| ← | → | |
| Приклади різноманітних моделей, їх задум і дизайн | Ідеї для моделювання |
Рекомендовані позначення в популяційно-екологічних моделях |
| Сотворіння світів-10 | Сотворіння світів-11 | Сотворіння світів — Додатки |
"Біосфера", що складається з "екосистем", що можуть перебувати у стійких або нестійких станах. За яких умов така біосфера буде еволюціонувати до більшої стійкості без дарвіновського добору та репродукції окремих одиниць?
Проблема має сенс, наприклад, з точки зору дискусії Дж. Лавлока та Р. Докінза. Концепція Геї Лавлока передбачає доцільні механізми планетарної регуляції. Доцільність — результат добору. Докінз стверджує, що добір завжди пов'язаний з діференціальним відтворенням реплікаторів. Якщо б існувало багато біосфер, деякі гинули, а інші — розмножувалися, тоді б, за Докінзом, біосфери могли розвинути здатність реагувати доцільно. Згідно з припущенням, яке можна перевіряти за допомогою моделювання, доцільність може бути наслідком і іншого типу добору, який можна назвати добором за Больцманом — поширенням більш термодінамчіно ймовірних (більш стійких) станів (деякі пояснення — тут). Якщо це припущеня є вірним, підсистеми такої системи як біосфера можуть знаходитися у станах різної стійкості. Чи може добір (перехід у більш термодінамічно ймовірні стани) таких підсистем бути механізмом розвитку доцільності у системи, що не залучена до процесу диференціальної реплікації?
Аналогія. Чи може добір організмого рівня бути причиною доцільності популяційних механізмів регуляції без дарвіновського добору (диференціальної загибелі та диференціальної реплікації, тобто розселення) популяцій?
Чи буде зростати гіперболічно (як чисельність людства за фон Ферстером) чисельність популяції, що існує не необмеженому двомірному просторі та здатна з певною швидкістю розселюватися у незайнятих напрямах? Точніше, за якою моделлю вона зростатиме (відносне збільшення площі ареалу буде зменшуватися).
Конкуренція за спільний ресурс двох популяцій. Чисельність однієї контролюється хижаками, чисельність іншої — нестачею ресурсу. За яких умов хижаки забезпечать "перемогу" своїх жертв над їх конкурентами?
Вплив кризи перенаселення на ємність середовища. Слони, що живляться акаціями у саванні. Коли їжі достатньо — поїдають листя, коли не вистачає — ламають стебла та обдирають камбій. Розширення моделі — вплив на цей процес гомотеріїв та смилодонів.
Істоти Деннета! Детальніше тут і тут. Скінерова істота, що буде радіти виграшам та сумувати у разі програшів.
Перехід популяції птахів, що живуть на витягнутому вздовж меридіана континенті, від осілості до перельотів. У високих широтах кращі умови для розмноження, у низьких — вищі шанси пережити зиму.
Регуляція річного циклу у популяції, яка живе у помірному кліматі: на що краще реагувати, на динаміку температури чи довжину світлового дня? Порівняти різні способи регуляції за кількістю "помилкових" днів за різних рівнів мінливості клімату.
Жіночі та чоловічі репродуктивні стратегії людини.
Різні горизонти планування за різного рівня мінливості середовища.
«Немногие для вечности живут,
Но если ты мгновенным озабочен —
Твой жребий страшен и твой дом непрочен!
(О.Мандельштам)
Конкуренція двох популяцій із статевим розмноженням та двома статями в умовах стабілізуючого відбору. В одній мінливість і смертність (що прибирає особин, що відхиляються від норми) однакові для самців і самок, в іншій — самці мінливіше і, отже, сильніше страждають від негативного відбору. Ідея В. Геодакяна.
Порівняння швидкості вироблення адаптацій у разі клонального та статевого відтворення.
Порівняння поширення нових ознак при генетичному та культурному наслідуванні.
Фазовий перехід в еволюції (від генів до мемів, від мемів до т-мемів і далі).
Повільні та швидкі інфекції за умов різної популяційної структури.
Принцип Фішера при різній "ціні" особин різних статей або у разі зміни співвідношення статей під час онтогенезу.
Хроповик Меллера.
"Гіпотеза бабусі" та походження клімаксу у людини.
"Стокротковий світ" Лавлока.
ІмІтаційне доведення парадоксу Монти Голла. Модель, яка демонструватиме успішність двох різних стратегій (міняти вибір чи не змінювати) можна зробити за аналогією з моделлю про нарожденння хлопчиків та дівчинок (що вирішує одну из відомих демографічних задач). Якось Великий Султан зіштовхнувся з складною проблемою. Представники збіднілої знаті стали скаржитися, що їм складно набирати собі гареми, що відповідають їхньому статусу. Дівчаток народжується приблизно стільки ж, скільки і хлопчиків, і після того, як багатії заберуть до себе в гареми по сотні дружин, їм, власне, нікого й не лишається. Щоб вирішити проблему, Султан вирішив збільшити кількість народжуваних дівчаток по відношенню до кількості хлопчиків. Для цього він видав такий указ: «Жінці дозволяється народжувати дітей доти, доки вона не народить першого хлопчика. Після цього їй народжувати забороняється. Таким чином, у кожній родині може бути не більше одного хлопчику, а дівчаток скільки завгодно». Наскільки зміниться відношення кількості дівчаток до кількості хлопчиків після виконання цього указу?
Ймовірно, у такій моделі треба буде згенерувати безліч випробувань з випадковим розміщенням кози та автомобілів, відкриванням одного з дверей з козою та підрахунком виграшів двох гравців, один з яких змінює двері, а другий – ні. Збирати статистику результатів цих спроб слід за допомогою лічильника ітерацій, аналогічному такому в моделі про хлопчиків або моделі "Гаузе".
Динаміка концентрації кисню під час кисневої революції. Треба врахувати:
— запас відновників, який поступово витрачається;
— приплив відновників з надр, що поступово слабшає;
— відносно постійну втрату атмосфери;
— зниження ефективності примітивних фотосинтетиків у міру виникнення локальних острівців кисню;
— появу і поширення більш досконалих фотосинтетиків, що ефективно працюють в окисному середовищі.
А потім можна спробувати підібрати параметри так, щоб динаміка концентрації кисню приблизно відповідала тому, що є відомим з геологічного літопису.
Перехід від гермафродитизму з перехресним заплідненням до роздільної порожнини. Модель та її опис є тут, але тема далеко ще не вичерпана. Аналогічна ситуація з переходом від ізогамії до оогамії (і, взагалі, від злиття при заплідненні одноклітинних організмів до використання спеціалізованої стадії розвитку — гамет).
Розмножуватись інтенсивно і жити недовго, чи розмножуватись стримано і намагатися жити довго? Внутрішньопопуляційні онтогенетичні стратегії у постійному чи мінливому, сприятливому чи суворому середовищі. У якому разі вони працюють ефективніше: за великої спадкової компоненти у визначенні стратегії чи залежності стратегії від короткострокових умов?
Зв'язок між вибагливостю однієї статі та мінливостю іншої.
Гра, що обговорюється в цьому тексті; розподіл її результатів.
Чому всі євреї є нащадками царя Соломона, а всі українці — Ярослава Мудрого? Яка ймовірність того, що якийсь із членів репродуктивної сукупності, який жив певну кількість років тому і залишив нащадків, є предком усіх сучасних членів цієї сукупності?
Партеногенез у хлыстохвостых ящериц
Прежде всего, предлагаю посмотреть следующий фрагмент фильма. Речь идет о хлыстохвостых ящерицах.
Как можно понять, речь идет о популяциях с клональным размножением, где потомки генетически идентичны своим родителям. Все особи в нем — самки, но они способны под влиянием всплеска мужских гормонов выполнять мужскую роль, имитируя спаривание. Такая имитация спаривания необходима для самок, чтобы у них происходило созревание яиц. В связи с этим фрагментом фильма возникает множество вопросов.
1. Если в популяции таких ящериц возникнет особь, не способная претерпевать всплеск мужских гормонов и помогать размножаться другим самкам, будет ли отбор благоприятствовать этой особи и ее клональным потомкам? Это представляется достаточно вероятным. Как минимум, ее потомки не будут испытывать конкуренции со стороны потомков других клонов. В общем случае отношения между самками, которые помогают другим особям беременеть, и теми, которые этого не делают, можно рассматривать как отношения между "альтруистами" и "эгоистами". Всегда ли отбор будет способствовать "эгоистам"? Существуют ли условия, в которых преимущественно будут распространяться "альтруисты"?
2. Как мог произойти переход от нормального размножения с двумя полами, рекомбинацией и оплодотворением, к варианту, показанному в данном видеофрагменте? Для этого, вероятно, понадобилось закрепление у клонов сразу двух необычных особенностей: способности к партеногенетическому развитию яйцеклеток и способности самок временно переходить в гормональное состояние, характерное для самцов. Как мог происходить такой переход?
Хоровое пение зеленых лягушек
Все мы знаем, что зеленые лягушки, сидящие по берегам водоема, поют не хаотично, а хором: то вместе кричат (перекрикивая друг друга), то вместе молчат. Альфред Брем рассказывает о старом немецком поверьи, что в каждом пруду среди лягушек есть свой "хормейстер". Может ли хоровое пение лягушек быть результатом самоорганизации, который не требует никакого "хормейстра"?
Один из способов ответа на этот вопрос — построение модели. Например, для построения простой модели можно воспользоваться гидравлической моделью инстинктивного поведения, предложенной Конрадом Лоренцем. Несмотря на критику этой модели, она в простой форме отражает несколько простейших наблюдений. Так, в том случае, если животное в ходе своей жизнедеятельности должно переходить к определенной форме поведения, сплошь и рядом удается наблюдать следующее. Переход к ожидаемому поведению часто является ответом на определенный стимул (релизер). Чем дольше животное ожидает этого релизера, тем чувствительнее к нему оно оказывается. Если животное находится в ожидании стимула-релизера, а его нет, оно может переходить к соответствующей форме поведения самопроизвольно.
Простой моделью (!), описывающей эти закономерности, и является гидравлическая модель Лоренца. В ней описанные особенности инстинктивного поведения объясняются накоплением в животном некой специфической энергии. Чтобы прояснить гидравлическую модель, можно воспользоваться ее описанием (достаточно удачным) из Википедии.
Конрад Лоренц, в частности, для объяснения феномена срабатывания инстинктивных реакций в отсутствие специфического раздражителя, предложил оригинальную модель осуществления инстинктивного поведения. Модель была построена на основе принципов гидравлики и получила название «гидравлической модели Лоренца». В гидравлической модели специфическая энергия действия представлена водой, непрерывно текущей в резервуар. Отток воды из резервуара преграждён клапаном, к которому крепится груз. Груз обозначает действие специфических раздражителей, его вес прямо пропорционален интенсивности раздражителя. Вода из резервуара может вытечь в двух случаях: если суммарное давление воды и груза откроют клапан, либо уровень воды превысит допустимый и клапан откроется под напором воды.[13]
Таким образом, по мере накопления специфического потенциала действия снижается порог, необходимый для запуска соответствующей инстинктивной реакции. В случае, когда необходимый раздражитель отсутствует в течение долгого времени, реакция может осуществиться «вхолостую», без специфического стимула.
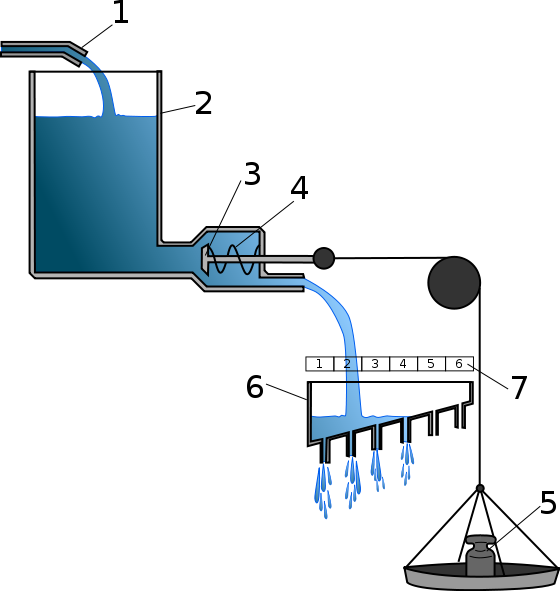
1-приток специфической энергии действия (СЭД) создаёт давление в резервуаре (2)
3,4 - Врождённый спусковой механизм. Испытывает давление со стороны СЭД (3) и релизера (5)
Лоренц сам признавал ограниченность гидравлической модели. Хотя она и объясняет некоторые наблюдаемые феномены, но обладает рядом серьёзных недостатков. Прежде всего, она не учитывает изменчивость поведения. Во-вторых она предполагает существование некоторой лишней сущности — гипотетической энергии. Следует отметить, что нервная структура, ответственная за накопление специфического потенциала действия, не была обнаружена. В третьих, имеются факты, противоречащие данной теории.
Благодаря своей простоте и наглядности, гидравлическая теория изучается в университетских курсах зоопсихологии по сей день.
Подчеркну, что перечисленные в статье Википедии недостатки гидравлической модели ни в коем случае не являются противопоказаниями для ее использования в моделировании. Не надо требовать от модели, чтобы она была идентична оригиналу по своей структуре; если модель обеспечивает то же поведение, которое характерно для оригинала, ее использование (пока она полезна) оказывается вполне оправданным. Описанная Лоренцем модель приводит к реализации тех особенностей поведения (повышение готовности прореагировать на релизер со временем и способность к спонтанному включению определенной формы инстинктивного поведения), которые были описаны во многих экспериментах.
Кстати, вы ведь поняли, что на схеме из Википедии обозначено цифрами 6 и 7? Реализация инстинктивного поведения и его интенсивность.
Итак, я предлагаю сделать имитационную модель, позволяющую проверить, объясняет ли гидравлическая модель Лоренца феномен хорового пения лягушек. Рассмотрим круглый водоем, по периметру которого равномерно сидят лягушки. Переход лягушек от молчания к пению объясняется на приведенном ниже рисунке (отражающем концептуальную модель, описывающую переход лягушек от молчания к пению и обратно).
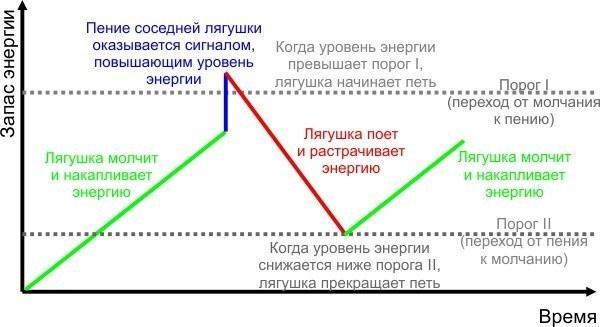
Как следует из модели, для описания поведения лягушек достаточно следующих основных параметров:
— скорость накопления энергии у молчащей лягушки (на сколько единиц повышается уровень энергии за каждый шаг модели);
— скорость снижения энергии у поющей лягушки;
— пороговое значение энергии для перехода лягушки от молчания к пению;
— пороговое значение энергии для переходя лягушки от пения к молчанию (этот порог ниже предыдущего);
— эффект пения соседних лягушек (повышение энергии у лягушки, которая на предыдущем шаге модели услышала пение другой лягушки) в зависимости от расстояния до соседки (вероятно, эффект пения ближайшей соседки самый высокий, соседки через одну — ниже, а через две и через три — еще ниже).
Вероятно, следует предусмотреть еще изменчивость некой случайной величины, от которой зависит значение порогов "включения" и "выключения" у каждой конкретной особи.
Фактически, для каждого шага для каждой особи надо проводить следующие вычисления:
1. Определяется, сколько энергии было у особи на предыдущем шаге, а также молчала они или пела. Если молчала, ее энергия увеличивается (в соответствии с заданной общей скоростью увеличения энергии у молчащей лягушки, с учетом индивидуальной поправки, зависящей от случайного числа.
2. Вычисляется сила внешнего стимула. К достигнутому уровню энергии добавляется его увеличение от пения ближайших соседей, с понижающим коэффициентом — от соседей "через одного", и с еще более низким — от соседей "через два"...
3. Если уровень энергии молчащей лягушки с прибавкой на стимулирование соседями превышает порог "включения" (вероятно, с учетом индивидуальной поправки, зависящей от случайного числа) — лягушка начинает петь.
4. Если лягушка молчит, ее энергия снижается (с соответствии с заданной общей скоростью снижения энегии у поющей лягушки, с учетом индивидуальной поправки, зависящей от случайного числа).
5. Если уровень энергии поющей лягушки с прибавкой на стимулирование соседями оказывается ниже порога "выключения" (вероятно, с учетом индивидуальной поправки, зависящей от случайного числа) — лягушка прекращает петь.
Только что перечисленные пять шагов являются приложением концептуальной модели управления пением лягушки (показанной на схеме) к алгоритму такого управления в модели.
Самым простым выходом модели может быть динамика количества поющих лягушек на каждом шаге. Можно также визуализировать распределение поющих и молчащих лягушек по периметру пруда.