|
|
|||
|
БІОСИСТЕМИ. БІОСФЕРА
ЕКОСИСТЕМИ. ПОПУЛЯЦІЇ
ОРГАНІЗМИ У ДОВКІЛЛІ
ЛЮДСТВО ТА ЙОГО ДОЛЯ
|
|||
|
← IV-18. (доповнення) Стратегії внутрішньопопуляційноі взаємодії |
IV-19. (доповнення) Секс має бути неможливим! Чому ж він існує? |
||

IV-19. (доповнення) Секс має бути неможливим! Чому ж він існує?
(у роботі)
Еволюція сексу є найскладнішою проблемою еволюційної біології.
Джон Мейнард Сміт
Стать — головний виклик сучасної теорії еволюції... королева проблем еволюційної біології.
Грем Белл
Одна з дивних особливостей сексу полягає в тому, що, за загальноприйнятими науковими уявленнями, він є неможливим. Щоб зрозуміти це, перш за все слід розглянути, що саме ми будемо вкладати у поняття «секс» у ході даного обговорення; для цього доведеться розплутати непрості термінологічні складнощі. Розмежуємо три різні розуміння поняття «секс»:
— секс у побутовому сенсі — статевий акт, «заняття сексом»; це — реалізація у звичайному житті загадкового явища, що пов'язане з відтворенням нових генотипів та розмноженням;
— секс sensu stricto (у вузькому розумінні) — стать, приналежність особини до однієї з двох категорій, які відрізняються статевими клітинами (гаметами), які вони можуть продукувати; як відомо, особини жіночої статі продукують відносно великі гамети, яйцеклітини, чоловічої — дрібні сперматозоїди; поняття «секс» у цьому сенсі важливо не плутати з поняттям «гендер», яке стосується культурного конструкта, що описує уявлення про особливості та ролі представників різних статей;
— секс sensu lato (у широкому розумінні) — сексуальна рекомбінація, утворення нових генотипів, що у нових випадкових комбінаціях поєднують генетичну інформацію батьківських особин.
«Під сексом в еукаріотів ми розуміємо більш-менш регулярне чергування мейозу та сингамії. Природним наслідком цього є чергування гаплоїдних і диплоїдних фаз у життєвому циклі».
Джон Мейнард Сміт, Ерс Сатмарі. Основні переходи в еволюції. 1995.
Третє з перелічених способів розуміння поняття «секс», і той сенс, про якій йдеться у наведеній цитаті — те ж саме. «Ядром» комплексу феноменів, у число яких входить і наявність статей, і різниця між ними, і статеві акти, є саме сексуальна рекомбінація, утворення нових комбінацій генів внаслідок поєднання спадкових ознак батьківських особин внаслідок запліднення — злиття їхніх гамет (сингамії). Злиття гамет призводить до збільшення кількості спадкової інформації вдвічі; тому в життєвому циклі, де є запліднення, має бути й двократне скорочення кількості спадкової інформації. Таке скорочення є ймовірнісним, і його забезпечує мейоз. Мітози забезпечують клонову передачу генетичної інформації, мейоз у поєднанні з заплідненням — сексуальну.
Різне розуміння понять «секс» та «сексуальний» є причиною низки непорозумінь і відрізняється у різних наукових традиціях. Порівняємо два визначення, з англійської та української Вікіпедій.
«Parthenogenesis <...> is a natural form of asexual reproduction in which the embryo develops directly from an egg without need for fertilization / Партеногенез <...> — це природна форма безстатевого розмноження (= асексуального відтворення), при якій ембріон розвивається безпосередньо з яйцеклітини без необхідності запліднення» (Wikipedia. The Free Encyclopedia).
«Партеногенез (від грецького παρθενος — незаймана та γενεσις — народження) — форма статевого розмноження, коли розвиток зародка відбувається без запліднення» (Вікіпедія. Вільна енциклопедія).
Де помилка? Помилки нема, є різні традиції використання понять. Українська Вікіпедія у даному питанні наслідує російську; те ж саме у більшості випадків стосується традиції викладання відповідних університетських курсів. Ми вважаємо кращим вибором приєднатися до англо-американської традиції розгляду цього питання.
Є секс формою розмноження чи відтворення? У низці випадків секс не пов'язаний зі збільшенням чисельності особин (власне розмноженням), він є причиною лише утворення нових (відтворення) генотипів. Ми вважаємо доцільним використовувати слово «розмноження», коли мова йде про організми, а при обговоренні процесів, що відбуваються в популяціях (на вищому від організмового рівні) та з генотипами (на нижчому рівні) використовувати «відтворення». Звісно, коли мова йде про відтворення, поняття «sexual / сексуальний» та «asexual / асексуальний, клоновий» використовуються не для опису привабливості чи непривабливості особин, а для зазначення того, як утворюються нові генотипи.
Звісно, парадоксальність сексу має сенс обговорювати саме у разі описаного розуміння цього поняття (втім наявність статей також є парадоксальною!). У разі наявності двох особин різної статі, що мають можливості та бажання здійснити статевий акт, секс (у побутовому сенсі) стає можливим. Але на чому базуються ці можливості та такі бажання? На наявності морфологічних, фізіологічних та поведінкових пристосувань, що забезпечують запліднення, у представників двох різних статей. На існуванні роздільностатевих популяцій, де взаємодіють особини різних статей. На тому, що сексуальна рекомбінація стала та лишається основним засобом утворення нових генотипів у переважної кількості високоорганізованих видів, включно з людиною. Чи є така ситуація єдиною можливою? Звісно, ні; згадаємо про клонові популяції, що обходяться без сексу; про популяції гермафродитів, що обходяться без різних статей; про можливість відтворення людей внаслідок природного клонування — утворення однояйцевих близнюків внаслідок мітотичних поділів заплідненої яйцеклітини.
Ми згадали кілька типів відтворення. Для подальшого обговорення нам потрібно запропонувати їх типологію. Пропонована нами типологія показана в табл. IV-19.1 Її було розроблено (Шабанов, 2013, 2015) для обговорення особливостей геміклонових популяційних систем гібридогенного комплексу зелених жаб, і цим пояснюється увага до IV типу відтворення. Безумовно, найдавнішим типом відтворення популяцій є Ia, клоновий з горизонтальним перенесенням генетичної інформації; цей тип є характерним для прокаріотів. Ймовірно, у більшості випадків представники типу Ib (клонові без горизонтального перенесення) є організмами, що колись мали статеву рекомбінацію, а далі її втратили. До якого типу належали перші організми з сексуальним відтворенням — питання, яке можна обговорювати; можливо, це був тип, що не показаний в табл. IV-19.1 (однотипні організми, що продукували однотипні гамети). В усякому разі, зрозуміло, що виникнення сексуальної рекомбінації не могло бути пов'язаним з типом IIIb; для виникнення суттєвих екологічних відмінностей між статями вже мали існувати окремі статі, тобто популяції IIIb мали розвинутися з популяцій IIIa.
Таблиця IV-19.1. Типи популяційного відтворення та пов’язані з ними особливості біосистем
|
Біосистеми |
Популяції |
Геміклонові популяційні системи |
||||
|
Клонові популяції |
Менделевські популяції |
|||||
|
з горизонтальним перенесенням |
без горизонтального перенесення |
гермафродитні |
роздільностатеві |
|||
|
без статевого диморфізму |
зі статевим диморфізмом |
|||||
|
Ia |
Ib |
II |
IIIa |
IIIb |
IV |
|
 |
 |
 |
 |
 |
 |
|
|
Особини |
Однотипні |
Однотипні, дві статі |
Дві статі, що значно відрізняються |
Дві статі, різні форми |
||
|
Генеалогія |
Лінійна |
Сітчаста |
Фактично лінійна (у диплоїдних гібридів) |
|||
|
Генетична |
Нема; всередині клону особини не унікальні |
Переважна більшість особин генетично унікальна |
Неспадкова (у геміклонів і клонів) |
|||
|
Генофонд |
Нема, набір клонів |
Є; пул генів, що рекомбінують |
Генофонд батьківського виду + набір геміклонів і клонів |
|||
|
«Подвійна |
Нема; потомство можуть безпосередньо продукувати усі особини |
Є; безпосередньо потомство можуть продукувати лише самиці |
||||
|
Статевий |
Нема |
Відносно менш ефективний |
Відносно більш ефективний |
Є |
||
|
Внутрішньовидова |
Негомологічна, іноді — гомологічна |
Нема |
Гомологічна (між відповідними хромосомами та локусами завдяки кросинговеру) |
Є у триплоїдних гібридів між хромосомами батьківського виду, що представлені двома копіями |
||
|
Міжвидова |
Негомологічна |
Нема (внаслідок безпліддя міжвидових гібридів, яка викликана порушеннями впізнавання гомологічними хромосомами одна одною у мейозі) |
Гомеологічна (між відповідними хромосомами різних видів) |
|||
| Існування «справжніх» видів |
Сумнівне | Існують; у типовому випадку види є спільнотами з певним генофондом, що генетично відокремлені від інших видів | Сумнівне | |||
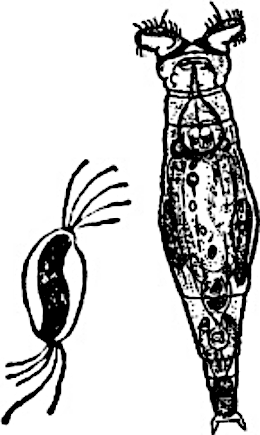
Перш за все, розглянемо у чому полягає диво сексуального відтворення. Через наші смертні тіла тече потенційно безсмертна послідовність клітин герми — зародкової лінії. І тіло, і «Я» кожного з нас утворює сома, тупикова бічна гілка герми (рис. IV-19.1). Успіх соми — це передача герми далі, крок до її безсмертя... У разі сексуальної рекомбінації кожного разу, коли герма проходить через сому, вона відновлюється: кожен з фрагментів ядерної батьківської генетичної інформації з ймовірністю 1/2 втрачається, і з ймовірністю 1/2 переходить до наступного покоління. Це особливо дивно у порівнянні з цитоплазматичною спадковою інформацією: мітохондріальна ДНК спадкується клоново; спадкування Y-хромосоми у людини відбувається без рекомбінації, але з ймовірністю 1/2.
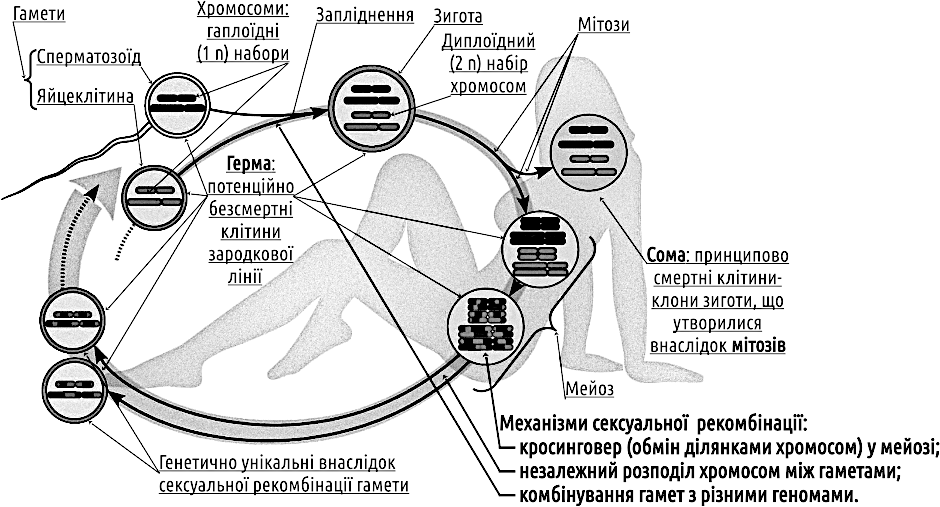
Рис. IV-19.1. Типовий життєвий цикл з сексуальною рекомбінацією, заплідненням, мейозом та диплоїдною і гаплоїдною фазами
Ми зазначили, що існування статевої рекомбінації є парадоксальним. Щоб довести це, розділимо пояснення існування спостережуваних природних феноменів на два етапи: по-перше, пояснення того, як обговорюваний феномен виник, і, по-друге, пояснення того, чому він продовжує існувати, зберігається. Розглянемо існування сексу (сексуальної рекомбінації) та роздільностатевості з цього погляду. У такому разі ми стикнемося з низкою нез'ясовних обставин. Для зручності обговорення зазначимо їх як парадокси, що пов'язані з видатними постатями в історії еволюційної біології.
Парадокс Докінза (як ми його назвемо), стосується появи сексу. Показані на рис. IV-19.1 процеси є результатами узгодженої роботи низки молекулярно-генетичних механізмів. Їх комплекс настільки складний та взаємоузгоджений, що, ймовірно, ми можемо бути впевнені, що він виник один раз в історії життя. Складні адаптації біосистем є наслідком добору. Таким чином, ми бачимо результати тривалого добору, що сформував генетичний комплекс, який забезпечує секс. Як діє добір на гени (та й на біосистеми інших рівнів)? Через підвищення їхньої пристосованості: міри внеску у наступні покоління. Попри існування різних способів вимірювання пристосованості, усі вони спираються на те ж саме: у майбутньому зберігаються такі особливості, які підвищують шанси на збереження та поширення у майбутньому. З цього погляду парадоксальним є те, що перехід від асексуального відтворення до сексуального пов'язаний з двократним зниженням пристосованості усіх генів, що його забезпечують!
Цей парадокс ми пов'язали з ім'ям людини, яка внесла найбільший внесок у поширення ідеї про те, що добір відбувається саме на генному рівні (див. пункт I-18). На рівні організмів (наприклад, у випадку гермафродитів) та популяцій перехід до сексуальної рекомбінації не обов'язково пов'язаний з зниженням пристосованості вдвічі, але на рівні генів цей ефект є очевидним! Навіть не поділяючи погляди Докінза на всемогутність добору на генному рівні, важко зрозуміти, як організмовий чи груповий добір міг сформувати настільки складні генетичні пристосування. Для Докінза, який наполягає на тому, що усі складні пристосування пов'язані з генним добором, цей парадокс особливо болісний.
«Коли відтворення асексуальне, ставки є рівними та спільними, оскільки всі реплікатори мають однакові 100-відсоткові шанси опинитися в кожному потомку, утвореному їхніми спільними зусиллями. При сексуальному відтворенні відповідний шанс для кожного реплікатора — лише половина від повного, але ритуалізована ввічливість мейозу, «гавот» Гамільтона, значною мірою гарантує кожному алелю рівні шанси пожинати плоди успіху спільного репродуктивного підприємства. Це, звичайно, питання: чому «гавот» хромосом такий ввічливий. Це надзвичайно важливе питання, від якого я ухилюся з простого посилання на боягузтво. Це одне з низки питань про еволюцію генетичних систем, з якими більш-менш марно боролися кращі уми, ніж мій (Williams 1975, 1980; Maynard Smith 1978a), низка питань, які спонукали Вільямса зауважити, що «в еволюційній біології назріває свого роду криза». Я не розумію, чому мейоз відбувається саме так, але з огляду на те, що він відбувається, багато чого випливає. Зокрема, організований чесний процес мейозу допомагає пояснити узгодженість і гармонію, яка об'єднує частини індивідуального організму. Якби на рівні групи особин як потенційного носія привілей на розмноження надавався з такою ж скрупульозною чесністю в такому ж добре дисциплінованому «гавоті організмів», груповий добір міг би стати більш правдоподібною теорією еволюції».
Річард Докінз. Розширений фенотип: Далекосяжність гена, 1999.
На наш погляд, пояснення появи сексу можливо лише у разі, якщо ми передбачатимемо велику роль в цьому процесі групового добору. Як зазначає Мейнард Сміт, ідея, що поява сексу може бути наслідком групового добору, належить самому Рональду Фішеру — творцю генетичної теорії еволюції. Визнавати ключову роль групового добору Докінз не хоче; хоча в «Розширеному фенотипі» він визнає принципову можливість групового добору, але наполягає на тому, що жодні складні адаптації з'явитися не можуть. Саме тому Докінз у якості кращого рішення обирає розписатися у своєму незнанні. Чому мова йде про груповий добір? Тому що ми маємо конструювати аргументи на кшталт таких: «хоча представники тієї філогенетичної лінії, яка перейшла до сексу, відтворювалися менш ефективно, ніж представники споріднених з нею клонових ліній, ймовірність зникнення представників цієї лінії внаслідок [обрати потрібне: непрогнозованої зміни умов; еволюції паразитів; деградації геномів тощо] є нижчою». Таким чином, ми розглядаємо не успіх окремого організму чи пріоритетне поширення певних генів, а успіх або неуспіх певних груп організмів.
Скоріше за все, виникнення сексу було унікальною подією в історії життя. Ймовірно, поява мейозу, запліднення, клітинного ядра, характерної для еукаріот організації геному — це комплекс новацій, що були наслідком походження еукаріот внаслідок симбіогенезу між археями та альфапротеобактеріями (які стали мітохондріями). Згідно з гіпотезою Вільяма Мартина та Євгена Куніна, що запропонована у 2006 році, зміни, що відбулися під час становлення еукаріот, були пов'язані з необхідністю захисту геному клітини-хазяїна від мобільних генетичних елементів (ретротранспозонів), які приніс з собою симбіонт; ці мобільні елементи перетворилися на інтрони II групи. Згідно з цією гіпотезою, в ході швидких еволюційних перебудов, викликаних необхідністю захисту геному хазяїна, відбулася перебудова прокаріотичної системи генетичної рекомбінації та виник мейоз. У разі таких подій важливо не те, як змінюється пристосованість «генів статевого розмноження» (при усій віртуальності уявлень про такі гени), а те, чи зберігаються організми, що поєднують переваги архейного та бактеріального метаболізму, чи зникають внаслідок деструктивних процесів у геномі. Ця гіпотеза пов'язує появу сексу не з добором на рівні генів, а з груповим добором.
Досліджувати унікальні події, що відбулися у важкореконструйованих умовах, — складна задача. Але навіть якщо ми не зможемо пояснити, як виник секс, ми будемо примушені визнати, що він існує. Ми бачимо численні приклади відмови від сексу у ході еволюції. Якщо деякі групи відмовляються від сексу, чому це не роблять усі? Стійкість сексу простіше досліджувати, ніж його походження, адже забезпечення стійкості сексу відбувається безпосередньо зараз у величезної кількості сучасних груп.
Парадокс Дженкіна, як ми його розуміємо, стосується стійкості сексуальної рекомбінації попри те, що вона раз за разом руйнує вдалі сполучення спадкових ознак. Слід зазначити, що так званий «кошмар Дженкіна» (так у приватному листі Чарльз Дарвін назвав аргумент Флемінга Дженкіна, шотландського професора інженерії, що відомий як винахідник канатної дороги та підводного телеграфного кабелю) зазвичай розуміють неправильно. Щоб детально розібратися в цьому, процитуємо першоджерело.
«Уявімо, що білий чоловік зазнав кораблетрощі на острові, населеному неграми <...> Уявімо, що він володіє фізичною силою, енергією та здібностями домінуючої білої раси <...> надамо йому всі переваги, які ми можемо собі уявити для білого над тубільцем <...> але з усіх цих допущень не випливає висновок, що через обмежену чи необмежену кількість поколінь мешканці острова будуть білими. Наш герой, що зазнав корабельної аварії, ймовірно, стане королем; він уб'є багато негрів у боротьбі за існування; у нього буде багато дружин і дітей, тоді як багато його підданих житимуть і помруть холостяками <...> і все ж йому не вистачить жодного покоління, щоб зробити нащадків своїх підданих білими. Можна сказати, що білий колір не є причиною переваги. Це правда, але він може бути використаний лише для того, щоб показати, як якості, що належать одній людині у великій кількості, повинні поступово стиратися. У першому поколінні з'явиться кілька десятків розумних молодих мулатів, які за середнім рівнем інтелекту значно переважатимуть негрів. Можна було б очікувати, що трон протягом кількох поколінь посяде більш-менш жовтий король; але чи може хтось повірити, що весь острів поступово заселиться білим, або навіть жовтим населенням, або що остров'яни набудуть енергії, мужності, винахідливості, терпіння, самовладання, витримки, завдяки яким наш герой убив стільки своїх предків і породив стільки дітей; тих якостей, які, власне, відібрала б боротьба за існування, якби вона взагалі щось могла відібрати?».
Флемінг Джексон. Огляд «Походження видів». Північно-Британський огляд, 1867.
Ми бачимо, що аргумент Дженкіна (відповідно до поглядів того часу) є відверто расистським; сперечатися тут з глибоко скомпрометованими уявленнями про можливості представників різних рас нема сенсу. Зрозуміло, чому аргумент Дженкіна намагаються не цитувати напряму і обговорювати на рівні аналогій, наприклад, таких. «Припустимо, на полі білих маків з'явився червоний мак. Він помітніший для запилювачів і тому отримає перевагу перед білими маками. Як не сумно, його потомство буде лише рожевим, у другому поколінні — блідо-рожевим, і незабаром нова ознака зійде нанівець. Лише відкриття Менделя, який довів, що колір маку визначається дискретно успадкованим геном, допомогло розв'язати цю проблему». Але ж у разі використання подібних аналогій відбувається підміна аргументу! Колір маку — ознака, що може залежати від стану одного гену, а колір шкіри людини, тим більш — її «енергія, мужність, винахідливість, терпіння, самовладання, витримка» не є моногенними ознаками! Ті ознаки, які згадує Дженкін, залежать від дуже складної взаємодії великої кількості спадкових ознак (і від розвитку в сприятливих умовах). Навіть, якщо аргумент Дженкіна пояснюють без конкретизації, про які ознаки йдеться, відбувається заміна однак, що залежать від одного гену чи невеликої кількості генів на ознаки, що є похідними ледве не усього генотипу.
«По суті, менделізм підірвав твердження Флемінга Дженкіна про те, що природний відбір є неефективним, оскільки нові ознаки з'являтимуться внаслідок схрещування з незмінними особинами. Якщо спадковість є частковою, а не змішаною, то сприятлива нова ознака зберігатиметься недоторканою, і її частота поступово збільшуватиметься в популяції. <...> Там, де існувала значна генетична мінливість, оскільки кілька генів або алелів впливали на одну й ту саму ознаку, варіація підтримувалася в популяції лише тому, що спадковість зберігала гени-варіанти недоторканими з покоління в покоління».
Петер Боулер, «Менделівська революція», 1989.
Сутність аргументу Дженкіна полягає в тому, що в умовах сексуального відтворення ознаки, що забезпечили успіх певної особини, можуть «розсипатися» у наступних поколіннях, якщо вони залежать від взаємодії великої кількості генів. Успішна особина з вищою ймовірністю залишить успішне потомство, якщо буде відтворювати свій генотип, схвалений у ході взаємодії з середовищем, ніж у разі, якщо буде поєднувати випадково обрану половину своїх генів з випадково обраною половиною генів іншої, нехай також дуже успішної особини. Цей феномен не здається вам дивним? Подивіться на нього з наступного погляду.
Ми розглядаємо еволюцію як вдосконалення програм, що впливають на розвиток організмів. Генотипи, що сприятимуть розвитку фенотипів, які краще відповідатимуть взаємодії з середовищем, з більшою ймовірністю зберігаються та відтворюються. «Переможці природного добору» — це ті, хто вижив і розмножився; ймовірно, на їхній успіх впливала не лише випадковість, а й досконалість, взаємоузгодженість їхнього генотипу. І що роблять ці «переможці»? Руйнують свої генотипи, комбінуючи їх фрагменти з з фрагментами генотипів інших «переможців природного добору»! Уявіть собі альпініста, який піднявся на Джомолунгму; він подолав складний шлях, минаючи тіла переможених, адже повідомлюють, що схили Джомолунгми усипані трупами альпіністів-невдах. Ось, наш «переможець» на вершині — тобто, відповідно до нашої аналогії, вижив та приступає до розмноження. Він має певний комплект спорядження (геном); ймовірно, раз з цим комплектом він здолав усі перешкоди, цей комплект довів свою досконалість: надійність та взаємодоповнюваність. Вчинок альпініста, який почне обмінюватися випадково відібраними частинами спорядження з кимось іншим, хто дістався до цієї ж вершини, ймовірно, має нас здивувати. Тоді нас має здивувати й характерний для «переможців природного добору» спосіб реорганізації їх власних геномів, що сприяли іхньому успіху!
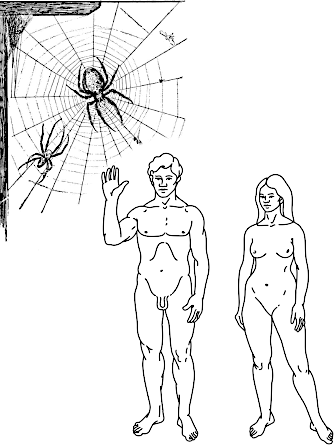
Ще один аспект, який можна пояснити за допомогою цієї аналогії, пов'язаний з тим, що для розмноження «переможця природного добору» потрібний інший «переможець», причому у більшості випадків — не будь-який, а такий, що належить до іншої статі. Переваги перемоги використає тій, хто підійметься на вершину алегорічної Джомолунгми не сам... За літературною аналогією це обмеження можна позначити як парадокс Робінзона Крузо (рис. IV-19.2). Прототипом персонажу роману Данієля Дефо був британський моряк Александр Селкірк, що перебував на безлюдному острові біля узбережжя Чілі у 1704-1709 рр. Робінзон Крузо, згідно з романом, провів на острові у самотності 24 роки, а потім врятував тубільця, якого назвав П'ятницею. Після 4 років життя вдвох з П'ятницею Робінзон зміг покинути острів. Якби популяції людини належали до I типу за показаною у табл. IV-19.1 класифікацією, за 24 роки Робінзон Крузо заселив би острів своїми клоновими потомками. Якби популяції людини належали до II типу, Робінзону довелося б чекати П'ятницю, зате їхні потомки були б рекомбінованими і відчували б усі переваги сексуальної репродукції. Але популяції людей належать до III типу і це накладає важливі екологічні обмеження на пошук партнерів для розмноження.
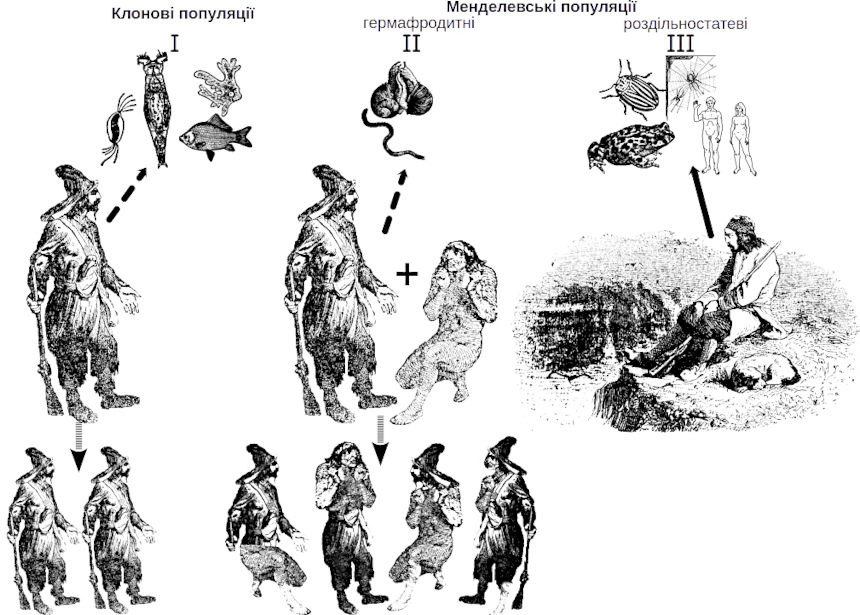
Рис. IV-19.2. Парадокс Робінзона Крузо: якщо б люди відтворювалися клоново, Робінзон Крузо заселив би острів своїми клоновими потомками; якби люди були гермафродитами — з'явилися б рекомбінантні потомки Робінзона та П'ятниці. Але люди відтворюються «найневдалішим» для поширення на нові території способом, роздільностатево, і саме тому заселити острів своїми потомками Робінзон не зміг
У такому разі виходить, що поки справи йдуть добре (або нема відповідної компанії), краще відтворюватися клоново, а до сексуальної репродукції можна звертатися лише тоді, якщо починають накопичуватися проблеми, причому у разі наявності потрібного партнера. Таке припущення можна підтвердити низкою прикладів. Так, для низки груп, що демонструють чергування сексуального та асексуального розмноження (дафнії, попелиці тощо) типовим є випадок, за якого в сприятливих умовах відбувається асексуальне (партеногенетичне) розмноження, а при погіршенні умов (наприклад, під час осінніх холодів) відбувається перехід до сексу та утворенню зимувальних яєць (рис. IV-19.3). Життєві цикли, що подібні показаному на рис. IV-19.3, можуть допомогти у розумінні наступного парадокса (який ми назвемо парадоксом Вільямса та обговоримо дещо пізніше, у зв'язку проблемами, що пов'язані з виникненням та підтримкою роздільностатевості).
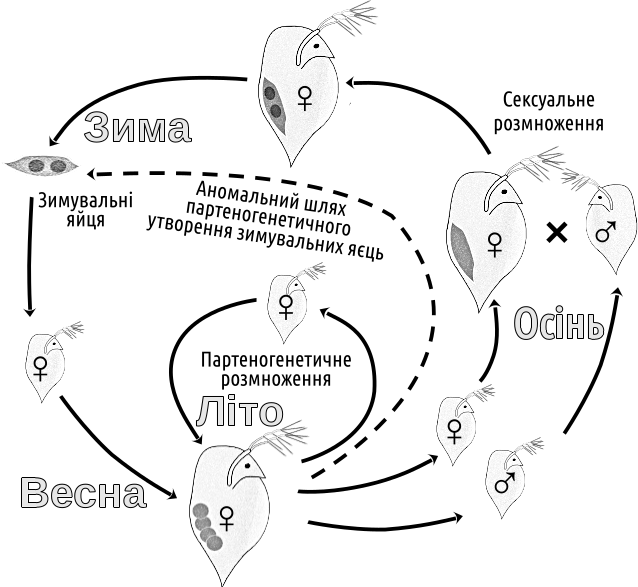
Рис. IV-19.3. Типовий річний цикл дафній
Ми вже зазначили, що не маємо достатніх даних для того, щоб впевнено описувати перші організми, що використали секс. Цілком можливо, що секс не був пов'язаним у них безпосередньо з розмноженням; це міг бути засіб виникнення гібридних генотипів, розмноження яких відбувалося на наступному етапі життєвого циклу. Ядра яких клітин зливалися при заплідненні? Цілком ймовірно, це могли бути ядра неспеціалізованих клітин. Різноманіття форм сексуального процесу до цього часу ми можемо спостерігати у різноманітних водоростей та грибів. Так, запліднення, за якого зливаються дві неспеціалізовані вегетативні клітини водоростей має назву кон'югації, а у разі злиття двох вегетативних клітин, що мають джгутики (що спостерігається й у деяких водоростей, й у деяких грибів) — хологамія. Вже після поширення сексу клітини, що беруть участь у заплідненні, набули спеціалізації та перетворилися на гамети. Нема підстав очікувати, що ці гамети відразу були розподілені на два типи, великі та дрібні, яйцеклітини та сперматозоїди. Якщо еволюція сексу проходила через етап хологамії, наступним кроком могла бути ізогамія, випадок, коли у заплідненні беруть участь дві рухливі гамети, що є морфологічно однаковими. Анізогамія — це випадок, коли з двох рухливих гамет одна є більшою, а інша — дрібнішою. Анізогамія могла бути переходом до оогамії — найпоширенішого випадку, коли запліднення відбувається між позбавленою джгутиків відносно великою яйцеклітиною та дрібним рухливим сперматозоїдом.
Перехід від ізогамії до анізогамії та оогамії має непогане теоретичне пояснення, що часто позначають як ПБС-теорією (PBS-теорією, за першими літерами прізвищ авторів статті G. Parker, R. Baker, R. and V. Smith (1972) «The origin and evolution of gamete dimorphism and the male-female phenomenon»). Згідно з підходом ПБС-теорії, в популяції, де нормою є продукування гамет, що несуть приблизно половину запасу поживних речовин, що є необхідним для розвитку зиготи, можуть отримувати перевагу особини, що продукують дещо збільшену кількість дещо дрібніших гамет. У таких умовах збільшення кількості потенційних потомків може сильніше впливати на пристосованість таких особин, ніж зниження виживаності потомства. В умовах, коли поширюються дрібні гамети, перевагу можуть отримувати особини, що продукують збільшені гамети, що містять достатній запас поживних речовин поза зв'язку з тим, скільки їх надасть інша гамета.
Втім, слід підкреслити, що перехід від ізогамії до оогамії може бути пов'язаним з переходом від відсутності статі (і продукування гамет, що не поділені на два типи) до роздільностатевості (продукування певного типу гамет) чи до гермафродитизму (продукування двох типів гамет водночас чи послідовно). Ми не можемо впевнено сказати, чи є гермафродитизм первинним (спочатку виникли гермафродити, а після них — самці та самиці окремо), чи вторинним (використовуючи позначення, використані в табл. IV-19.1, можна припустити, що популяції II типу потенційно можуть виникати з популяцій типу IIIa). Так чи інакше, важливим історії величезної кількості груп відбувся загадковий перехід до роздільностатевості. Почнемо з парадокса, що пов'язаний з появою двох статей.
Парадокс Мейнарда Сміта або парадокс подвійної вартості самців («two-fold cost of males») чи парадокс подвійної вартості сексу («two-fold cost of sex») полягає в тому, що перехід до роздільностатевості призводить до подвійного зниження репродуктивного потенціалу популяції. Порівняймо кількість потомків у двох особин у разі сексуального (роздільностатеві організми, тип III за табл. IV-19.1) та асексуального (наприклад, клонового партеногенетичного, тип Ib) розмноження. Позначимо розмір виводка, що приносить роздільностатева або партеногенетична самиця, як n. У разі використання сексу кількість потомків у батьківської пари дорівнюватиме n, у разі відмови від сексу — 2×n. У наступному поколінні сексуальні організми можуть отримати n×(n/2) потомків (адже кількість потомства залежить від чисельності самиць, що складають половину особин в кожному поколінні), а асексуальні — 4×n2! Це — дуже вагома різниця! Обговорюючи походження сексу, ми погодилися з припущенням Фішера, що він міг бути підтриманим груповим добором. Відповідно до парадоксу «two-fold cost of males» груповий добір мав би потужно «грати» проти сексу та роздільностатевості!
«По суті, партеногенетична самиця має подвійну селективну перевагу, оскільки вона не витрачає енергію на народження синів. Звичайно, цей аргумент справедливий лише за інших рівних умов, тобто, якщо немає генетичних відмінностей, що впливають на пристосованість між сексуальними та асексуальними самицями».
Джон Мейнард Сміт. «Еволюція сексу», 1978
До речі, яке з використовуваних позначень, «two-fold cost of males» або «two-fold cost of sex» є кращим? Поширенішим є друге, але більш точним — перше. Наприклад, популяції гермафродитів, популяції II типу згідно з класифікацією, що показана в таблиці IV-19.1, не платять цю подвійну вартість: секс у них є, а самців — нема.
Може постати закономірне питання — чи може бути «вартість самців» не подвійною, а меншою? Уявимо популяцію, де 9/10 самиць і лише 1/10 самців. В такій роздільностатевій популяції кількість потомків сягатиме 90% від такої в клоновій. У такому разі «вартість самців» дорівнюватиме не 1/0,5=2, а тільки 1/0,9=~1,11 — набагато менше. Якщо «вартість самців» так легко знизити, чому ж ми так часто спостерігаємо співвідношення статей, наближене до 1:1? Це питання має загальновизнане пояснення, яке Фішер надав 1930 року.
Рівновага Фішера (або принцип Фішера) — принцип, причина якого полягає в тому, що у роздільностатевих популяціях кожна особина має одну мати та одного батька. Внаслідок цього в популяції, де 9/10 самиць і лише 1/10 самців, самець матиме в середньому в 10 разів більше потомків, ніж самиця! У такому разі добір сприятиме тим особинам, у потомстві яких частка самців буде вищою, ніж 1/10. Частка самців зростатиме, але не безконечно. Коли припиниться її зростання? Коли середня кількість потомків у самця дорівнюватиме такій у самиці. Насправді ситуація є дещо складнішою. У разі, коли продукування потомка-самця та потомка-самиці вимагатиме різних витрат енергії для батьків, добір вирівнюватиме не очікувану кількість подальших потомків на одне дитинча, а таку кількість потомків на однакову витрату енергії. Втім, у більшості випадків витрати на потомків різної статі приблизно однакові, і це є причиною того, що співвідношення статей у більшості роздільностатевих популяцій наближається до рівного. Ще одним цікавим наслідком є те, що гермафродитні особини з перехресним заплідненням мають витрачати однакову кількість енергії на продукування жіночих та чоловічих гамет!
«Розглянемо спочатку гермафродитний вид тварин із зовнішнім заплідненням (наприклад, асцидій), або, якщо хочете, самонесумісну гермафродитну чи однодомну рослину. Такі види, як правило, розподіляють ресурси, що виділяються для розмноження, порівну між яйцеклітинами і сперматозоїдами або між насінням і пилком. Причина такого рівного розподілу, по суті, та сама, що дозволила Фішеру (1930) передбачити співвідношення статей 1:1 у роздільностатевих видів <...> Звідси випливає, що партеногенетична особина, якій не потрібно виділяти жодних ресурсів на виконання чоловічих функцій, могла б виробляти вдвічі більше яєць або насіння».
Джон Мейнард Сміт. «Еволюція сексу», 1978
До речі, рівновага Фішера є прикладом того, що у багатьох випадках добір на рівні генів «виграє» у групового добору. Хоча популяції з суттєво зсунутим співвідношенням статей могли б бути набагато успішнішими, ми не спостерігаємо їх навколо нас (в усякому разі — у типовому випадку). Добір на збільшення репродуктивного успіху кожної особини (чи, з іншого погляду, — на більш ефективне відтворення її генів, що впливають на співвідношення статей у потомстві) підтримує тих особин, що продукують відносно однакову частину потомків обох статей.
Таким чином, популяції, що переходили до роздільностатевості, мали подолати високий поріг: вдвічі знизити ефективність відтворення. Більше за те: якщо на будь-якому етапі роздільностатеві популяції відмовляться від свого парадоксального способу відтворення, вони мають отримати відповідні преференції! Цей феномен також детально розглядав Джон Мейнард Сміт, але посилався він при цьому на Джорджа Вільямса. Вільямс розглядав ситуації, коли два способи відтворення реалізовані в одній популяції, як на рис. IV-19.2. Звісно, у показаних на рисунку дафній співіснування сексуального та асексуального відтворення є нормою. З іншого боку, можливо розглянути й інший випадок, коли асексуальне відтворення з'явиться в сексуальній популяції (наприклад, внаслідок появи партеногенетичної форми, що відмовилася від мейозу). Переваги сексуального відтворення є «повільними», вони проявлять себе в еволюційно значущій перспективі. Переваги сексуального відтворення мають бути «швидкими», такими, що проявляються протягом короткого терміну.
«Найсильніший аргумент на користь швидкої переваги сексу висунутий Вільямсом (1975) як «балансовий» аргумент. Якщо партеногенетичний і статевий способи розмноження співіснують в одній популяції, то повинна існувати якась безпосередня перевага статевого розмноження, яка б врівноважувала його подвійний недолік. Дійсно, можна було б очікувати, що природний додбір змінюватиме відносні частоти сексуального і асексуального відтворення до тих пір, поки селективні переваги не врівноважаться. Сформулюємо той самий аргумент по-іншому. Груповий добір може підтримувати секс, усуваючи партеногенетичні різновиди, якщо останні виникають досить рідко, але не може підтримувати баланс між цими двома процесами, якщо групове вимирання не є надзвичайно поширеним».
Джон Мейнард Сміт. «Еволюція сексу», 1978
Парадокс Вільямса або балансовий аргумент Вільямса полягає в тому, що у разі поєднання в одній популяції сексуального та асексуального відтворення (як-от на рис. IV-19.3) асексуальне матиме «швидку» (таку, що проявиться протягом лічених поколінь) перевагу. Щоб сексуальне відтворення не зникло, необхідно, щоб його весь час підтримувала якась інша «швидка» перевага.
Чи є поява партеногенетичної самиці в роздільностатевій популяції чимось реалістичним? Так; подібні явища зареєстровані у великій кількості груп (за виключенням ссавців, у яких, ймовірно, партеногенез заблокований епігенетичними механізмами, що забезпечують геномний імпринтинг). Наведемо приклад бірманських пітонів (Python bivittatus), яких раніше вважали підвидом тигрового пітона (застаріла назва — P. molurus bivittatus). Це — великі і красиві змії, яких нескладно утримувати та розмножувати у неволі. Завдяки цьому накопичені численні дані щодо репродуктивної біології цього виду. Відомо, що іноді пітони виходять з незапліднених яєць, що відкладає самиця, яка не парувалася з самцями; останнім часом було доведено, що такі особини — самиці, і що вони генетично ідентичні своїй матері. Наскільки спадковою є здатність до партеногенезу у таких самиць, невідомо, але можна припустити, що, як і у багатьох інших подібних ознак, у схильності до партеногенезу має бути суттєва спадкова компонента.