| ← |
«Секс на R» |
→ |
| Секс на R–07: Еволюція та секс (Опис проблеми I) |
Секс на R–08: |
Секс на R–09: Моделювання парадокса Мейнарда Сміта (Завдання I) |
«Стать – королева проблем еволюційної біології»
«Стать — головний виклик сучасної теорії еволюції... королева проблем еволюційної біології», сказав американський біолог Грем Белл. Чому він так вважав? Чому з ним є сенс погодитися?
1. Парадокс Мейнарда Сміта (те, що роздільностатеве відтворення в популяціях чомусь не витісняється клональним) можна пояснювати:
— або наявністю внутрішнього запобіжника втрати роздільностатевості;
— або швидкою «карою» середовища за перехід до клонального відтворення.
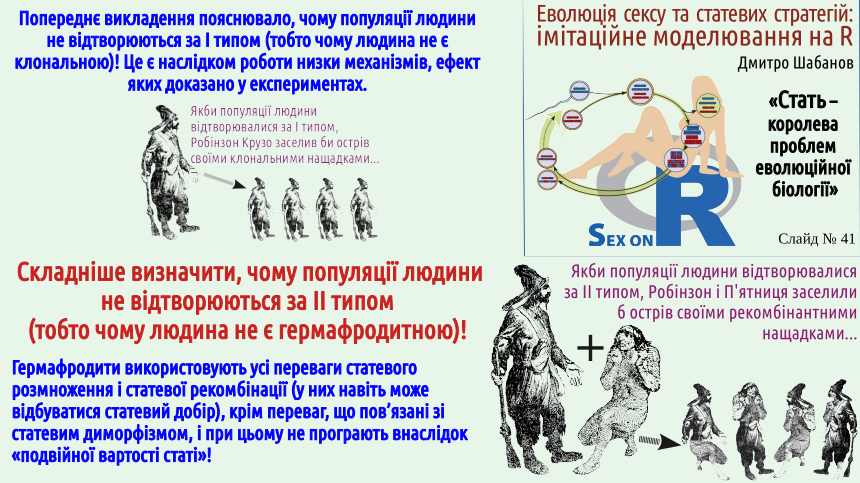
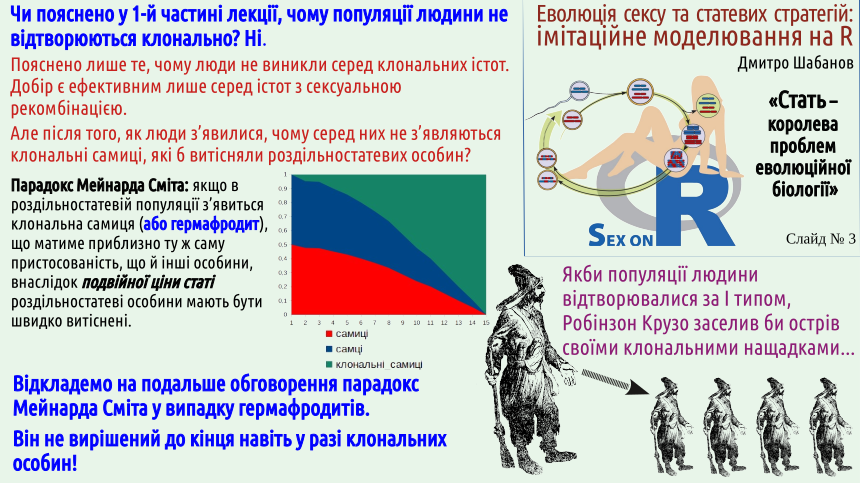
Наприкінці попередньої лекції пояснено парадокс Мейнарда Сміта. З одного боку, нам відомо чимало випадків появи клональних (наприклад, партеногенетичних) самиць. З іншого, ми бачимо стійке існування двостатевих популяцій, які, здавалося б, мали бути дуже крихкими, нестійкими до появи клональних особин. Чому це відбувається?
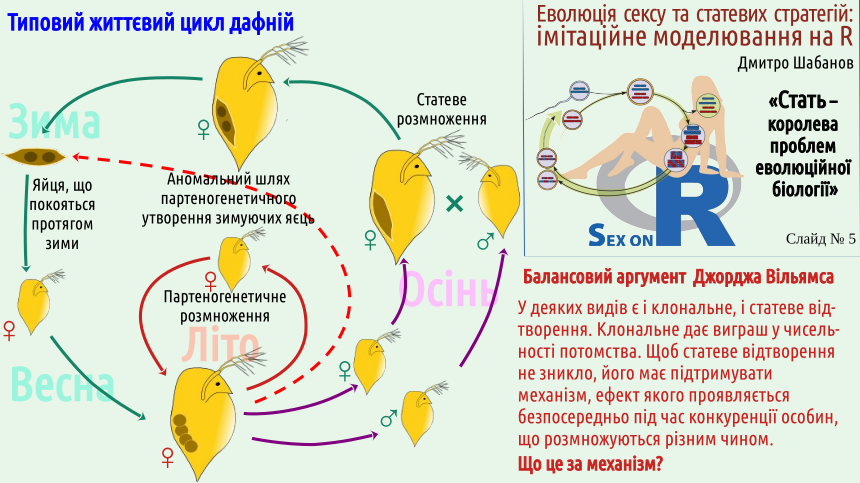
Відразу слід зазначити, що партеногенез та інші способи клонального відтворення відомі не у всіх груп тварин. Наприклад, невідомі партеногенетичні ссавці; ймовірно, механізм регуляції розвитку ссавців є складнішим за аналогічний механізм інших груп. Здається, відомим є лише один випадок партеногенезу у птахів (в індиків). А, припустимо, у рептилій, амфібій та риб випадків клонального відтворення відомо чимало (і ми розглядали його на прикладі рептилій: ящірок даревській. Серед численних груп безхребетних тварин відомо безліч випадків клонального відтворення; дещо далі в цій лекції ми згадуємо досить типовий випадок дафній, які чергують партеногенетичне та статеве відтворення (і тим самим в запропонованій класифікації типів відтворення має розташовуватися на якомусь містку між I та III типами — як це показати на схемі не дуже зрозуміло).
На слайді йдеться про те, що у людей могли б з'явитися партеногенетичні самиці. Скоріше за все, це неможливо внаслідок того, що люди є ссавцями. До речі, свого часу можливість партеногенезу у людини серйозно розглядалася як можливість наукового пояснення незайманого зачаття діви Марії. Це є непоганим прикладом того, наскільки невдалими стають такі спроби «всидіти на двох стільцях». З наукової точки зору це пояснення не годиться. Крім іншого, у видів з XY-системою визначення статі (а це, як раз, і випадок людини), потомство партеногенетичних самиць має складатися виключно з самиць. Відкіля в жіночій яйцеклітині могла б з'явитися Y-хромосома? Але це ідея не годиться і з точки зору віри. Для тих, хто вірить євангельській розповіді, незаймане зачаття і дивом, проявом порушення природних закономірностей, а не цікавою ембріологічною аномалією. Зверніть увагу: у цьому абзаці ми не приймали те, у що вірять сумлінні віряни, і не спростовували це, ми лише підкреслювали те, що наша розмова і віра лежать у різних площинах, і спроба їх поєднання є контрпродуктивною.
На наступному слайді також нагадується те, про що йшлося у попередній лекції.

На погляд автора, те, що в популяціях багатьох видів не відбувається перехід до партеногенетичного розмноження, який мав би відбуватися, згідно з парадоксом Мейнарда Сміта, може бути поясненим двома різними способами.
— Перше: у системі регуляції розвитку багатьох видів існує якийсь механізм (умовно назвемо його запобіжником втрати роздільностатевості). У такому разі постає складне питання: як міг виникнути такий запобіжник? Який добір міг його сформувати? Нагадаємо ще дві тези з попередньої лекції: першу, про те, що доцільність створюється добором, та п'яту, про відсутність передбачення в еволюції.
— Друге: популяціям, які могли б переходити до клонального відтворення, не дозволяє це робити якийсь зовнішній механізм (умовно назвемо його швидкою карою середовища за клональність). Довгострокова кара за клональність нам відома: це храповик Мюлера, але для накопичення несприятливих наслідків клональності потрібним є значно більший час, ніж для перемоги клонального відтворення. Знову згадаємо тезу про відсутність передбачення в еволюції.
Цей вибір можна сформулювати інакше. Самі популяції не переходять до клонального відтворення чи це не дозволяє їм середовище? Відповідь не така очевидна. Відомі спонтанні переходи до клонального відтворення у багатьох видів (не серед ссавців, але, припустимо, серед рептилій — достатньо часто). Є популяції, які легко чергують статеве та клональне відтворення. Балансовий аргумент Вільямса очікує певного впливу від середовища.
Можливо й таке, що не має сенсу ставити питання так: внутрішні АБО зовнішні механізми підтримують статеве розмноження. Можливо, працюють І внутрішні, І зовнішні механізми.
Блискучі дослідники висунули низку гіпотез, що пояснюють збереження статевого розмноження. Перелічимо їх певний набір, а далі розглянемо їх детальніше. До речі, і в цьому разі можливо, що різні механізми працюють водночас.

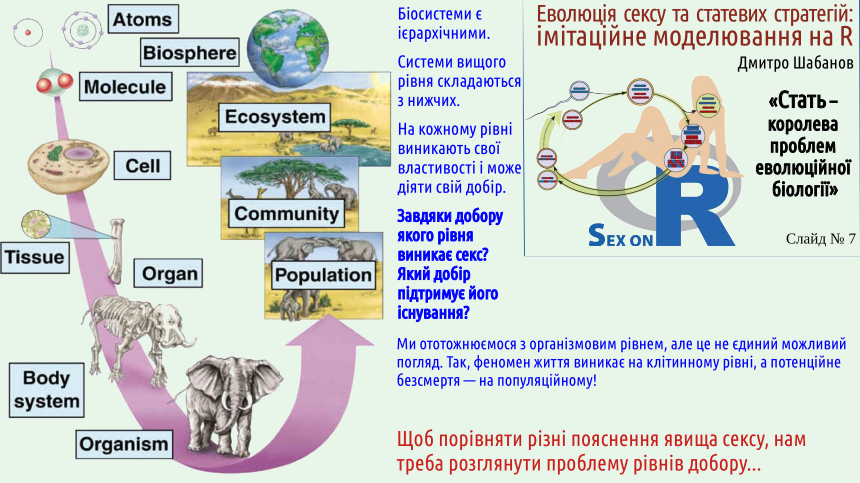
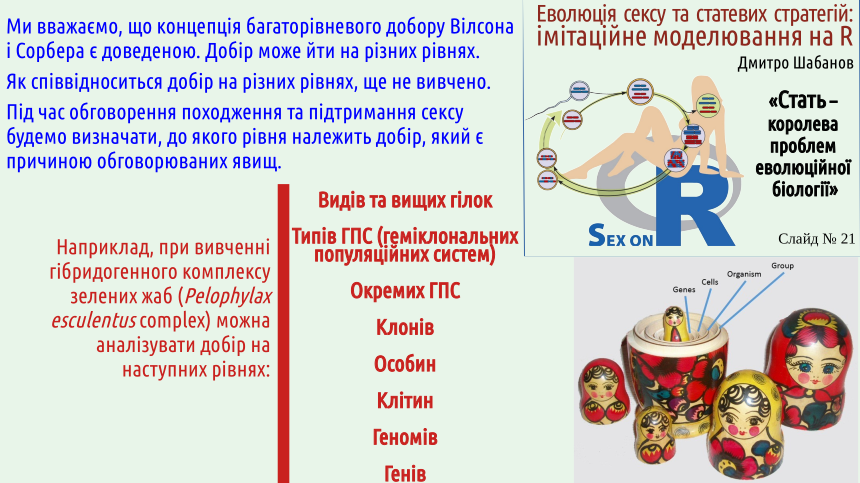
2. Біологічні системи є ієрархічними; теоретично добір може йти на різних рівнях ієрархії біосистем.
Під час характеристики перелічених гіпотез, що намагаються пояснити механізм підтримки статевого розмноження, ми будемо визначати, добір якого рівня створює та підтримує роботу цих механізмів. Для цього нам треба розібратися з проблемою рівнів добору.
На слайді показано певний варіант ієрархії рівнів організації біосистем (детальніше — тут). Слід підкреслити, що універсального набору рівнів не існує; у різних випадках може бути різною та кількість рівнів, на яких внаслідок об'єднання підсистем та їх взаємодії виникають нові якості. Підставою розглядати якусь біосистему як особливий рівень організації є саме виникнення у неї якихось властивостей, яких нема у її окремих частин. Оскільки популяція може бути безсмертною, а організм — ні, у нас є усі підстави вважати популяційний рівень окремим рівнем.
3. Різні автори розглядали добір як мінімум на генному, клітинному, організмовому, груповому (популяційному) та видовому рівнях.
На якому рівні діє природний добір? Класичний британський еволюціонізм був впевнений у відповіді: організменому, адже саме організми гинуть та виживають, залишають потомство або відсторонюються від розмноження. Дарвін (для якого були характерні пронизливі здогадки) припустив, коли обговорював еволюцію людини, і можливість добору груп.

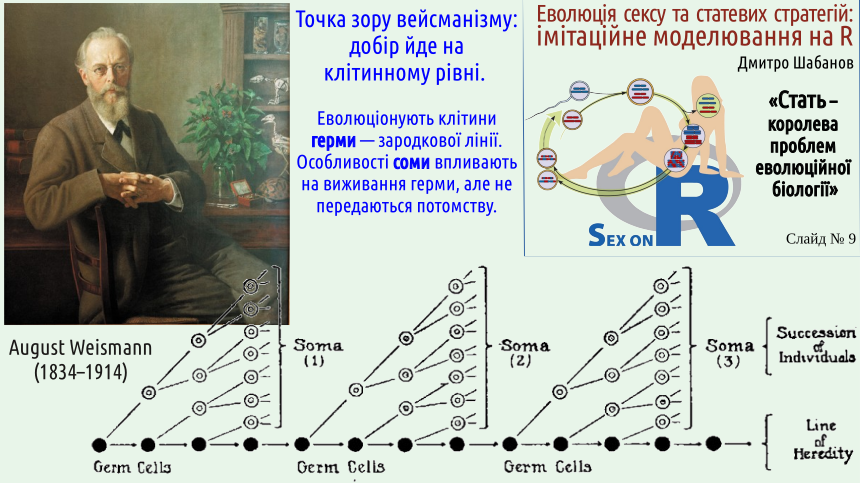
Вейсман зрозумів, що історію організмів можна розглядати й на клітинному рівні. Більшість клітин організму є смертними; потенційне безсмертя, перехід від покоління до покоління характерний для клітин зародкової лінії. Ми вже обговорювали різницю між сомою та гермою (зародковою лінією). Добір можна розглядати як щось, що йде і на клітинному рівні.
До речі, припустимо, під час запліднення у багатьох видів тварин неможливо не признати добір на клітинному рівні: з багатьох сперматозоїдів яйцеклітину запліднить один. Якщо це буде випадковий сперматозоїд, добору нема, є лише скорочення чисельності (адже добір — це успіх або невдача, що залежить від системи, яка є об'єктом добору). Якщо успіх сперматозоїда залежить від його властивостей (а, скоріше за все, так і є) ми спостерігаємо приклад добору на клітинному рівні.
У XX столітті відбулася революція. Почав її Холдейн, а фактом суспільної свідомості зробив її Докінз. У наступне покоління переходять не організми, а їх гени. В генофонді популяції ці гени переходять від особини до особини, утворюють різні комбінації, змінюються з часом. Таким чином, можна розглядати добір на генному рівні.
«Егоїстичний ген» Докінза став книгою, яка за своєю популярністю конкурує з «Походженням видів» Дарвіна. Багато хто впевнений в тому, що в цій книзі Докінз довів: добір йде виключно на рівні генів. Насправді навіть сам Докінз не стверджував це категорично. В «Розширеному фенотипі» — іншій своїй книзі, яка великою мірою присвячена темі рівнів добору. «Розширений фенотип» — набагато поміркованіша та розумніша книга, ніж «Егоїстичний ген» (і саме тому отримала набагато меншу популярність). Точка зору «Розширеного фенотипу» така: добір міг би йти й на інших рівнях, наприклад на рівні груп, але груповий добір буде неефективним у порівнянні з генним, і саме тому слід зосередитися на розумінні добору на генному рівні.

Ідея групового добору стала інтенсивно обговорюватися внаслідок ідей Веро Вінн-Едвардса. Він вважав такий добір важливим фактором, що спрямовує еволюцію.

Аргументи Вінн-Ердвардса розкритикував Джордж Вільямс (тій самий, що висунув балансовий аргумент). Розглянемо (може слід розглянути не лише на рівні обговорення, а й на рівні імітаційного моделювання) типову ситуацію.
Коли в певній популяції починаються голодні часи, багато організмів стримують своє розмноження. Припустимо, у роки, коли мало мишей, спеціалізовані хижаки, що живляться мишами, можуть стримувати розмноження. Гадюки можуть пропускати розмноження або народжувати меншу кількість дитинчат; сови зроблять одну кладку замість двох, і кількість яєць (і пташенят) у кожній кладці буде меншою, ніж у «мишачий» рік. Чому?
Версія Вінн-Едвардса така. Особини відчувають, що на популяцію чекає важкий рік і стримують розмноження, щоб зменшити конкуренцію та смертність.
Версія Вільямса така. Ті особини, що не витратять ресурси на розмноження (яке, внаслідок великої смертності, скоріше за все виявиться неефективним) матимуть більші шанси вижити та дочекатися на кращі часи. Навіть в умовах, коли смертність в популяції буде великою, ті особини, що продукуватимуть більшу кількість потомків, матимуть й більші шанси на виживання цих потомків. Якщо особина внесе більший внесок у майбутнє популяції, вона буде підтримана добором, навіть в умовах, коли надмірне розмноження буде наносити шкоду на популяційному рівні.

Дискусія щодо рівнів добору ще ускладнилася після того, як блискучі американські палеонтологи припустили, що одиницею добору можуть бути й цілі види.

Чи повним є розглянутий нами перелік рівнів, на яких може йти добір? Насправді ні. Можна, наприклад, згадати концепцію екосистемної еволюції, яка зосереджує увагу на стійкості або нестійкості екосистем (чи угруповань у їх складі) в цілому.
Втім, в XX столітті найпопулярнішими сталі ідеї, що еволюцію слід пояснювати добором на генному рівні. Здавалося, що один з найавторитетніших біологів, Едвард Вілсон, був одним з архітекторів такої точки зору. Крім іншого, Вілсон став творцем соціобіології — науки про еволюцію соціальної поведінки внаслідок добору генів.

4. Переконливо виглядає концепція багаторівневого добору, згідно з якою добір може йти на різних рівнях водночас.
Девід Вілсон (однофамілець Едварда Вілсона) і Еліот Собер наприкінці XX століття висунули дуже просту ідею: добір працює водночас на кількох різних рівнях, що вкладені один в одного як матрьошки.

Скандал, що пов'язаний з проблемою рівнів добору, викликала зміна точки зору патріарха американської еволюційної біології Едварда Вілсона: він погодився з концепцією багаторівневого добору. Спільно з молодими математиками він зайнявся імітаційним моделюванням і визнав важливу роль групового добору. Між Вілсоном і Докінзом розпочалася гостра дискусія, в якій, кінець-кінцем, Докінз став заперечувати саму можливість групового добору, а Вілсон — саму можливість добору на генному рівні. Здається, і те, й інше — перекоси.

Є чимало прикладів доведеної дії «егоїстичних генів», наприклад таких, які втручаються у процес мейозу та забезпечують свою пріоритетну передачу у наступні покоління.

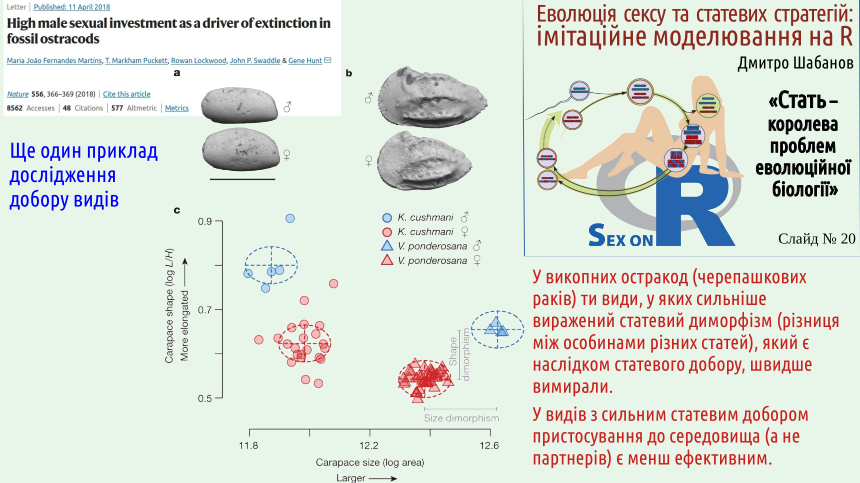
Є й добре доведені приклади добору на вищих рівнях, у тому числі — на рівні видів.

В обговорюваному прикладі ми бачимо, як добір на рівні видів впливає на еволюцію статевого розмноження у рослин. Перевага таких досліджень в тому, що сучасні молекулярно-генетичні методи дають змогу точно визначити, як гілкувалося еволюційне дерево у ході утворення сучасних видів.

На погляд автора, є усі підстави прийняти концепцію багаторівневого добору. Таким чином, ми припускаємо, що на ознаки, які нас цікавлять, може впливати добір, який діє на різних рівнях. Виникає можливість аналізувати, на якому саме рівні діє добір, який відповідає за ті або інші еволюційні зміни.
5. Запропонована низку гіпотез, що пояснюють переваги статевого відтворення. Деякі з цих гіпотез отримали експериментальні підтвердження. Особливої уваги заслуговують «сокира Кондрашова», гіпотеза статевого добору Менінга, гіпотеза «Червоної Королеви» Ван-Валена.
Тепер ми можемо перейти до обговорення гіпотез, які пояснюють стійкість роздільностатевості (їх перелік міститься на слайді № 6). Перша — це гіпотеза добору гілок Наннея.
Фактично, це спроба пояснити роздільностатевість наслідком добору видів. Нема підстав відкидати цю гіпотезу як неможливу, але усі процеси, які ми спостерігаємо, пояснити за її допомогою неможливо.
У комодоського варана іноді виникають партеногенетичні самиці. Це — представник достатньо давньої еволюційної гілки. І ми будемо вважати, що такі варани існували-існували, а зараз потрапили на межу зникнення внаслідок появи партеногенетиків? А чому добре себе почувають попелиці та дафнії?

«Сокира Кондрашова» працюватиме при певному характері взаємодії несприятливих алелів.

На наступних двох слайдах — ілюстрація гіпотези Кондрашова. Ми розглядаємо потомків від батьків, кожен з яких має по 5 шкідливих алеля. У потомків кількість шкідливих алелів визначається комбінаторикою; звісно, їх кількість може бути від 0 до 10; більше за усіх інших буде потомків, які будуть мати по 5 дефектів Ми розглядаємо ситуацію, коли наявність кожного дефектного алеля знижує життєздатність особини.
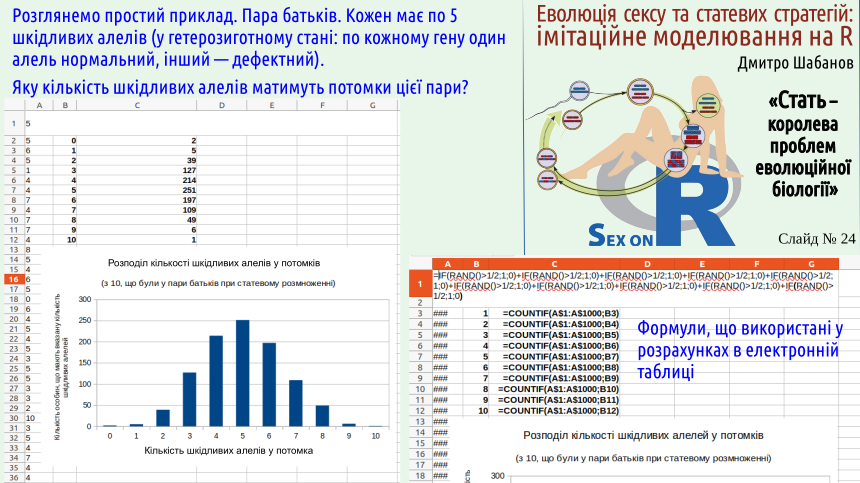
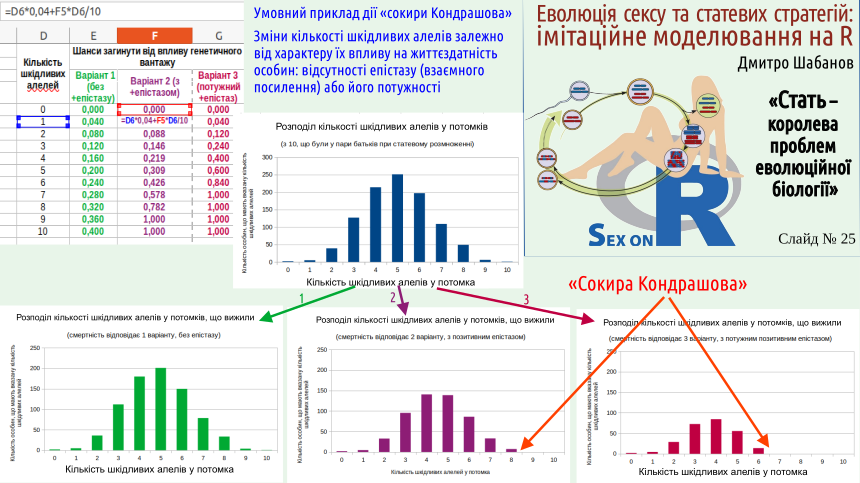
Наскільки гірше мати 2 дефектних алеля, ніж 1? Тут можливі різні варіанти. Може, мати два дефекти вдвічі гірше, ніж мати один. Такий випадок буде спостерігатися, якщо ці генетичні дефекти та ефекти від них ніяк не взаємодіють. Такий варіант показаний на слайді зеленим кольором. В ньому особина з одним дефектом має шанси померти 0,04, а з двома — 0,08.
Може бути й так, що мати два дефекти більше ніж вдвічі гірше, ніж мати один. Цей варіант показано червоно-фіолетовим, «буряковим» кольором. Носій двох дефектів має шанси померти 0,088. Це відповідає варіанту генетичної взаємодії, що має назву позитивного епістазу (взаємного посилення дії генів).
Малиновим кольором показано випадок, коли шкідливі алелі суттєво підсилюють дію один одного. Мати два дефекти втричі гірше, ніж мати два: шанси померти дорівнюють 0,12. Особини, що мають 7 та більше дефектів, виявляються за таких умов нежиттєздатними. На розподілі за кількістю генетичних дефектів цю частину ніби-то відрубає сокира. Цих особин, що перевантажені генетичними дефектами, можна порівняти з цапами-відбувайлами, які уносили з собою «гріхи» усієї популяції.
Чим цей механізм очищення добором від носіїв генетичних дефектів переважає те, що можна спостерігати у організмів з клональним відтворенням? Звісно, позитивний епістаз буде працювати і у клональних організмів. Але в клональних популяціях не будуть з'являтися «цапи-відбувайли», позбавляючись від яких популяція очищується від великої кількості шкідливих алелів.
Автору цієї лекції було б цікаво побудувати модель, де два види-конкуренти будуть боротися за спільний ресурс. Чи існують умови, за яких очищення від негативних мутацій у виду з сексуальним відтворенням дозволить перекрити несприятливий ефект «подвійної вартості статі»?
Цю лекцію ми почали цитатою з Белла про секс як королеву проблем. Самому Беллу належать дві з гіпотез, які ми маємо обговорити. Перша з них — гіпотеза «вікарія з Брея». Фігурально її можна описати так: у світі, де можливі несподівані зміни, корисно продукувати різноманітних потомків: хтось з них може виявитися пристосованим до чогось ще невідомого.
У більшості випадків особини, які продукують потомків, що відповідають наявним умовам, збільшать свій внесок у наступне покоління. Робити певну ставку у розрахунку на малоймовірні зміни — це рішення, яке потребує передбачення. Як ви пам'ятаєте, ми обговорюємо речі, де нема підстав очікувати можливості до передбачення.
А чи може груповий добір підтримати різноманіття ставок, яке передбачається гіпотезою «вікарія з Брея»? На погляд автора цієї лекції — може (і без групового добору цей механізм не запрацює). Це було б непогано перевірити з використанням імітаційного моделювання...

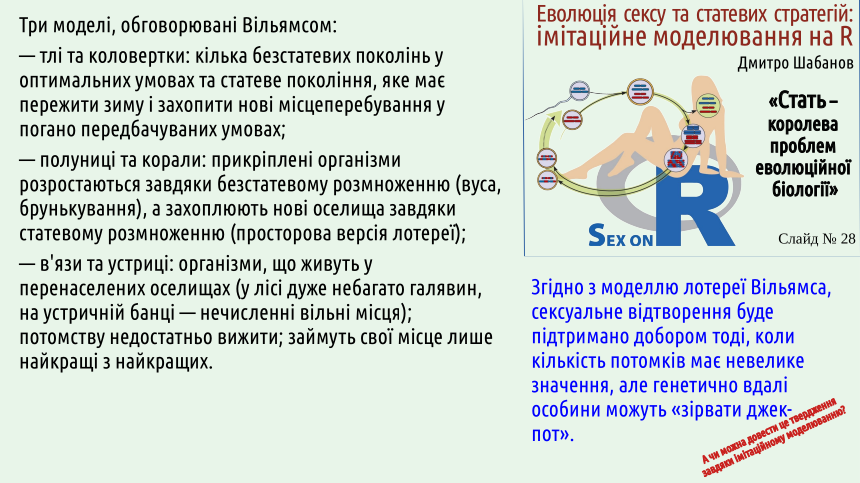
Гіпотеза лотереї Вільямса певним чином розвиває подібний підхід, але з суттєвою різницею. «Вікарій з Брея» передбачає розрахунок на щось, що може статися у майбутньому; якщо мова йде про малоймовірні зміни, їх треба дочікуватися (і дочекатися не вийде, оскільки у передбачливих особин виграють ті, хто будуть збільшувати частку своїх потомків «тут» і «зараз»). Одного з цих недоліків гіпотеза лотереї позбавлена: в ній розглядається пристосування до того, що вже є «зараз», але не «тут».

Критика гіпотези Белла з боку Вільямса частково беззаперечна. Перелічені на наступному слайді моделі переконливі. У двох перших випадках відбувається чергування статевого та клонального відтворення; у третій моделі спостерігається виключно статеве відтворення.
Контраргументам Белла також не можна відмовити у переконливості. Дійсно, припустимо, даревскії (клональні ящірки, яких ми згадували у попередній теоретичній лекції) живуть у досить суворих умовах. Цю гіпотезу Белл назвав з натяком на славнозвісну цитату з важливішої книги Дарвіна.
Знову (як і у випадку «сокири Кондрашова») збереження статевого відтворення пов'язується з міжвидовою конкуренцією. Але... Автору лекції важко уявити, як статева популяція може переграти популяцію, яка не платить «подвійну вартість статі».
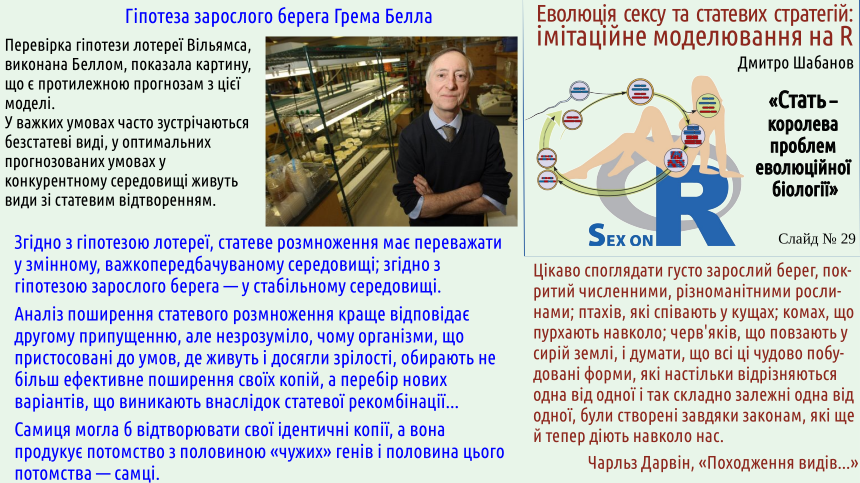
Гіпотеза Менінга стосується механізму, який безумовно вносить певний внесок у стабільність статевого відтворення. Втім... Ані появу статевого відтворення, ані
Цікаво, а можливий статевий добір в популяції гермафродитів? Уявіть собі: кожна особина несе певні маркери своєї потенційної пристосованості і реалізує таку поведінку, за якої вона буде паруватися з особинами, що мають хоча б такий самий (бажано кращий) статус. Втім, якщо складається така ситуація, коли важко знайти партнера, треба обирати: паруватися с тим, хто є, або очікувати принца на білому коні. У роздільностатевих організмів чекати принца має сенс для привабливих самиць; якщо вони погодяться на залицяння сусіда, у разі появи принца на коні може виявитися, що приваблива дівчина вже вагітна і для принца не цікава. А принц може й не шукати виключно принцес; цікава історія з мрійливою дівчиною з периферії не закреслить для нього перспектив з принцесою. Попри певні спрощення у наведеному викладенні, відносини між статями у роздільностатевих організмів базуються на їх принциповій асиметрії (фізіологічні наслідки від успішного парування для самиці та самця є зовсім різними). А що буде, якщо цю асиметрію прибрати, у разі сексу в популяції гермафродитів? Втім, проблему гермафродитів слід детальніше розглянути у наступній теоретичній лекції.

Експериментальне підтвердження робить гіпотезу Менінга набагато вагомішою. «Оздоровчий» ефект статевого добору можна вважати доведеним. Сутність гіпотези — у спробі пояснити статевим добором стійкість роздільностатевості. З цим важче. Певний вплив напевно є, але чи його достатньо?
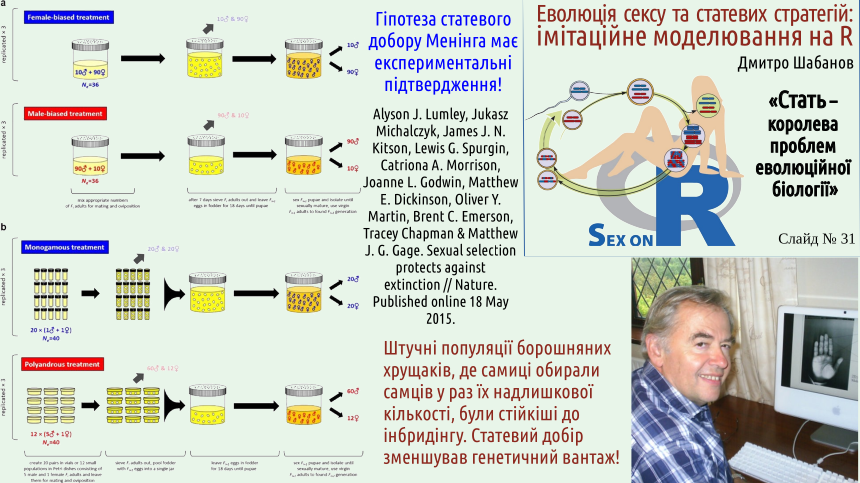
На пострадянському просторі популярними є погляди Вігена Геодакяна, крім іншого — внаслідок здатності цього яркого вченого до пропаганди власних поглядів. Науковий спадок Геодакяна є різнорідним; на погляд автора лекції, в цьому спадку є беззаперечні біологічні помилки. Навіть до цитати, з якої починається наступний слайд, може виникнути багато питань. Втім, багато думок, які пропагував Віген Артаваздович, є безумовно переконливими. Втім, ці ідеї не спростовують теорію статевого добору, а доповнюють її.

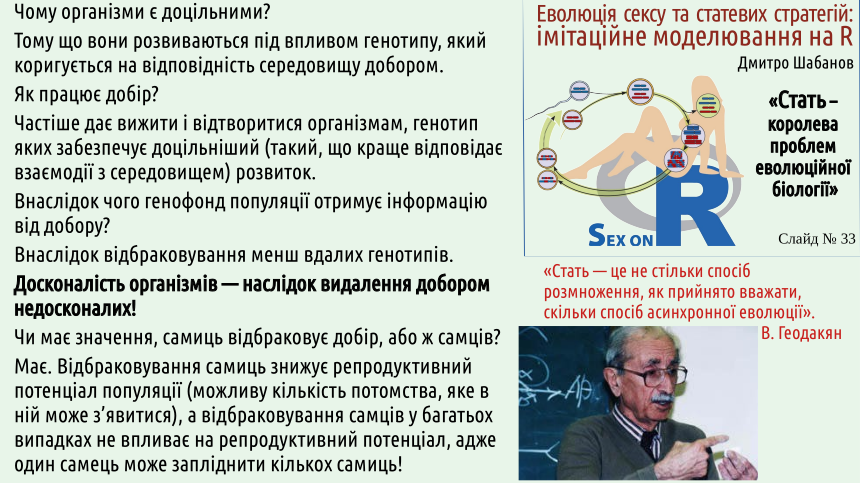
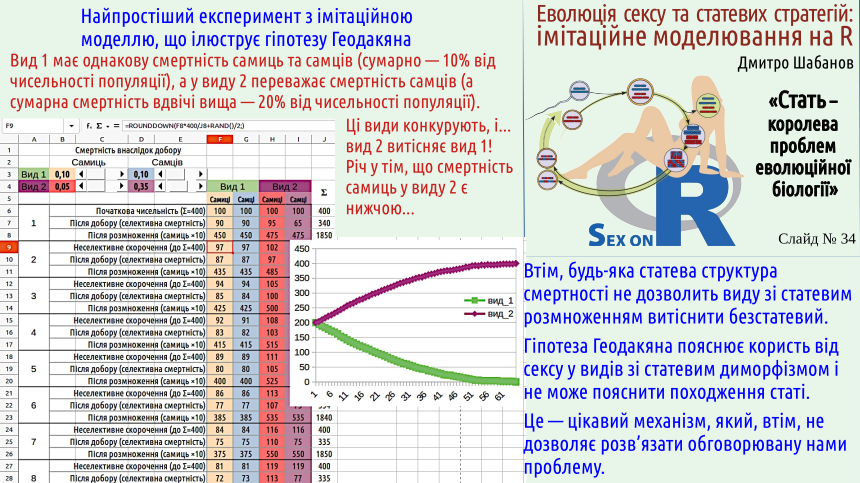
На цьому слайді показано ще одну модель, яка доводить, наскільки суттєвою може бути різниця в плодючості між двома формами, що конкурують. В цій моделі можна змінювати відносну смертність самиць та самців, а після того встановити, як це відбивається на конкуренції між видами. Цю модель можна завантажити або у форматі LibreOffice Calc, або у форматі Microsoft Excel.
Чи може цей механізм пояснити парадокс Мейнарда-Сміта? Ні, адже популяція, що складається з клональних самиць буде відтворюватися ефективніше, ніж будь-яка популяція, де є самці! Чи може ця гіпотеза пояснити появу роздільностатевості? Ні. Щоб цей ефект проявився, необхідно, щоб роздільностатевість та екологічна диференціація статей вже існували!
Ще одна гіпотеза, що отримала експериментальне підтвердження, — це гіпотеза Червоної Королеви (іноді в перекладах — Чорної Королеви) Ли Ван-Валена. Тут ми бачимо відсилання до епізоду з безсмертної казки Льюїса Керролла.
Який механізм може вимагати швидких еволюційних змін? Захист від паразитів, особливо таких, які використовують певні біохімічні відмички для зламу захисних систем своїх хазяїв!
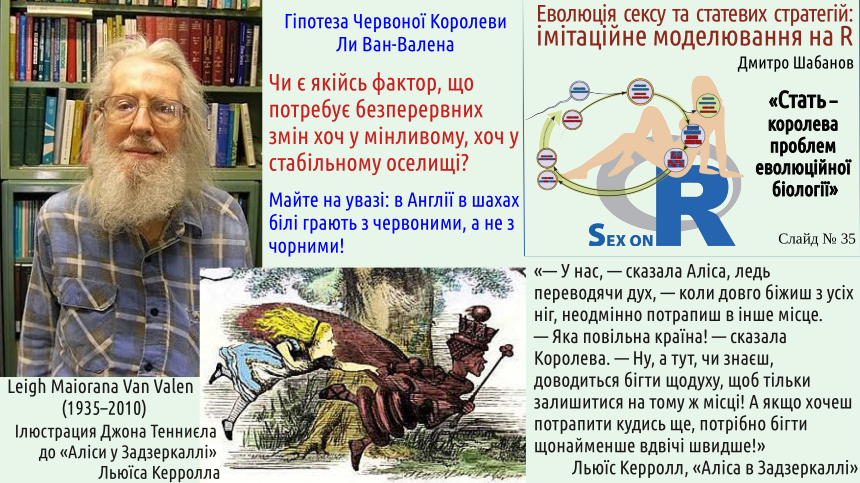
Що відбувалося в експерименті, описаному в наступному слайді? Насамперед, відбувався добір патогенних бактерій. Бактерії здатні швидко еволюціонувати. Захисні системи хазяїв є фільтром, який відбирає таких бактерій, які долають цей захист і можуть ефективно розмножуватися у таких умовах. Якщо черви генетично однакові, бактерії, що пройшли добір в одній особині, вже виявляються оптимально налаштовані до усіх інших. Якщо особини генетично різноманітні (а саме це забезпечує статеве відтворення), ті особини, що відрізняються від хворого, від якого вони заражаються, мають фору. Бактерії погано адаптовані до них, і поки відбувається відносно помірний розвиток інфекції та еволюція паразитів, хазяїн може «навчитися» протистояти загрозі.
Зверніть увагу: у ценорабдитісів гермафродити відтворюються за рахунок самозапліднення. Це не нормальне статеве відтворення, і не типове клональне відтворення. Одна особина продукує й яйцеклітини (з випадковою втратою половини генетичного матеріалу), й сперматозоїди (також з випадковою втратою половини генетичного матеріалу). Запліднення відбувається між яйцеклітиною і сперматозоїдом тієї самої особини. Це призводить до того, що половина генів, за якими батьківська особина була гетерозиготна (мала різні алелі) у потомків стає гомозиготними (будуть мати однакові алелі). Нові алелі (які у випадку справжнього відтворення дає статевий партнер) отримати у такому разі неможливо; таке відтворення гермафродитів призводить до збіднення генетичного різноманіття.
«Червона Королева», безумовно, вносить певний внесок у підтримання статевого відтворення. У випадку, коли виживання популяції критично залежить від паразитарних атак, вплив цього механізму може бути достатньо вагомим. Але чи достатньо його (навіть у разі спільної роботи з іншими механізмами, які ми перелічили), щоб утворити та підтримувати статеве відтворення?
До речі, такі гермафродити, які практикують перехресне запліднення (тобто нормальне сексуальне відтворення без роздільностатевості) повністю використовують усі переваги підтримки «Червоної Королеви»...
6. Відомі й механізми, які визначають недоліки статевого відтворення у порівнянні з клональним. Серед них — «кошмар Дженкіна».
Після того, як ми обговорювали переваги статевого відтворення, можна згадати і про його недоліки. Один з таких недоліків, «подвійну вартість статі», ми вже обговорювали. До речі, перехресно запліднювані гермафродити не «сплачують» цю «вартість». Але як мінімум один недолік сексу (як відтворення з рекомбінацією) стосується і гермафродитів. І зараз нам доведеться розглянути одну історію, яку систематично згадують у спотвореному вигляді.
«Кошмар Дженкіна» — аргумент проти поглядів Дарвіна, який здавався найвагомішим самому Дарвіну.

Автора цієї лекції колись здивувало, що у книгах, де обговорюється «кошмар Дженкіна», не цитують самого Дженкіна. Зазвичай це описували так: «припустимо, на полі білих маків з'явиться червоний мак; він приваблюватиме бджіл, але з його насіння виростуть рожеві маки, потім — блідо рожеві тощо. Але тепер ми знаємо, що колір квітів може залежати від роботи одного гену і спадкуватися як єдине ціле! Як шкода, що Дарвін не поставився уважно до роботи Менделя, який відкрив дискретну спадковість».
З великими труднощами автору цієї лекції вдалося замовити в одній бібліотеці в США копію роботи Дженкіна (зараз усе стало набагато простішим — ця робота доступна у мережі).

Расистські погляди у ті часи були поширеними, майже загальноприйнятними. Обговорювати це зараз — зайве. Цікавіше інше. Ознаки, про які писав Дженкін, очевидно не є такими, які могли б залежати від певного гена. При статевому відтворення ці переваги мали б «розмиватися», втрачатися!
До речі, перехід до клонального відтворення у вдалих гібридів може бути шляхом уходу від «кошмара Дженкіна». Наведемо приклад.
Значна частина сортів плодових культур, які ми вирощуємо у наших садах, є гібридами, які відтворюють клонально. Колись, внаслідок вдалого схрещування, утворився надзвичайно вдалій гібрид, який помітили селекціонери. При насінневому відтворенні видатні спадкові ознаки цього вдалого гібрида втрачаються, розмиваються у повній відповідності до логіки «кошмару Дженкіна». Саме тому такі вдалі гібриди розмножують клонально: живцями, відводками. Насіння, що утворюються на таких рослинах, не висаджують, адже отримані від нього потомки не зберігають видатні якості батьківської особини.
Ймовірно, походження таких клональних видів, як обговорювані в попередній лекції даревскії, також пояснюється подібним механізмом. Внаслідок схрещування (часто — міжвидового) виникає надзвичайно вдалий гібрид. Якщо не відбувається зміни способу його відтворення, він просто зникає — чи то внаслідок порушення утворення статевих клітин, чи до внаслідок «кошмару Дженкіна». Якщо ж такий гібрид переходить до клонального відтворення, його копії поширюються у відповідних умовах і будуть існувати там, поки умови не зміняться (або поки «храповік Мюлера» не зробить свою чорну справу). Здатність до подальшої еволюції у таких гібридів зніжена, але на певний час їх існування буде відносно стійким...
7. Поява сексуального відтворення (за якого вдвічі зменшується ймовірність передачі наступних поколінням будь якого гена), скоріше за все, не може бути пояснена добором на генному рівні. Ймовірно, пояснення обговорюваних феноменів потребує врахування ролі групового добору.
Ще більші теоретичні проблеми пов'язані з поясненням походження сексуального відтворення, самим походженням сексу.
Наведена на наступному слайді цитата Докінза доводить, що він є чесною людиною. З легкої руки Докінза поширилося уявлення, що еволюція є наслідком селекції «егоїстичних генів», виключно результатом добору на генному рівні. Для прибічників Докінза очевидно, що й походження сексу можна пояснити таким добором. Автор цієї лекції вважає, що походження сексу без групового добору пояснити неможливо. А якою є точка зору самого Докінза?
Докінз є прибічником уявлень про виключно роль добору на генетичному рівні. Він розуміє, що перехід від клонального відтворення (за якого кожен ген передається нащадкам з 100% ймовірністю) до сексуального відтворення (за якого гени мають лише 50% шанси на передачу) пояснити генним добором неможливо. Як добір, який має збільшувати пристосованість генів (їх здатність передаватися наступним поколінням) міг привести до зниження їх пристосованості вдвічі? Перед тим, як утворювати статеві клітини, гени ніби-то чемно поступаються своїм конкурентам (з цим пов'язана згадка про гавот) і уходять з половини утворюваних клітин. Здається, це можна пояснити лише добором більш високого рівня. Розуміючи ці проблеми, Докінз просто визнає, що не має відповіді відносно проблеми походження сексу...

8. Найважче пояснити, чому найпоширенішим варіантом серед просунутих організмів не є той, який може здаватися оптимальнішим — гермафродитизм з перехресним заплідненням. Такі гермафродити отримують майже усі переваги статевого розмноження і відтворюються з максимальною ефективністю завдяки тому, що не йдуть на зайві витрати внаслідок «подвійної вартості статі».
Підведемо підсумок лекції. Дев'ять перелічених пояснень переваг статевого відтворення не вирішують парадокс Мейнарда Сміта остаточно, хоча, ймовірно, в цих гіпотезах міститься чимало правильних ідей. Але пояснення проблемі, яка обговорюється на останньому слайді цієї лекції, в них нема. Загалом, пояснення того, чому гермафродити поширені набагато менше, ніж могли би, потребує окремої лекції.