| ← |
«Секс на R» |
→ |
| Секс на R–06: Заготовки для створення R-моделей (R-довідник II) | Секс на R–07: Еволюція та секс Опис проблеми I |
Секс на R–08: «Стать – королева проблем еволюційної біології» (Опис проблеми II) |
Еволюція та секс
Перша лекція, що розглядає еволюційні проблеми сексу, була записана на відео навесні 2022 року і була навіть змонтована, але... Виявилося, що бомбардування Харкова, яке було чуто за вікном, не сприяло якісному запису. Автору цієї лекції його власна важка мова та нерівномірний темп мовлення дуже не сподобалися. У останні дні зими 2023 року автор вирішив не записувати відео, а написати коментарі до слайдів презентації. Може, з часом з'явиться і відеозапис цієї лекції, а поки що студентам доведеться користуватися лекцією в текстовому вигляді.
Цю лекцію хотілося б зробити максимально зрозумілою та коректною. Прошу усіх її читачів повідомляти про побачені недоліки (від граматичних помилок до «темних» фрагментів) і про ідеї та аргументи, що прийшли їм у голову.
Насправді головною темою, яку ми будемо розглядати в цьому курсі, є навіть не секс та стать, а еволюція. Для розуміння того, як саме відбувається еволюція, треба витратити чимало зусиль. Це — тема, якій можна було б присвятити окремий великий курс. Якщо ми почнемо доводити усі твердження, які нам необхідно мати на увазі, ми витратимо весь або майже весь час, відведений на наш курс. Тому доведеться сформулювати певну кількість ключових думок, і запропонувати їх для слухачів курсу. Не погоджуєтеся з чимось — пишіть у телеграм-групу, будемо обговорювати.
У будь-якому разі вам доведеться позбавитися поширених міфів на кшталт «у природному доборі виживає найсильніший», «людина є такою, якою її захотіла створити Природа», «між людиною та твариною — прірва» тощо. Пояснити, чому це маячня?
У природному доборі ймовірніше зберігаються ті, хто краще відповідає своєму способу життя у певних умовах середовища. Сила — зовсім не універсальне розв'язання усіх проблем. Ніякої персоніфікованої «Природи», яка б могла чогось бажати та планувати, нема. Людина є одним з видів тварин; як будь-який вид, вона має певні особливості. Звісно, найважливіші особливості нашого виду стосуються характерного для нас способу вироблення пристосувальної поведінки. Ми — насправді незвичний вид тварин. Крім іншого, в виникненні і розвитку наших особливостей велику роль зіграли особливості нашого сексуального життя і взаємодії між статями.
Наступний слайд є певним змістом, переліком основних думок, які розглядаються в цій презентації...
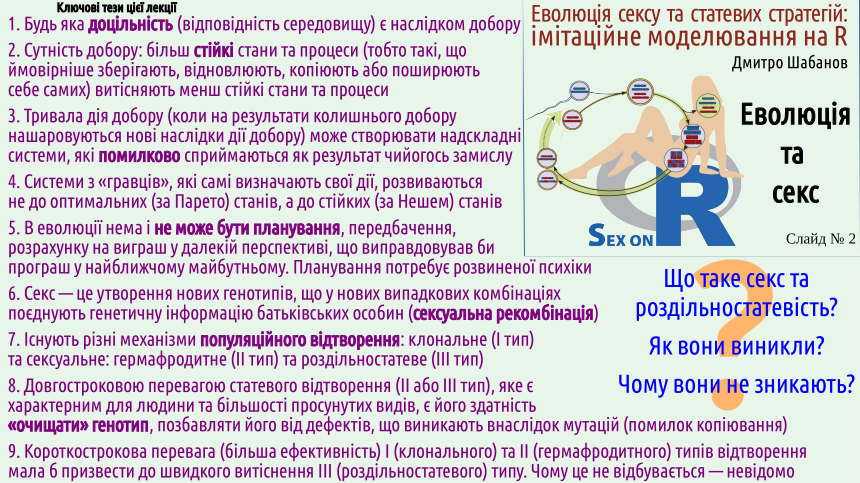
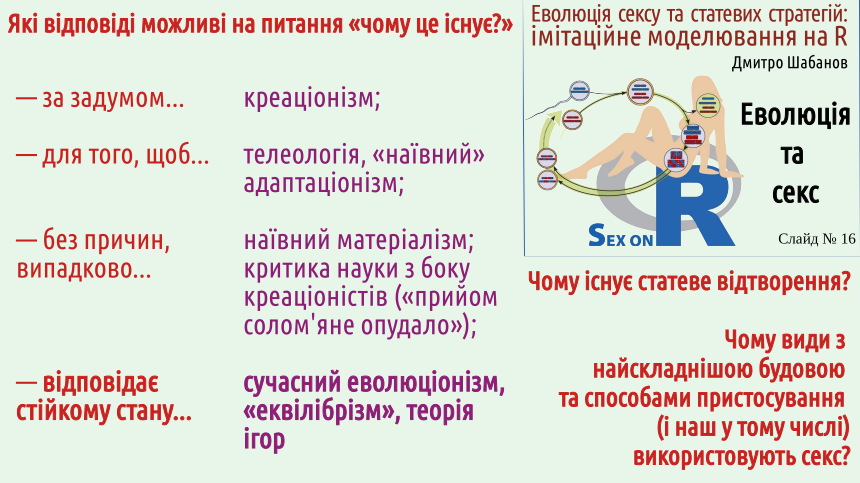
1. Будь яка доцільність (відповідність середовищу) є наслідком добору
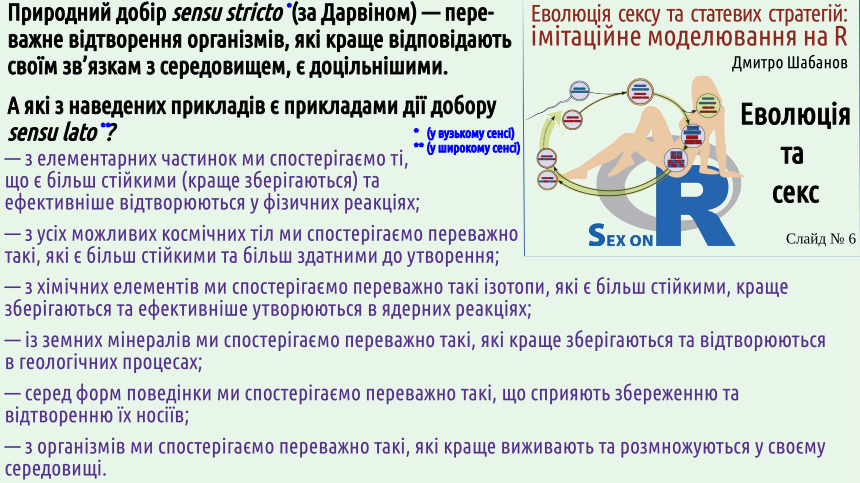
Поширеною є думка, що природний добір — це суто біологічне явище. Доводиться чути такий аргумент проти еволюції: ніби-то оскільки добір відбувається виключно між організмами, саму появу живих організмів добором пояснити неможливо. Звісно, це неправильно. Добір — дуже широкий феномен; його дію можна спостерігати й серед фізичних, і серед біологічних, і серед психічних та культурних явищ (деякі приклади будуть наведені на 6-му слійді). Загалом, теорія інформації вчить, що інформація виникає внаслідок запам'ятовування результатів випадкового вибору. Якщо це запам'ятовування є неселективним (тобто позбавленим добору), виникає сірий шум — тобто хаотичний сигнал, що немає жодної цінності. Селективне збереження інформації (тобто наявність певного добору, перевірка того, що виникло, на ту чи іншу відповідність середовищу) є причиною появи змістовної інформації.
Усі складні, доцільні системи, навіть ті, що уявилися у психіці людини, виникають внаслідок добору. Наприклад, що відбувається у голові людини, яка намагається розв'язати технічну задачу? Припустимо, слід скласти якусь головоломку. Менш досвідчена людина буде намагатися це зробити, крутячи цю головоломку у руках. Так повернути, отак, ще якось спробувати повернути, потягнути, натиснути... Після низки невдалих спроб кінець-кінцем удається підібрати рух, що потрібний для досягнення мети. Що це? Приклад добору. Людина випадково обирала різні рухи, перевіряла їх на відповідність поставленій меті та зберігала (реалізовувала) той варіант, що відповідав цій меті.
Більш досвідчена людина може не робити зайві рухи. Вона дивиться на головоломку (об'єкт зовнішнього середовища), створює у своїй психіці її модель, а далі оперує з цією моделлю. Що буде, якщо зробити це? А це? А до чого приведе такий рух? О, здається, рішення знайдено! Це також добір, але добір випадкового вибору, що перевіряється на моделі.
Насправді в обох випадках перевіряються (чи то у ході маніпулювання з об'єктом, чи то на моделі) не будь-які можливі рухи, а лише відібрані — ті, для яких можна очікувати, що вони можуть привести до бажаного результату. Певний добір є й на цьому рівні, але у будь-якому разі, розв'язання технічної задачі є наслідком добору.
Тут наведені висловлювання трьох видатних мислителів, кожен з яких викликає щире захоплення автора — Больцмана, Тейяра, Деннета. Це дуже різні люди: класик термодинаміки, ієзуїт-палеонтолог, філософ. Чому усі вони вважають ідею добору та еволюції настільки важливою?

На могилі Больцмана не стали писати, хто він такий; над його скульптурним зображенням навели формулу, що пов'язує ентропію (міру невпорядкованості, S) з термодинамічною ймовірністю, W. В більш сучасному записі це виглядало б так: S = k × ln(W). k — це стала Больцмана, 2,07 × 10−21 Дж. W можна як кількість мікростанів системи, що відповідає тому її макростану, що розглядається. Загалом, Больцману вдалося перекинути місток між мікроскопічними та макроскопічними станами досліджуваних систем. Больцман був генієм, що значно випередив свій час і не був зрозумілим сучасниками; викликана цим криза призвела до самогубства цього вченого.
Одним з найважливіших результатів Больцмана стало те, що йому вдалося визначити, які зміни можуть відбуватися самовільно (крім іншого, цей напрямок самовільних змін визначає напрям, у якому тече час). Ізольовані системи змінюються у напрямку зростання їх ентропії, тобто переходять з менш ймовірних станів у більш ймовірні. Ви ж зрозуміли, що термодинамічна ймовірність стану системи визначається кількістю мікроскопічних способів, якими можна отримати даний стан?

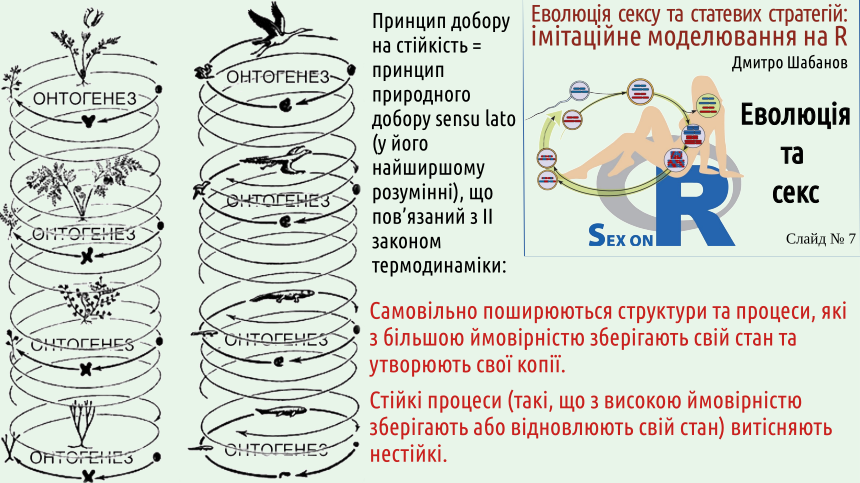
2. Сутність добору: більш стійкі стани та процеси (тобто такі, що ймовірніше зберігають, відновлюють, копіюють або поширюють себе самих) витісняють менш стійкі стани та процеси
Ще один поширений (і невірний!) аргумент проти добору полягає в тому, що, відповідно до другого закону термодинаміки у трактуванні Больцмана ізольовані системи мають еволюціонувати у напрямку збільшення невпорядкованості, а живі організми є напрочуд упорядкованими. Здається, що еволюція суперечить другому закону...
Ні, вона йому відповідає! Другий закон вимагає збереження більш термодинамічно ймовірних станів. Інакше можна сказати, що зберігаються більш стійкі стани. У самому широкому сенсі природний добір можна розглядати як процес, що переважно зберігає більш стійкі стани, і переважно руйнує менш стійкі стани різноманітних систем. На слайді наведено певну кількість прикладів переважного збереження більш стійких систем та процесів.
Якщо розглядати життєві цикли організмів, які реалізуються у певних умовах середовища, виявляється, що стійкими є ті цикли, що з більшою ймовірністю відтворюються у даних умовах (тобто породжують більш пристосовані організми), а ті організми, що відтворюються з меншою ймовірністю, є менш пристосованими. Стійкими є такі організми, що більш ефективно зберігають, відновлюють, копіюють або поширюють себе самих!
Середовище, у якому відбуваються життєві цикли організмів, не є ізольованою системою: це відкрита система, через яку тече потік енергії. Добір, який є наслідком другого закону термодинаміки, підтримує у таких умовах більш стійкі життєві цикли, що відповідають більш пристосованим організмам.
Одним з видатних дослідником самоорганізації у відкритих системах, через які тече потік енергії, був Ілля Пригожин. Він показав, що в так званих дисипативних системах (сильно нерівноважних системах, які розсіюють енергію, яка через них тече) можуть самовільно виникати складно організовані структури. Це — наслідок зростання загальної невпорядкованості, якої вимагає другий закон термодинаміки. Процеси, у яких діє природний добір за Дарвіном, є прикладом такої самовільної самоорганізації.

3. Тривала дія добору (коли на результати колишнього добору нашаровуються нові наслідки дії добору) може створювати надскладні системи, які помилково сприймаються як результат чийогось замислу
Повернемося до першої тези. А чи може бути так, що доцільність виникає не внаслідок добору, а виключно у разі випадковості? Розглянемо ще один поширений аргумент проти еволюційної біології. Наводять ніби-то розрахунки ймовірності випадкової появи амінокислотної послідовності певного білку, чи, припустимо, ймовірність того, що молекули, які утворюють живу клітину, випадково поєднаються у таку структуру, яку ми спостерігаємо. Звісно, це вигадки. Випадково може виникнути результат, що потребує однократної випадковості.
Розглянемо простий приклад. Знаєте таку гру: один ховає якійсь предмет, а інший шукає його, пропонуючи ті чи інші рухи, і отримуючи відповіді «гаряче» або «холодно». До речі, іноді той, хто шукає предмет, зчитує несвідомі реакції партнера і завдяки порівнянню реакцій на різні рухи визначає, де знаходиться прихований «скарб». Якщо у гравців вистачить терпіння, таким чином можна передати дуже складні інструкції. Може бути так, що спочатку треба шляхом проб та помилок отримати розуміння, у яке приміщення треба потрапити; потім (тим самим шляхом) — який об'єкт обрати в цьому приміщенні, потім — що слід зробити з цим об'єктом... Результати проб та помилок (=добору), якими встановлюються наступні дії, нашаровуються на результати проб та помилок, що привели гравця у потрібне місце.
А можна обійтися без добору? Ви загадали певне однозначне число, я зробив спробу його вгадати, сказав, припустимо, «3» і... вгадав! Тут був добір? В усякому разі, коли ми встановили, що я вгадав (тобто перевірив свою здогадку, перевірили пробу на відповідність середовищу, тобто задуманому вами числу), ми вже використали процедуру добору.
Саме тому жива клітина не виникла внаслідок того, що молекули випадково «зістрибнулися» потрібним чином. Вона є наслідком тривалого, «багатошарового» добору. Як це могло відбуватися — проблема, яку можна обговорювати, але це занадто далеко уведе нас від теми нашого курсу.
Цікаво, що наша інтуїція не пристосована для розпізнавання результатів «багатошарового» добору. Коли ми дивимося на складні системи, що є наслідком тривалої дії добору, ми сприймаємо їх як щось заздалегідь сплановане. Річ у тому, що наш інтелект еволюційно сформований не як засіб для досягнення істини, а як засіб пристосування до наявних умов. Краще побачити задум там, де його не може бути, ніж не побачити його у тому разі, коли він є (наприклад, у разі необхідності зрозуміти наміри хитрого противника).

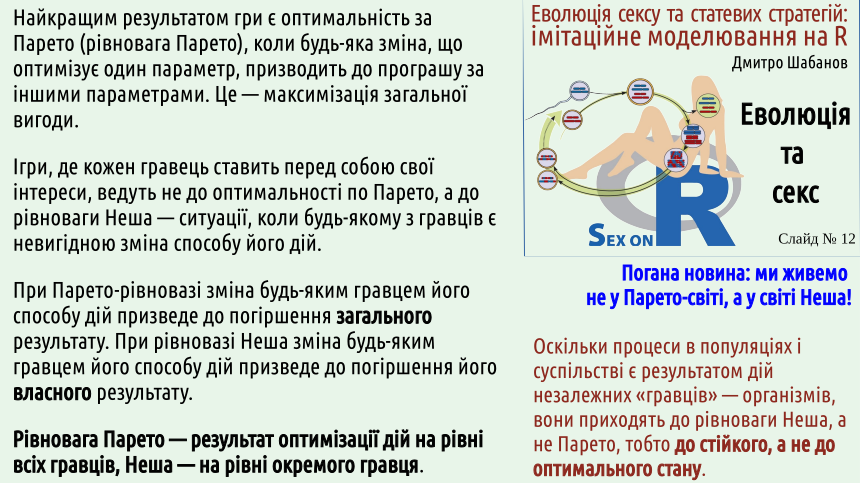
4. Системи з «гравців», які самі визначають свої дії, розвиваються не до оптимальних (за Парето) станів, а до стійких (за Нешем) станів
Вільфредо Парето описав, який стан системи з кількох гравців є оптимальним для усіх. Це — ситуація максимальної спільної вигоди. Світ, побудований за Парето, був би оптимальним. У разі, коли ми маємо справу з системами, де можливе планування, можна досягати Парето-станів.

На жаль, якщо гравці є незалежними, вони діють так, щоб максимізувати власну вигоду, незважаючи на те, як їх дії впливають на інтереси інших гравців. Вперше це показав Джон Неш.

На відміну від Вільфредо Парето, якій, скоріше за все, був дуже гарною людиною, Неш хворів на шизофренію і був досить важкою у спілкуванні персоною. Світ, що живе за Парето, був би набагато кращим, ніж світ, що розвивається за Нешем.
...На жаль, ми живемо у світі Неша...
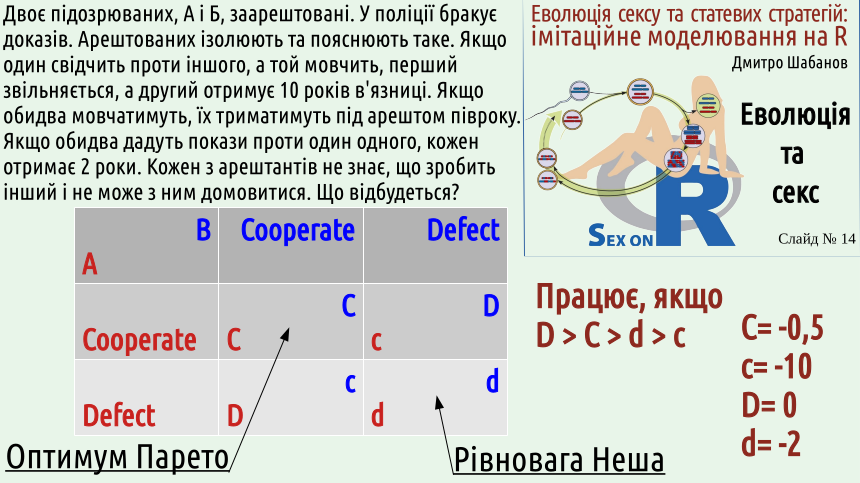
Чому гравці обирають рівновагу Неша, а не оптимум Парето? Тому що вони віддають перевагу власним інтересам. Це можна показати на прикладі славнозвісної «дилеми в'язня». Її формулювання наведено на цьому слайді. «Дилему в'язня» було сформульовано у 1950 році, але слідчі почали використовувати у своїй діяльності набагато раніше.

Максимізації загальної вигоди, оптимуму Парето відповідає випадок, коли обидва в'язні зберігають лояльність один одному. Рівновазі Неша відповідає випадок, коли обидва в'язні «здають» один одного. ЧОМУ?
Поставте себе на місце одного з в'язнів. Припустимо, ваш подільник мовчить. Якщо ви дасте покази, вас звільнять, а якщо будете мовчати, сидітимете півроку. Таким чином, якщо ваш партнер мовчить, вам вигідніше давати покази.
Інша ситуація: ваш подільник дав покази проти вас. Якщо ви дасте покази, ви сидітимете два роки, а якщо мовчатиме — усі десять! Таким чином, якщо ваш партнер дає покази, вам вигідніше давати покази.
В обох випадках вам вигідніше давати покази! Ну що, розповідайте усе слідчому, і будьте впевнені, що ваш подільник зробить те ж саме!
5. В еволюції нема і не може бути планування, передбачення, розрахунку на виграш у далекій перспективі, що виправдовував би програш у найближчому майбутньому. Планування потребує розвиненої психіки
Коли долю системи з кількох гравців вирішує одна людина, що здатна до планування та передбачення наслідків своїх дій, є надія, що результатом розвитку такої системи стане оптимум за Парето. Але навіть у разі, коли ми розглядаємо дії незалежних гравців, навіть у тому разі, якщо вони можуть планувати та передбачати майбутнє, система потраплятиме у пастку за Нешем. Сумно... Зверніть увагу: це стосується навіть еволюції систем, що складаються зі здатних до передбачення гравців!
А що відбуватиметься з системами, де нема здатних до планування і передбачення персон, тобто будь-яких фізичних та біологічних систем, крім тих, де визначальну роль грає поведінка високоорганізованих тварин, таких як люди? Там нема навіть підстав сподіватися на врахування можливих наслідків процесів, які відбуватимуться. Еволюція йде не до оптимуму за Парето, вона йде до рівноваги Неша!
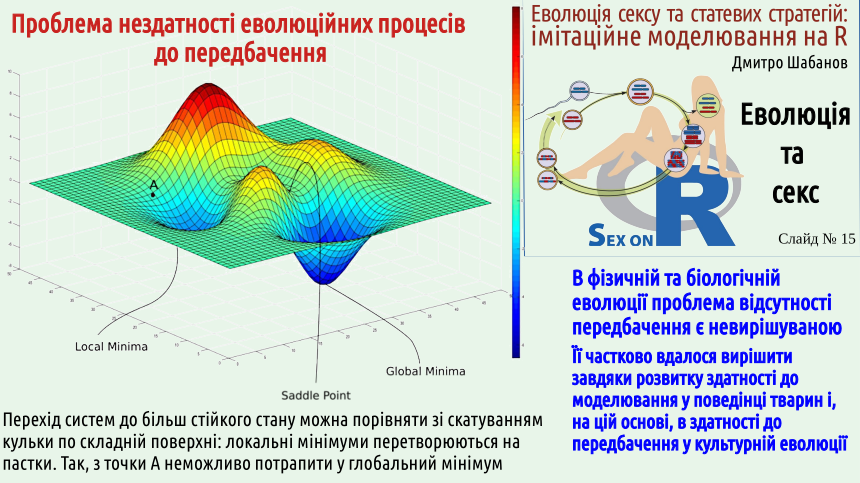
Так що, досягти оптимуму за Парето загалом неможливо? Можливо, коли ми маємо справу з системою, що централізовано керується інтелектом, який здатний до передбачення. Розвиток психіки високорозвинений тварин пішов у напрямку вдосконалення моделей середовища. Користуючись цією моделлю, тварина може побудувати прогноз (згадайте приклад з ґавою!) і діяти відповідно до нього. Людина обігнала в цій здатності усі інші види і навіть створила штучний інтелект. В психіці людини, як ми вже обговорювали, працює певна форма добору; досить часто добір використовується і в роботі штучного інтелекту. Наприклад, для пошуку глобальних мінімумів у випадках, подібних до того, що показано на попередньому слайді, алгоритми починають оптимізацію з випадкових положень на досліджуваній площині, а потім визначають, яка спроба привела до найкращого результату.
З викладеного є цікавий наслідок. Досліджуючи наслідки еволюції недоцільно задаватися «дитячим» питанням «навіщо так зроблено». Ті речі, про які розповідав Метт Рідлі, процитований на 9-му слайді, загалом не є наслідком будь-якого планування! Зрозуміло, вони й не є наслідком чистої випадковості, без добору випадковість не може утворити нічого складного. Якщо ми намагаємося зрозуміти, чому існують певні феномени, нам слід встановити, як і чому виникають стійкі стани, наслідком яких є існування таких феноменів.
Усе наведене було вступом (прелюдією, якщо бажаєте) до нашої основної розмови. Важливо, щоб розглядачи сексуальне відтворення, ви задавалися не питанням «для чого?», а питанням «чому?». Чому існує секс?
6. Секс — це утворення нових генотипів, що у нових випадкових комбінаціях поєднують генетичну інформацію батьківських особин (сексуальна рекомбінація)
Ми закінчили вступну розмову про еволюцію і нарешті переходимо до розгляду сексу!
Почнемо з визначень. Ми вже говорили про це, але і в цій лекції слід дати ці визначення. Студенти, які прийшли на наш курс для того, щоб навчитися отримувати більше задоволення від занять сексом, можуть бути розчаровані: задоволення, яке можна отримати від наших занять, носить переважно інтелектуальний характер.
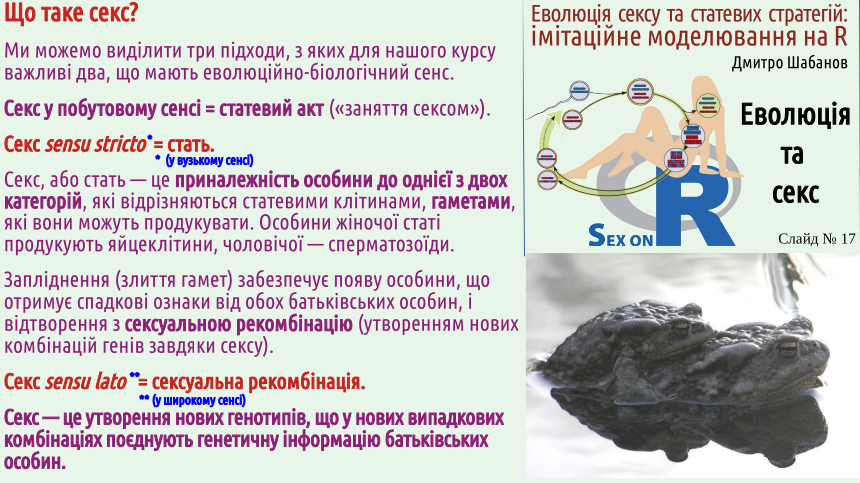
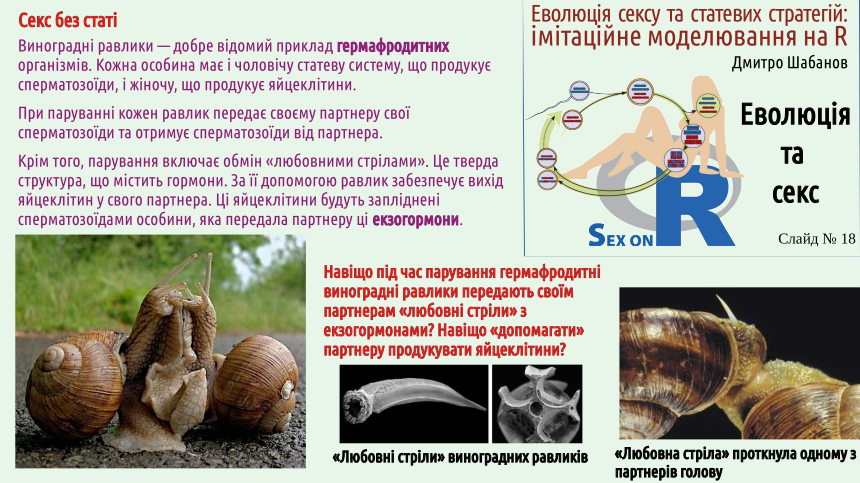
Вам здається, що секс нерозривно пов'язаний зі статтю? Поруч з нами існує чимало істот, що практикують секс, не належачі до жіночої або чоловічої статі. Це не лише равлики, що показані на слайді, та дощові черви. Значна частина покритонасінних (квіткових) рослин є однодомними, тобто такими, що продукують і пилок, і насіння (тобто фактично є гермафродитами). Не будемо зараз заглиблюватися у дискусію, чи мають гермафродити стать (на погляд автора лекції — ні, але деякі його колеги з цим не погодяться). Можна згадати ще й інші приклади. Один з них такий. Зелена водорость спірогіра (Spirogyra) є однією з поширеніших водоростей прісних водойм; вона часто бере участь у «цвітінні» води. Тіло спірогіри — це нерозгалуджена нитка. Дві таки нітки наближуються, і відбувається кон'югація. Вміст клітини з однієї нитки перетікає в клітину іншої нитки і зливається з нею. У заплідненні беруть участь не гамети, а просто клітини! Утворюється спора (зигоспора — спора, що утворюється внаслідок запліднення), яка зимує і наступного року проходить мейоз.
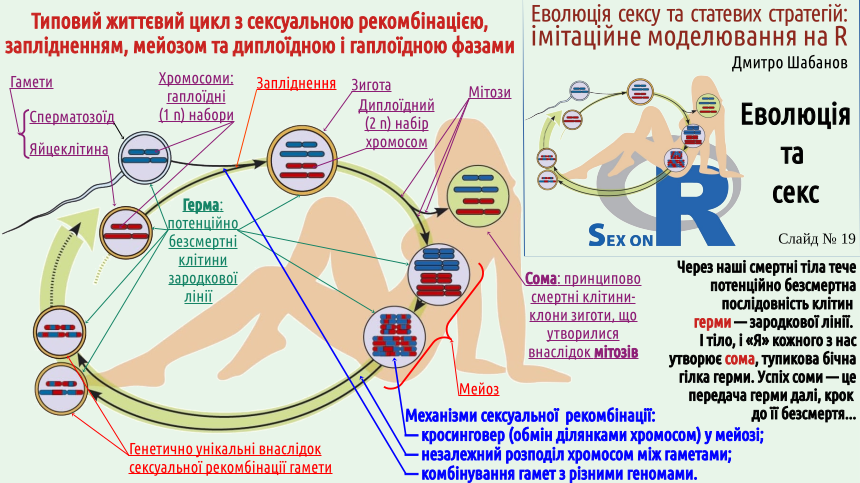
Переважна кількість поділів клітин у життєвому циклі людини (його можна розглядати як типовий варіант життєвого циклу з сексуальним відтворенням) є мітозами, тобто такими поділами, наслідком яких є утворення двох генетично ідентичних (клональних) клітин. Майже усі клітини нашого тіла є клонами (генетичними копіями) зиготи, заплідненої яйцеклітини. В таких клітинах є по комплекту хромосом, отриманих від матері, і по комплекту хромосом отриманих від батька. Ці клітини утворюють сому, власне тіло. Втім, сама зигота не є частиною соми; вона належить до сукупності потенційно безсмертних клітин, зародкової лінії або герми. Більшість нащадків зиготи утворює сому, але деякі з них стають клітинами, що утворюють гамети (статеві клітини). В тілі молодої жінки є кілька сотень тисяч клітин, які можуть розвинутися у жіночу гамету, яйцеклітину. Ці клітини-попередниці утворилися ще тоді, коли майбутня жінка ще була плодом і перебувала всередині матки своєї матері. Під час дорослішання та старіння жінки кількість клітин-попередниць яйцеклітин незворотно зменшується. За усе життя жінки в її організмі може утворитися кілька сотен яйцеклітин — зовсім небагато, якщо порівнювати цю кількість з кількістю сперматозоїдів.
Чоловікам потрібно продукувати величезну кількість гамет. Зрілий чоловік за своє життя може утворити кілька трильйонів сперматозоїдів (згадайте, що сучасна чисельність людства нещодавно перевалила за 8 мільярдів). Явище сексу пов'язано саме з формуванням гамет. Під час утворення гамет (гаметогенезу) кількість генетичного матеріалу в клітинах зменшується вдвічі. Це забезпечує особливий тип клітинного поділу — мейоз. У ході мейозу відбувається випадковий обмін фрагментами материнських і батьківських хромосом (у ході процесу, що названий кросинговером). Наслідком цього є те, що усі гамети є генетично унікальними.
Вам простіше буде повірити, що жінка не має двох однакових яйцеклітин, адже внаслідок кросинговеру і незалежного розподілу між дочірніми клітинами гібридних хромосом, що утворилися в результаті кросинговеру, кожна клітина отримує свій варіант генетичної інформації. Роздивіться схему на слайді: там умовно показаний цей процес!
А якою є ймовірність того, що з трильйонів сперматозоїдів, що утворив чоловік, два виявляться генетично однаковими? Вона є меншою, ніж одиниця, що поділена на кількість атомів у Всесвіті! Саме це різноманіття і є результатом сексуальної рекомбінації (утворення нових сполучень генетичної інформації).
Якщо ви роздивитеся слайд, ви побачите, що різні особливості життєвого циклу організмів з сексуальною рекомбінацією є пов'язаними. Запліднення подвоює кількість генетичної інформації, і тому у мейозі її кількість має зменшитися вдвічі. Механізм цього скорочення є таким, що він викликає утворення генетично унікальних гамет; ймовірнісний характер поєднання гамет є додатковим джерелом генетичного різноманіття організмів, утворених внаслідок запліднення.
7. Існують різні механізми популяційного відтворення: клональне (І тип) та сексуальне: гермафродитне (ІІ тип) та роздільностатеве (ІІІ тип)
Настав час запропонувати класифікацію способів відтворення організмів, якою ми будемо користуватися. Ми будемо відрізняти розмноження (те, що відбувається з організмами) та відтворення (явище, що є характерним для популяційного рівня біологічних систем). Відтворення популяцій відбувається завдяки розмноженню організмів... Чому ми розглядаємо не розмноження організмів, а відтворення популяцій? Тому що еволюціонують саме популяції, адже вони є потенційно безсмертними, а організми є принципово смертними.
Так ось, популяції бувають клональними (відтворення в яких відбувається без сексуальної рекомбінації) та менделевськими (названими на честь Грегора Менделя, який зробив величезний внесок у вивчення сексуального відтворення). На схемі, крім популяцій, показані геміклональні популяційні системи — біосистеми, що можуть виникати внаслідок особливої форми міжвидової гібридизації. Ці системи є об'єктом досліджень автора даної лекції, і прибирати їх з схеми він не хотів би. Втім, у даному курсі ми не будемо розглядати геміклональні популяційні системи. Нас будуть цікавити популяції I, II і, особливо, III типів.
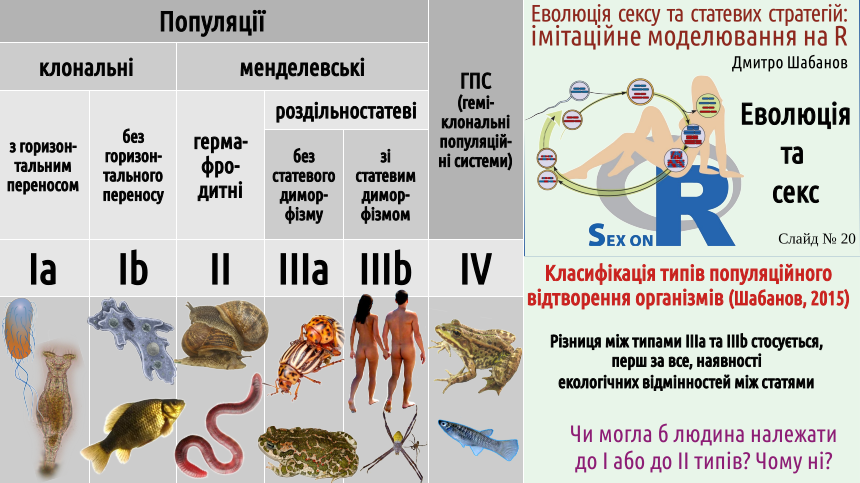
Як ви бачите, на класифікації типів відтворення показані і підтипи головних типів. Межа між цими підтипами є більш розмитою, ніж межа між типами. Усе ж таки, слід врахувати, що клонально відтворювані організми можуть більшою або меншою мірою практикувати те, що може бути певним аналогом сексу — передачу генетичної інформації від організму до організму. Цей процес відбувається набагато менш контрольовано, ніж у разі справжнього сексу, і має назву горизонтального переносу генетичної інформації (вертикальною передачею вважають передачу від предків потомкам). Скоріше за все, здатність до горизонтального переносу є типовою для популяцій I типу; випадки, коли ця здатність втрачається є, скоріше за все, тупиковими.
Серед різностатевих популяцій можна виділити такі, в яких самиці та самці ведуть приблизно однаковий спосіб життя, і такі, де екологічні ролі представників двох статей є достатньо різними. Різниця екологічних ролей статей стає причиною статевого диморфізму — різниці між статями, що напряму не пов'язана з розмноженням. Статевий диморфізм є характерним і для людини, але, наприклад, у багатьох павуків він є більш вираженим.
Після цього вступу ми можемо порівняти названі типи відтворення. Пам'ятаєте Робінзона Крузо, героя славетного роману Даніеля Дефо, написаного на початку XVIII століття? З цієї книги почався класичний англійський роман... Так ось, Робінзон був моряком, що потрапив на безлюдний острів і прожив на ньому 28 років. Завдяки своїй працелюбності від побудував цілком придатний для життя мікросвіт. До речі, історії людей, що опинялися на безлюдних островах у справжньому світі, були не настільки успішними, як історія вигаданого Робінзона; людина є біологічною істотою, що пристосована до функціювання у складі популяцій. Але навіть для літературного героя головною проблемою стала його самітність.
Як добре буле б для Робінзона Крузо, якби популяції людини відтворювалися б за I типом! Навіть самотня особина могла б утворити певну кількість своїх генетичних потомків. Для ізольованої особини такий спосіб відтворення став би перевагою. Звісно, клональні потомки з'являлися б позбавленими одягу з хутра, рушниці і навіть бороди з вусами, але копіювання зображення Робінзону на слайді допомагає показати, що його потомки у разі клонального розмноження були б генетично ідентичними до нього.
Через багато років життя на самоті, Робінзон спостерігає, як на острів прибувають дикуни-канібали, які збиралися з'їсти полонених, яких вони привезли з собою. Робінзон рятує полоненого, якого називає П'ятницею. З часом П'ятниця стає вірним компаньйоном Робінзона.
Уявіть собі, що людина б відтворювалася за II типом. У гермафродитів дві будь-які особини, що перебувають на відповідній стадії свого життєвого циклу, можуть стати один одному статевими партнерами та залишити потомство. На відміну від I типу відтворення, у такому разі це потомство буде генетично різноманітним. На слайді це різноманіття показане комбінуванням ознак Робінзона та П'ятниці.
Це ж дуже незручно: потенційним партнером для людини (серед особин відповідного віку) може бути лише половина особин! З ймовірністю близькою до 1/2 дві випадково обрані особини з популяції III типу позбавлені можливості спільного розмноження!
Хиба це не дивно, що наш спосіб розмноження є таким «незручним»?
8. Довгостроковою перевагою статевого відтворення (II або III тип), яке є характерним для людини та більшості просунутих видів, є його здатність «очищати» генотип, позбавляти його від дефектів, що виникають внаслідок мутацій (помилок копіювання)
Давайте розділимо кілька питань.
Перше — чому людина виникла серед істот зі статевим відтворенням. Відповідь на це питання відома, і зараз ми її обговоримо.
Друге — чому людина є саме роздільностатевою, відтворюється за III типом. Відповідь на це питання є більш складною, але у автора є гіпотеза, що пояснює цей феномен. Ми обговоримо цю проблему в наступній присвяченій проблематиці нашого курсу лекції. Цікавим інструментом встановлення того, наскільки ця гіпотеза є логічною, є імітаційне моделювання.
Третє — чому людина зберігає роздільностатеве відтворення. Загальноприйнятої відповіді на це питання нема; за думкою автора для відповіді на це питання слід відмовитися від поширеної в сучасній еволюційній біології точки зору. Для дослідження цієї проблеми також буде дуже корисним імітаційне моделювання.
Розуміючи, що вичерпна відповідь на перше питання не вирішує друге та третє, почнемо з початку...
Чим складнішою є система, тим більше в неї можливостей поламатися. Для того, щоб утворити таку складну істоту, як людина, необхідно, щоб її розвиток відбувався під керуванням дуже складного і досконалого геному (комплексу спадкової інформації). Давайте у самому загальному вигляді замислимося, що впливає на еволюцію геному?
Розглядаючи будь-який геном, ми можемо побачити, що він ускладнюється внаслідок добору і деградує внаслідок мутаційного процесу (випадкових змін). Людина є досконалою, тому що в її спадковій інформації міститься концентрований досвід виживання та розмноження усіх її предків, що існували на нашій планеті протягом майже 4 мільярдів років!
Замислитеся: ми знаємо, що ваші батьки (мати та батько) успішно дожили до статевозрілості та успішно залишили як мінімум одного потомка. Не усім людям, не усім організмам загалом так щастить! Кожен з вас може зрозуміти, що його батьки є переможцями у природному доборі, яка б доля не чекала на них після того, як вони дали вам життя. Але якщо є переможні, мають бути й переможені. Це — представники покоління ваших батьків, що не дожили до статевозрілості або не залишили нащадків.
Переможцями у природному доборі є не лише ваші батьки, а й усі четверо ваших бабусь та дідусів. В їх поколінні також були переможені; до речі, крім тих, хто не дожили до необхідного віку або не розмножилися, поразки у природному доборі зазнали й ті, хто лишили потомків, які кінець-кінцем загинули або не розмножилися. Той, хто програє у природному доборі, не лише стає переможеним сам; від відіймає пов'язану з його існуванням перемогу у всіх своїх предків!
Переможцями у доборі є усі вісім ваших прабабусь та прадідусів, усі шістнадцять ваших прапрабабусь та прапрадідусів, усі 32..., 64..., 128..., 256..., 512..., 1024...
Зараз ми живемо у світі, де до статевозрілості доживає більшість новонароджених, але ще кілька поколінь тому ситуація була зовсім іншою. Ми — нащадки величезної черги переможців! І тепер можна пояснити, чому ми це згадали. Геноми ваших предків відрізнялися від геномів їхніх однолітків. Деякі з цих відмінностей підвищували шанси на перемогу у природному доборі. Звісно, іноді особини з блискучими задатками програвали, а носії генетичних недоліків вигравали. Втім покоління за поколінням добір зберігав у геномах генетичні задатки, що сприяли перемозі (тобто ті, що підвищували доцільність, відповідність організмів їх способу життю у їх середовищі), а секс «тасував» ці задатки та ставав причиною перевірки на досконалість нових їх сполучень.
Ми описали процес, що є причиною вдосконалення нашого геному. На жаль, безперервно діє і зворотній процес, що веде його до деградації. Система копіювання генетичної інформації діє з фантастичною ефективністю, але й вона неминуче робить помилки. Наслідки цих помилок має відсіювати негативний добір: їх носії мають ставати переможцями у доборі з меншою ймовірністю, ніж особини, що позбавлені таких недоліків. Чим більшим є геном, тим більше в ньому помилок, тим більш ефективним має бути добір, що очищує від них геном. Як виміряти ефективність такого очищувального добору? Через кількість особин, що відсторонені від розмноження, тобто тих, хто програє у доборі. Чим більшим є геном, тим більше особин у кожному поколінні має програвати у природному доборі, щоб підтримувати його у порядку!
Ви розумієте, завдяки якому механізму наручники можуть лише затягуватися на руках арештованого, і не можуть самі собою вивільнити його руки? Цей механізм має назву храповика. Зараз зрозумієте, для чого ми його згадали...
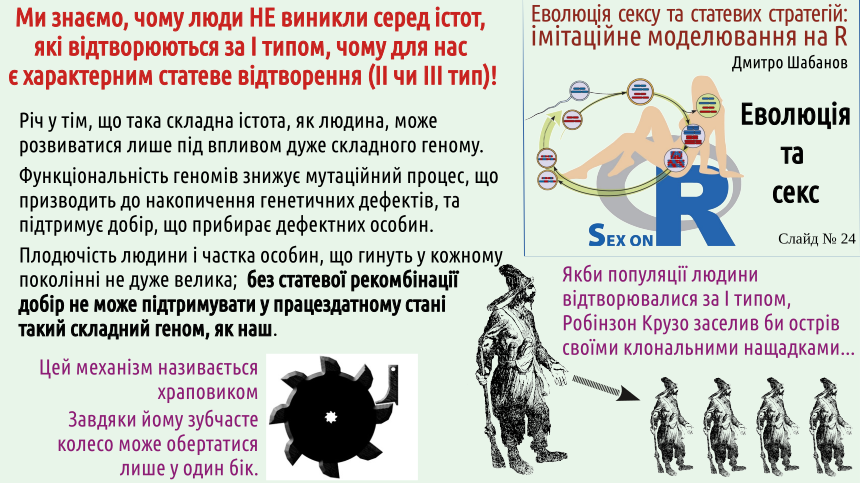
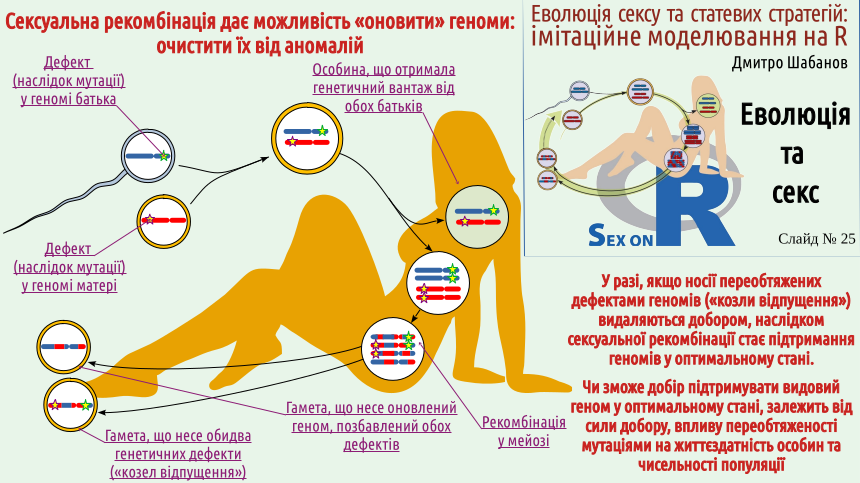
Повернемося до схеми сексуального відтворення (ту саму, що розташована й на логотипі нашого курсу). Розглянемо випадок, коли і хромосома матері, і хромосома батька несуть по дефекту — наслідку мутації. Кожна клітина соми буде нести обидва дефекти, але серед гамет з'являться як такі, що будуть нести по одному дефекту, так і такі, що мають обидва чи на мають жодного! Якщо обговорювані дефекти знижують шанси їх носіїв на перемогу у природному доборі, носії обох недоліків (їх можна порівняти з біблійними «козлами відпущення») з більшою ймовірністю зникнуть з популяції. З іншого боку, в популяції з'являться й особини, що позбавлені обох дефектів, що очищені сексом!
А що буде відбуватися в популяціях, що відтворюються без сексу? Розглянемо це на прикладі клональних ящірок, що поширені на Кавказі (а також у штучній популяції, яка існує на Південному Бузі в Україні). Зараз відомо кілька видів таких ящірок, які пізніше були виокремлені у рід Даревскія, названий на честь Іллі Сергійовича Даревського (1924–2009), який відкрив механізм їх відтворення. Даревського вважають російським вченим, хоча він був киянином і вчився у Київському університеті до того, як переїхав у Ленінград.
Одним з головних досягнень Даревського стало відкриття (у середині XX століття) партеногенезу скельних кавказьких ящірок. Мова йде про механізм розмноження самиць, яке не потребує участі самців. Це — приклад I типу популяційного відтворення, причому — вторинного. Предки даревській звісно були роздільностатевими тваринами, але на певному етапі своєї еволюції вони перейшли до партеногенезу. Усі їх клітини, і ті, що утворюють сому, і ті, що утворюють герму, діляться лише шляхом мітозу.
Нема сексу — нема й його очищувальної дії на рівні геномів! Генетичної рекомбінації (утворення нових сполучень генів) нема, і генетичні дефекти, що отримала даревскія, не можуть нікуди подітися (ймовірність зворотної мутації, що поверне мутантну ділянку хромосоми у нормальний стан є критично низькою і цій ймовірністю можна просто знехтувати). Мутації можуть призвести лише до збільшення кількості дефектів, що будуть нести партеногенетичні яйцеклітини даревскії. Ці партеногенетичні яйцеклітини не потребують запліднення; особини, що розвинуться з них, у кращому разі отримають усі генетичні дефекти, які несла материнська особина, а у гіршому разі отримають ще якісь нові дефекти на додаток до старих.
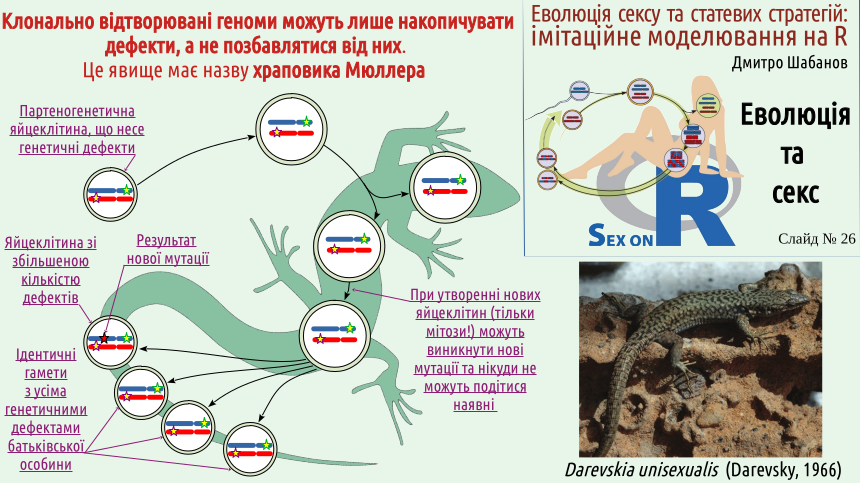
Тепер ви можете зрозуміти, чому Герман Мюллер, який описав це явище, порівняв його з храповиком. У разі відсутності сексуальної рекомбінації кількість генетичних дефектів може змінюватися лише у один бік — у бік збільшення їх кількості, як храповик, який може обертатися лише в одному напрямку.

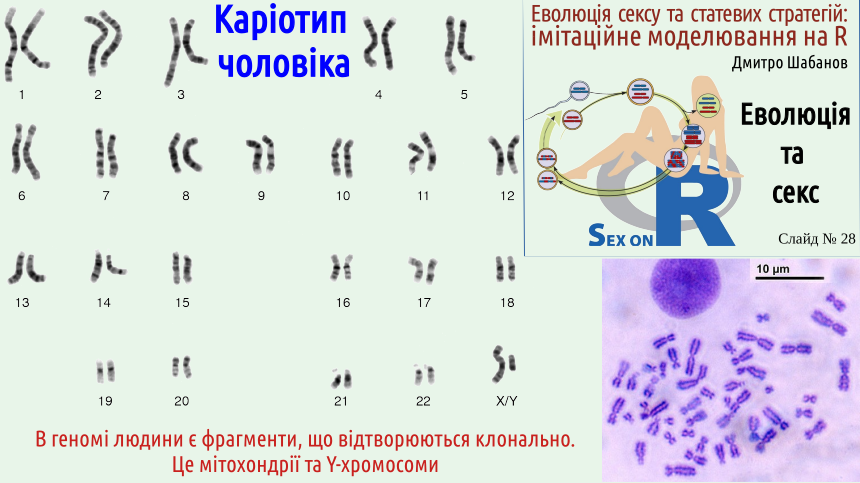
Наведені міркування можуть здаватися відірваними від дійсності... Розглянемо приклад, що стосується нашого геному. На наступному слайді показано каріотип (хромосомний набір) людини. Як у багатьох інших видів тварин стать людини визначає пара статевих хромосом. Статеві хромосоми бувають двох типів. X-хромосома є відносно звичайною хромосомою — не найбільшою, не найменшою. Ця хромосома несе достатньо велику кількість генів, робота яких впливає на низку характеристик особини. На відміну від неї, Y-хромосома є, по суті, уламком X-хромосоми, на ній працює набагато менша кількість генів, з яких головним є один: ген SRY (англ. Sex-determining Region Y). Це ген, функцією якого є перемикання розвитку на чоловічий тип. З заплідненої яйцеклітини, що має генотип XY (тобто несе одну X-хромосому, і одну Y-хромосому) розвинеться чоловік, тому що на певному етапі ембріонального розвитку Y-хромосома ввімкне роботу усіх генів, що роблять людину чоловіком. З яйцеклітини з генотипом XX розвинеться жінка. У чоловічі гамети, сперматозоїди, може потрапити і X-, і Y-хромосома; у жіночі гамети, яйцеклітини, потрапляють лише наявні у жінки X-хромосоми. Стать дитини визначає, яку статеву хромосому несе сперматозоїд, адже яйцеклітина у будь-якому разі принесе X-хромосому (звісно, ми розглядали тут типовий випадок; відомі й різноманітні аномалії, що пов'язані з відхиленням від нього).
Ви розумієте, що на наступному слайді зображено саме каріотип чоловіка, а не жінки, адже у складі цього каріотипу є і X-, і Y-хромосома. Але тут постає цікаве питання: чому чоловіча статева хромосома виглядає як уламок жіночої?
Як показано на логотипі нашого курсу, для того, щоб між материнською та батьківською хромосомами відбувся кросинговер, вони мають наблизитися одна до одної, а для цього необхідно, щоб вони одна одну «впізнали». Генетичні тексти, що відповідають один одному, розташовуються поруч, а потім обмінюються фрагментами. У чоловіка та інших типових ссавців X- та Y-хромосоми «впізнають» одна одну, але обмін ділянками на більшій їх частині майже повністю заблоковано. Почнемо з початку: з появи статевих хромосом ссавців.
Близько 150 мільйонів років тому ген фактору транскрипції SOX3, що був розташований на X-хромосомі, подвоївся. Одна з його копій змінилася і перетворилася на ген SRY. Цей ген перебрав на себе запуск розвитку за чоловічим типом. Добір підтримав розташування поруч з геном ген SRY інших генів, що були необхідні для формування чоловічої статі — наприклад, генів, що контролюють продукування сперми та деякі інші особливості самців. Якщо кросинговер розбивав цей «чоловічий» комплекс, його ефективність знижувалася. Тому добір підтримав просту аномалію, яка заважає кросинговеру: інверсію, обертання ключової ділянки Y-хромосоми на 180°. Хромосоми в цілому впізнають одна одну, а протилежно спрямовані ділянки — ні.
У самиць з генотипом XX сексуальна рекомбінація підтримувала функціональний стан X-хромосом. У чоловіків в інвертованій ділянці Y-хромосоми рекомбінація виявилася суттєво пригніченою, адже Y-хромосома зустрічається в парі з X-хромосомою, де генетичні послідовності є протилежно спрямованими. Якщо в X-хромосомі з'являвся дефект, він видалявся внаслідок сексу (тобто сексуальної рекомбінації). Якщо у Y-хромосомі з'являвся некритичний дефект, він там і залишався, а необхідні функції забезпечували гени X-хромосоми. Після того, як добір припинив підтримувати функціональний стан Y-хромосомних генів, мутаційний процес швидко їх зруйнував (ви ж пам'ятаєте, що функціональний стан генетичної програми потребує її постійної підтримки добором?).
А що відбувається з Y-хромосомою, якщо мутації пошкодять той самий ген SRY, що запускає чоловічий тип розвитку? Таке явище відомо і має назву синдрому Сваєра. Хворі на цей синдром зовнішньо виглядають як жінки, але не мають сформованих статевих залоз. Звісно, потомства вони не залишають, ефективно видаляючи з популяції дефектний варіант гену SRY.
У рідкісних випадках кросинговер між важливими для розвитку статі ділянками X- та Y-хромосомами усе ж таки відбувається. Внаслідок цього, ген SRY може потрапити на X-хромосому, але й ця аномалія не буде мати довгострокових наслідків. Що буде, якщо у особини з жіночим генотипом, XX, одна (чи обидві) хромосоми будуть нести ген SRY? Розвинеться особина, що зовнішньо виглядає як чоловік та є безплідною; це має назву назву синдрому де ля Шапеля. Внаслідок цього, коли X-хромосома з непритаманним для неї фрагментом потрапить у генетичну жінку, безплідність цієї жінки забезпечить видалення зміненої хромосоми з генофонду популяції.
У чому принципова різниця між більшістю генів на Y-хромосомі і тим самим, що є головним перемикачем статевого розвитку? Якщо дефекти перших не є критичними, вони відфільтровуються добором неефективно. «Поламки», що призводять до погіршення продукування сперми, будуть прибиратися добором навіть у разі дії храповика Мюллера. А ось дефект перемикача статевого розвитку SRY буде прибиратися добором цілком ефективно.
На жаль, останню думку не розуміє велика кількість людей. Ось, подивіться на фрагменти однієї російської науково-популярної статті (на жаль, такі міркування можна знайти й на інших мовах). Не дуже проникливі біологи, які зареєстрували зменшення кількості функціональних генів на Y-хромосомах (але не розуміють, як працює добір), намагаються вирахувати, коли зникне й останній ген. А до чого це призведе? До зникнення чоловіків, тобто, до вимирання людей або до перетворення жінок на «амазонок», які будуть відтворюватися «неприродним» чином! А-а-а-а, на нас чекає катастрофа!
Звісно, принципова різниця між наслідками мутацій різних генів на Y-хромосомі є причиною того, що процес їх «вимкнення» не має йти з постійною швидкістю. Y-хромосома стає достатньо простою, щоб її функціональність могла підтримуватися добором і без сексуальної рекомбінації.
Тепер ви зрозуміли, чому людській геном в цілому потребує сексуальної рекомбінації для своєї еволюції, а значно простіша Y-хромосома (поламки якої призводять до її вибраковування добором) — ні? Але, згадайте, на початку цієї, 8-ї частини лекції ми зазначили три різні питання, що стосуються характерного для людини типу популяційного відтворення? Друге питання залишимо для подальшого обговорення, а до третього (чому людина зберігає роздільностатеве відтворення) перейдемо прямо зараз.
9. Короткострокова перевага (більша ефективність) I (клонального) та II (гермафродитного) типів відтворення мала б призвести до швидкого витіснення III (роздільностатевого) типу. Чому це не відбувається — невідомо
Майже пів століття тому Мейнард Сміт (це таке подвійне прізвище, а ім'я у цього видатного англійського біолога — Джон) вказав на одне величезне питання, що пов'язане з різностатевістю. Роздивіться схему, що показана на жовтому тлі. У роздільностатевих організмів потомство безпосередньо залишають тільки самиці. Самці необхідні для цього процесу, але дитинчат не народжують. Наслідком є явище, яке Мейнард Сміт назвав «подвійною вартістю статі» або «ціною самців».
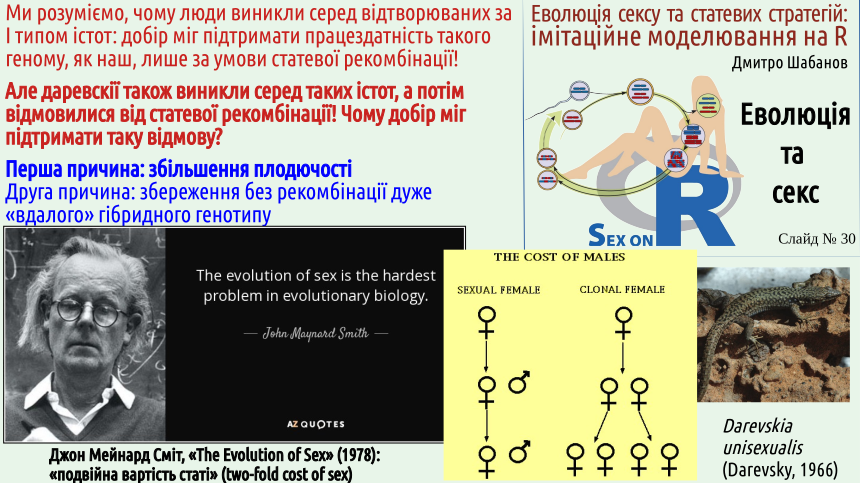
Обставина, на яку звернув увагу Мейнард Сміт, породжує низку питань. Як роздільностатевий вид може витримати конкуренцію з подібним видом, що відтворюється клонально, адже ефективність розмноження клонального виду є вдвічі більшою?
Розглянемо дві поширені відповіді на це питання. Перша. Роздільностатевий вид буде використовувати усі переваги сексуального відтворення, а клональний вид програє внаслідок дії храповика Мюлера. Цей аргумент не працює внаслідок двох обставин. По-перше, популяції гермафродитів не примушені нести витрати на «подвійну вартість статі». У гермафродитів безпосередньо може залишати нащадків будь-яка особина, і при цьому будуть працювати усі переваги сексуальної рекомбінації. По-друге, навіть клональний вид спочатку мав би виграти у свого роздільностатевого конкурента, а вже потім відчув би несприятливі наслідки дії храповика Мюлера. Популяційна динаміка, міжвидова конкуренція — відносно швидкі процеси, їх наслідки проявлять себе вже в короткостроковій перспективі. Дегенерація геномів внаслідок дії храповика Мюлера проявить себе лише у довгостроковій перспективі, а як ви пам'ятаєте, у еволюції передбачення нема.
Друга відповідь. Може бути так, що швидкість розмноження що у клонального, що у роздільностатевого виду, які конкурують, є надлишковою; ніби-то у такому разі зменшення ефективності розмноження у роздільностатевого виду вдвічі лише піде йому на користь. Але ж навіть у тому разі, коли більша частина потомства гине, у природний добір у разі однакової чисельності плідників вступить вдвічі більша кількість клональних потомків. Якщо усі молоді особини будуть мати однакові шанси на перемогу у природному доборі, це призведе до того, що кількість клональних організмів у наступному поколінні збільшиться.
Але «парадокс Мейнарда Сміта» можна сформулювати ще в більш дивній формі. Завдяки спостереженням за багатьма видами ми знаємо, що в популяціях роздільностатевих видів іноді з'являються партеногенетичні самки, такі, як ми можемо спостерігати у ящірок даревскій. Поява таких самок — перехід від III типу відтворення до I типу. Можна припустити, що такий перехід є можливим для багатьох роздільностатевих популяцій. Згідно з простими розрахунками, клональна самка, що з'являється всередині роздільностатевої популяції, за дуже невеликий час має витіснити своїх роздільностатевих родичів. Так, у довгостроковій перспективі потомство такої самиці зазнає несприятливого впливу дії храповика Мюлера. Втім, як демонструє приклад тих самих даревскій, клонально відтворювані організми усе ж таки можуть існувати протягом достатньо тривалого часу. Для перемоги у внутрішньопопуляційній конкуренції достатньо набагато меншого часу.
Сформулюємо «парадокс Мейнарда Сміта» так: чому поява партеногенетичних самиць в популяціях роздільностатевих організмів не призводить до швидкої відмови від сексуального відтворення?
Щоб продемонструвати, як може відбуватися така відмова, використаємо імітаційну модель, але не на мові R, яку ми будемо використовувати в більшості моделей нашого курсу, а набагато більш просту, створену за допомогою електронних таблиць. Цю модель можна завантажити, хоч у форматі Calc (електронних таблиць у складі вільного пакету LibbreOffice, в якому ця модель була зроблена), хоч у форматі Excel (електронних таблиць у складі Microsoft Office). На відміну від моделей в електронних таблицях, які вчаться будувати студенти кафедри зоології та екології тварин, в цій моделі навіть не використовується здатність електронних таблиць проводити розрахунки за введеними у таблиці формулами. Тут простір таблиці використовується просто як лист паперу: на ньому вказано, які розрахунки проводяться. Розглянемо цю модель детальніше.
Ми імітуємо популяцію, що складається з 20 особин. Спочатку це 10 самиць і 10 самців. На кожному циклі кожна самиця приносить по 2 потомка (самицю і самця). На другому циклі одна з самиць, що з'являється у ході розмноження, виявляється клональною (партеногенетичною). Після розмноження на кожному циклі сумарна чисельність усієї популяції скорочується приблизно до 20 особин. Усі групи особин скорочуються в однаковий пропорції. Оскільки скорочення чисельності проводиться за правилами округлення, іноді можуть лишатися, наприклад, 19 чи 21 особина.
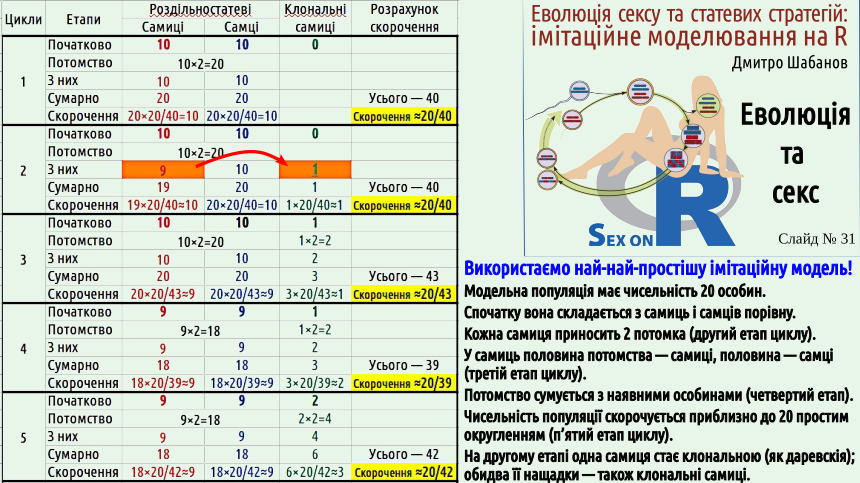
Та ж сама процедура повторюється 15 раз. Ми бачимо, що партеногенетичні самиці відтворюються швидше, а скорочуються так само, як роздільностатеві особини. На початок 16 циклу в популяції лишаються лише партеногенетичні самиці...
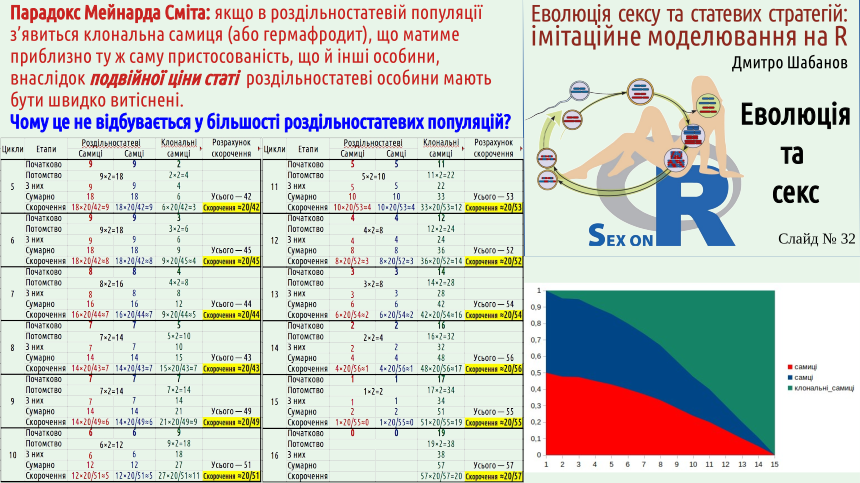
Зверніть увагу на графік в лівому нижньому куті останнього слайду цієї лекції. Він показує частку самиць, самців та клональних самиць в нашій модельній популяції.
Чому ж подібне не відбувається у більшості роздільностатевих популяцій?
Не повірите: відповіді на це питання до цього часу нема. Є низка аргументів, які треба враховувати. Є припущення, які суперечать поширеній точці зору. Це — одна з проблем, яку було б дуже цікаво дослідити за допомогою імітаційного моделювання.
Далі буде...