
96 (96%) |
1 (1%) |
100 |
|
2013 |
8 (10%) |
68 (82%) |
7 (8%) |
83 |
|
|
Lower Dobritskiy Pond |
2012 |
1 (4%) |
19 (83%) |
3 (13%) |
23 |
|
2013 |
9 (12%) |
47 (62%) |
20 (26%) |
76 |
|
|
Pond in the village Zhovtneve |
2012 |
3 (7%) |
28 (65%) |
12 (28%) |
43 |
|
2013 |
7 (20%) |
18 (51%) |
10 (29%) |
35 |
|
|
Total number (%) |
31 (8.5%) |
276 (76.5%) |
53 (15%) |
360 |
|
Previously it was assumed that the IpHPS crisis was associated with the loss of all clonal genomes except the male P. lessonae genome (Shabanov et al., 2006). The resumption of frog reproduction in the pond during the past years could potentially indicate that the HPS got new clonal genomes. We can only assume what kind of genomes was brought in the pond and by which specimens. It is most likely that the observed change of triploid hybrids proportion in 2013 is also a consequence of the ongoing transformations caused by penetrating of new clonal genomes into HPS. We assume that HPS state is premature in this case, and now it is going to the populational equilibrium.
Table 3. Sex composition of the studied HPS
|
Sample |
Year |
Females, number (%) |
Males, |
Total, number |
|
Iskov pond |
2012 |
34 (21) |
127 (79) |
161 |
|
2013 |
13 (14) |
80 (86) |
93 |
|
|
Lower Dobritskiy Pond |
2012 |
- |
25 (100) |
25 |
|
2013 |
30 (40) |
46 (60) |
76 |
|
|
Pond in the village Zhovtneve |
2012 |
18 (42) |
25 (58) |
43 |
|
2013 |
17 (49) |
18 (51) |
35 |
|
|
Total number (%) |
112 (26) |
321 (74) |
433 |
|
DpHPS comprised mainly by the diploid of the P. esculentus (Table 2). A lot of triploid hybrids were recorded there as well as in the previous studies (Borkin et al., 2004). The difference in the proportion of triploids in the samples 2012 compared to samples 2013 was not significant. Notably in 2012 exclusively males occurs in the sample from this HPS. In 2013, the proportion of females was about 40% (Table 3). Apparently, this difference is not associated with a change in the composition of HPS and with distinction in time of the samples collecting. In 2012, the frogs have been caught during the spawn by capturing the males in the spawning areas, but in 2013 the collections was done after spawning.
The results of the ZhpHPS study are of particular interest. Diploids of the edible frogs also dominate there. P. ridibundus makes up to 7% of the frogs total number in the sample in 2012 and 20% in the sample in 2013 (Table 2), but such difference in the sample composition is statistically insignificant. Triploids of P. esculentus accounted for about 30% of the studied samples and represented exclusively by females (group I, Fig. 1). At the same time, triploid hybrids are represented by both sexes in the two previously described HPS. This distinction indicates that a mechanism of triploids' reproduction in the ZhpHPS is different.
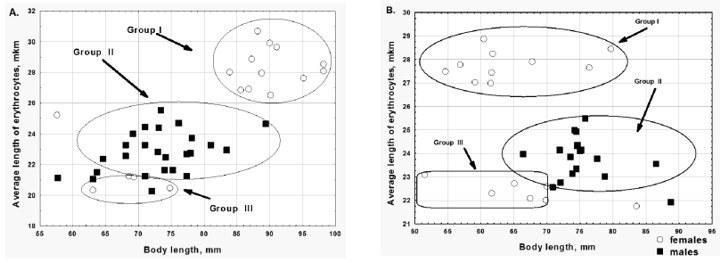
Fig. 1. The length of erythrocytes dependence on body length for green frogs from pond in the village Zhovtneve. A – sample 2012; B – sample 2013
The second interesting feature of the ZhpHPS was a significant difference in the size of red blood cells of diploid males (group II, Fig. 1) and diploid females (group III, Fig. 1): size of diploid males' erythrocytes was significantly higher. Once it was assumed that there were both di-and triploid individuals among males. This assumption was tested using karyoanalysis and proved to be wrong (O. Mikhailova, personal communication). We should state that samples from 2013 consists the females with a much smaller body size than in 2012. This is probably due to the fact that in 2012 collection of the material was done during spawning, and in 2013 - after it.
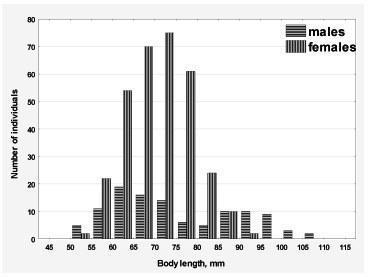
Fig. 2. Distribution of body size of green frogs from the samplings 2012-2013 years
In addition, we examined the size distribution in the all frogs HPS. The pattern of distribution was found to be similar for all the studied HPS, so all studied samples were considered in Fig. 2 together. Distribution of the female size has two peaks corresponding to the relatively small and larger individuals. This result can be considered as an indirect confirmation of the assumption (Usova, 2010) about two growth strategies specific to female of the green frogs: precocity (high fertility with the rapid growth and short life expectancy) and slow-growth (low growth rate and a relatively high life expectancy).
Thus, all studied HPS were REt-type; each of them was characterized by a particular structure. DpHPS characterized by high percentage of triploid hybrids. Restoration of reproduction is observed in IpHPS, passed through a critical condition. All triploid individuals were females in ZhpHPS. Pattern of the size distribution was similar in all three HPS and characterized by the separation of females at a relatively small and relatively large individuals.
Acknowledgements
The author takes this opportunity to thank M. Kravchenko, O. Mikhailova, A. Bondareva, O. Usova, and M. Stroilov for their invaluable research assistance. The author also thanks the students of V.N. Karazin Kharkiv National University involved in fieldwork: O. Klempert, A. Us, Yu. Kravchenko, S. Ogienko, N. Kozak, Yu. Gladkova, M. Kulymova, E. Malchenko, A. Ptushko, Yu. Baran, E. Beshevets, A. Klysa, D. Krugovoj, A. Mikhaylenko, A. Laptijy, S. Zyubrijy, Yu. Kostyuk, A. Miroshnichenko, and R. Mironov.
Literature cited
Berger L. Is Rana esculenta lessonae Camerano a distinct species? // Ann. Zool. PAN. – 1964. – Vol.22, №13. – P. 245–261.
Bondarieva A. A., Bibik Yu. S., Samilo S. M., Shabanov D. A. Erythrocytes cytogenetic characteristics of green frogs from Siversky Donets centre of Pelophylax esculentus complex diversity // Vistnyk of V.N. Karazin University. Series: biology. – Kharkiv, 2012. – Vol. 15, № 1008. – P 116–123.
Borkin L. J., Korshunov A. V., Lada G. A., Litvinchuk S. N., Rosanov J. M., Shabanov D. A., Zinenko A. I. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian Journal of Herpetology, 2004. – Vol. 11, No 3. – P. 194–213.
Borkin L. J., Litvinchuk S. N., Rosanov J. M., and Milto K. D. Cryptic speciation in Pelobates fuscus (Anura, Pelobatidae): evidence from DNA flow cytometry. // Amphibia–Reptilia. – 2001. Vol. 22(4). P. – 387 – 396.
Korshunov O. V. Ecological patterns of distribution Pelophylax esculentus complex habitats in the basin upstream of the Seversky Donets. Author. dis.... Candidate. biol. Sciences: 03.00.16 "ecology" // Dnepropetrovsk, 2010. – 24 p.
Lada G. A. “Middle European green frogs (the hybridogenetic Rana esculenta complex): an introduction to the problem,” in: V. M. Kolontaev (ed.), Flora and Fauna of the Chernozem Territory, Izd. TGU, Tambov, 1995. – pp. 88 – 109.
Meleshko E.V., Suvorova A. D., Kravchenko M. A., Shabanov D. A. The population system of green frogs (Pelophylax esculentus complex) of Iskov pond in Zmievsky area of the Kharkiv region: the history of researching and number estimation // Biologicheskiy vestnik. – Kharkiv, 2012. – Vol. 14, № 1, 2010. – P 63–68.
Mezhzherin S. V., Morozov-Leonov S. Y., Nekrasov O. D., Kurtyak F. F., Jalaj E. I. Spatial structure of the hybrid complex of green frogs Rana esculenta (Anura, Ranidae) in Ukraine // Materіal of First Ukranian Herpetological Community Conference, Kyiv. – 2005– P. 10–114.
Mezhzherin S. V., Morozov-Leonov S. Yu., Rostovskaya O. V., Shabanov D. A., Sobolenko L. Yu. The ploidy and genetic structure of hybrid population of water frogs Pelophylax esculentus complex (Amphibia, Ranidae) of Ukraine fauna // Cytology and Genetics. – 2010. – Vol. 44, No 4. – P. 212–216.
Plötner J. Die westpaläarktichen Wasserfrösche. Bielefeld: Laurenti-Verlag, 2005. – 161 P.
Shabanov D. A., Korshunov O. V., Kravchenko M. O. Which of the water frogs inhabit Kharkiv oblast? Perspectives on terminology and nomenclature. V. N. Karazin Kharkiv National University // Biology and valeology. – Vol. 11. – Kharkiv, 2009. – P. 116 – 125.
Shabanov D. A., Lytvynchuk S. N. Green frogs: life without rules or a special way of evolution? // Priroda, 2010. № 3 – P. 29–36.
Shabanov D.A., Zinenko O. I., Korshunov O. V., Kravchenco M. O., Mazepa G. A. The study of population systems of green frogs (Rana ecsulenta complex) in Kharkiv region: history, modern condition and prospects // Vistnyk of V.N. Karazin University. Series: biology. – Kharkiv, 2006. – Vol. 3, № 729.
Usova E. E. Skeletohronological study of representatives Pelophylax esculentus complex longevity and growth dynamics: check differences between fast–growing and long–lived frogs // Biodiversity and sustainable development. Abstracts Intern. scientific and practical. conf. Simferopol, 2010. – P. 121–124.
Uzzell T.M., Berger L. Electrophoretic phenotypes of Rana ridibunda, Rana lessonae and their hybridogenic associate Rana esculenta // Proc. Acad. nat. Sci. Phila. –1975. –Vol. 127. –P. 13–24.