Подкласс Lepidosauromorpha ныне насчитывает более 7 тысяч видов ящериц и змей и является второй после птиц самой богатой видами группой тетрапод. Термин Lepidosauromorpha ввел в употребление М. Бентон в 1983 г. Представителей лепидозавров, как и примитивных архозавроморф, считают примитивными диапсидами: в настоящее время Archosauromorpha + Lepidosauromorpha составляют Sauria и Sauria + Younginiformes составляют Neodiapsida (Laurin, 1991). Вероятно, Lepidosauromorpha отделились от Archosauromorpha в середине перми.
Лепидозавры произошли от ящерицеобразных лепидозавроморфных таксонов, которые процветали во время поздней перми и раннего триаса. Во время раннего триаса эти разнообразные формы породили такие специализированные формы как kuehneosaurs, завроптеригии (возможно) и настоящие лепидозавры. Все эти рептилии распространялись, занимая освободившиеся в конце пермского вымирания экологические ниши, прежде занимаемые преобладавшими ранее зверообразными рептилиями. Лепидозавры возникли как небольшая группа с акцентом на небольшие ниши, которые не были заполнены архозаврами.
Характерными чертами лепидозавроморф являются:
- неуклюжая осанка с извилистыми движениями (примитивная характеристика, свойственная начальным тетраподам);
- ацент на боковых волнообразных изгибах позвоночника;
- жесткая грудная клетка, большая грудина;
- нет двуногой специализации;
- запястья и кисти, а также шейные позвонки немодифицированы (по аналогии с примитивными диапсидами);
- перекрывающаяся чешуя.
Прогресс лепидозавров можно объяснить эволюцией стрептостилии (передне-заднего движения квадратной кости) и мобильности челюстей и черепа в целом. Стрептостилия у лепидозавров достигается путем частичной или полной потери мощной нижне-височной дуги. Диапазон изменений нижне-височной дуги диапсид и, в частности, лепидозавров отражен на рисунке:
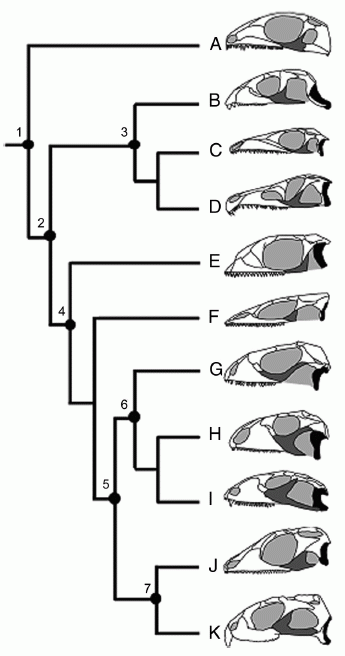
Рис. 1. Упрощенная филогения диапсидных рептилий, показывающая вариации в равзвитии нижней височной дуги.
Черным показана квадратная кость, серым - скуловая. (A) диапсида Petrolacosaurus; (B) ранний ринхозавр Mesosuchus; (C) архозавроморф Prolacerta; (D) архозавроформ Proterosuchus; (E) лепидозавроформ Kuehneosaurus; (F) ранний лепидозавр Marmoretta; (G) сохранившееся чешуйчатое Iguana, отражающее более-менее неспециализированную форму; (H) огромный травоядный меловой сцинкоморф Polyglyphanodon, (I) позднемеловая ящерица Tianyusaurus, уникальная среди чешуйчатых ввиду акинетической ринхоцефалальной квадратной кости, (J) ринхоцефал Gephyrosaurus; и (K) ныне живущий ринхоцефал Sphenodon. Пометки: 1, Diapsida, примитивная нижне-височная дуга, фиксированная квадратная кость, крепкий эпиптеригоид; 2, Sauria, нет нижней височной дуги, фиксированная квадратная кость и эпиптеригоид; 3, Archosauromorpha, нет нижней височной дуги в базальных таксонах, но развитая у Rhynchosaurs и Archosaurs; 4, Lepidosauriformes и 5, Lepidosauria, нет нижней височной дуги в базальных таксонах, квадратная кость, эпиптеригоид срастаются с птеригоидом, большая чешуйчатая кость; 6, Squamata, перекрывание квадратной кости и птеригоида уменьшено, эпиптеригоид связан с птеригоидом, нижняя височная дуга развита только у Tianyusaurus only; и 7, Rhynchocephalia, нет нижней височной дуги в базальных таксонах, развитая только у некоторых потомков, квадратная кость, птеригоид и эпиптеригоид крепко зашиты. Из Moazen et al, 2009
В состав Lepidosauromorpha входит инфракласс Лепидозавры (Lepidosauria) с надотрядами Чешуйчатые (Squamata) и Клювоголовые (Rhynchocephalia или Sphenodontida). Сфенодонты были очень распространены и успешны с конца триаса до поздней юры, но постепенно уменьшились в течении мелового и третичного периодов. В то же время, чешуйчатые, которым были редкими или неизвестными до средней юры, до сих пор являются весьма успешными после прогрессирования в мелу и кайнозое. Разница между этими группами лепидозавров состоит в том, что они имеют различные адаптивные стратегии, у одной группы развивающие сильный, неподвижный, приспособленный для мощного укуса череп, а у другой - череп с пониженной силой и высокой мобильностью. Последнее приспособление получили змеи, которые могут открыть рот чрезвычайно широко для того, чтобы проглотить крупную добычу.
Ископаемые находки лепидозавроморф.
Наиболее раниие предполагаемые лепидозавтры - поздне пермские Lanthanolania (Россия, Modesto and Reisz, 2002) и Saurosternon (Южная Африка, Carroll, 1975).
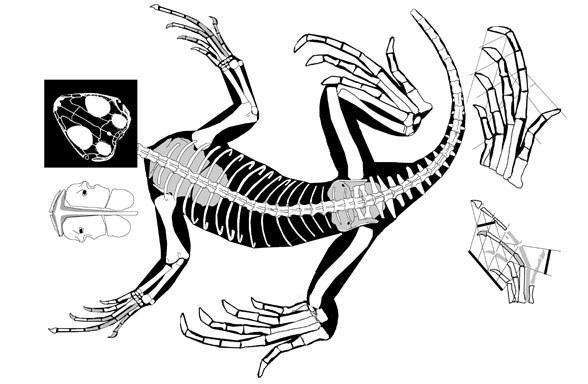
Saurosternon bainii (Huxley 1868)
Paliguana (ранний триас, Южная Африка, Carroll, 1975) представлен одним поврежденным черепом с огромной квадратной костью, соответствующей признакам лепидозавроморфов. Paliguana относят к Eolacertilia, куда также относится группа Marmoretta.
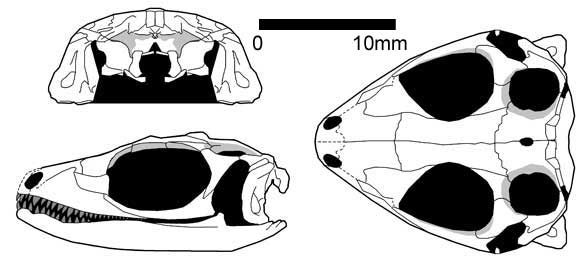
Paliguana whitei (Broom 1903)
Кроме этого, останки, относимые к раннетриасовым лепидозавроморфам, были найдены в Польше (Czatkowice, Borsuk-Białynicka
et al., 1999). Эти отложения сформировались в засушливой среде с локализированными водоемами, фауна включала рыб, темноспондилов, амфибий, проколофонов и отдельных архозавроморфов.
Kuehneosauria. Кюнеозавры являлись специализированными, длинно-ребристыми планерами, известные из раннего триаса Польши и позднего триаса Англии и Северной Америки. Интересной особенностью кюнеозаврид были необычно длинные летательные перепонки, по крайней мере вдвое превышающие их размеры.
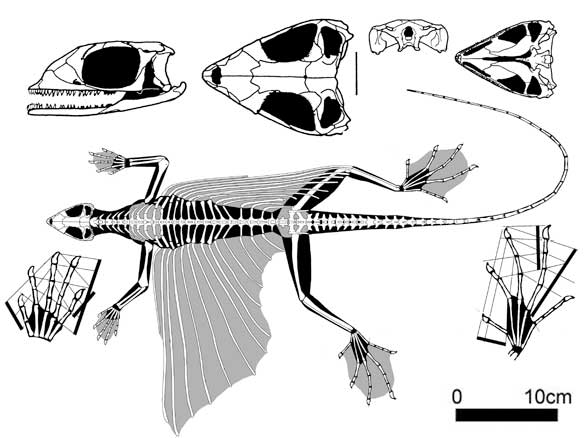
Kuehneosaurus latus (Robinson 1962), поздний триас, Англия
Кюнеозавры были способны планировать на кожной перепонке, натянутой на 10-11 парах подвижных ребер, сочлененных с платицельными позвонками. Особенно длинные ребра имел североамериканский Icarosaurus siefkeri с черепом длиной всего 25 мм, «летные» ребра у него были четко изогнуты:

Icarosaurus siefkeri Colbert, 1966
Кюнеозавры жили на небольших влажных прибрежных морских островах вместе с птерозаврами, арозавроформами, ринхоцефалами и редкими млекопитающими (Robinson, 1962; Fraser, 1994).
К другим мезозойским лепидозавроморфам, не относящимся к лепидозаврам, относятся среднетриасовый Coartaredens (Англия, Spencer and Storrs, 2002) и Megachirella (Италия, Renesto and Posenato, 2003); раннеюрский Tamaulipasaurus (Мексика, Clark and Hernandez, 1994). Филогенетическое родство Megachirella и Tamaulipasaurus остается неразрешенным.

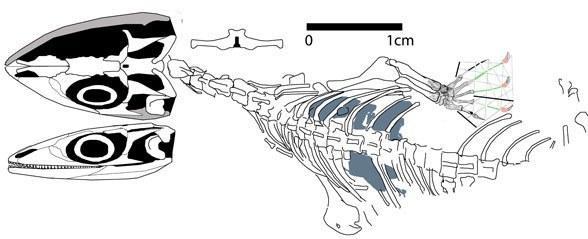
Ископаемая и реконструированная Megachirella wachtleri (Renesto and Posenato 2003).
Tamaulipasaurus morenoi (Clark and Hernandez 1994).
Надотряд Rhynchocephalia (Sphenodonta)
Клювоголовые - настоящие диапсиды некрупных размеров. Ископаемые остатки ринхоцефалов относятся к более 40 таксонам в диапазоне от позднего триаса до нынешнего времени.Географическое распространение: Европа, Северная и Южная Америка, Китай, Индия, Марокко, Южная Африка и Новая Зеландия.
Настоящие ринхоцефалы появляются в нижнем триасе Южной Африки (Palacrodon) и, возможно, Восточной Европы, но достигают расцвета в верхнем триасе Западной Европы, откуда описывают более десятка родов сфенодонтид. Одна форма описана из Кении. Из нижней юры Индии по обломкам челюстей описано два рода ринхоцефалов. В средней юре Мексики появляются уже сфенодонты, близкие к современной гаттерии, в том числе карликовый Zapatodon [Reynoso, Clark, 1998] и Cynosphenodon. Сфенодонтиды известны из верхней юры Северной Америки (три рода) и Западной Европы. Из мела известны только немногие сфенодонты. В кайнозое испокаемые ринхоцефалы неизвестны, но ныне существует новозеладнский ринхлцефал гаттерия - Sphenodon punctatus.

Sphenodon punctatus Evans, 1980
В экологическом отношении мезозойские клювоголовые были разнообразны. Они включали длиннотелых морских пловцов (плеврозавры, сафеозавры, Кэрролл and Wild, 1994), грацильных бегунов (Homoeosaurus, Cocude-Мишель, 1963), бронированных Pamizinsaurus (Reynoso, 1997) и группу большетелых рептилий с копытообразными когтевыми фалангами (Priosphenodon, и Apesteguia Novas, 2003). Среди них были насекомоядные формы (Gephryosaurus; Diphydontosaurus), оппортунистические "хищники" (Sphenodon, Dawbin, 1962; Cree и др., 1999), предположительно ядовитые хищники (Sphenovipera, Reynoso, 2005, известный из средней юры Мексики), а также специализированные травоядные животные (например, Toxolophosaurus, Priosphenodon, Apesteguía и Novas, 2003). Несмотря на то. что некоторые их характеристики менее продвинутые, чем у ящериц, некоторые примитивные их черты являются вторичными. Сфенодонты могут оставаться активными при более низких температурах, чем ящерицы. Также для клювологовых характерен длинный репродуктивный цикл.

Рис. 2. Сфенодонты
Характерные особенности строения сфенодонтов: акродонтные зубы с замедленным замещением, полная нижняя височная дуга, неподвижная квадратная кость (монимостилия), укороченные шейные позвонки, хвостовые позвонки зачастую поделены надвое автотомальной септой, облегчающей произвольную автотомию (отрыв хвоста). При питании пользуются движениями нижней челюсти.
Отряд Pleurosauridae. Из морской юры и нижнего мела Западной Европы известны два или три рода специализированных водных плеврозавров. Они имеют акродонтные зубы и длинное туловище и превышающий длину тела хвост. У нижнеюрского Palaeopleurosaurus в теле было только 37 позвонков, в то же время плеврозавр Pleurosaurus – крупнейших не только из плеврозавров, но и ринхоцефалов в целом (длина – 1,5 м).

Palaeopleurosaurus posidoniae

Pleurosaurus goldfussi
Надотряд Squamata
Чешуйчатые насчитывают более 7000 современных видов, от крошечных гекконов до комодосских варанов и анаконды. Squamata - это специализированные планеры, норники, бегуны, скалолазы и пловцы. Исторически (Romer, 1956), чешуйчатые были разделены на две группы, "Lacertilia" (ящерицы, амфисбены) и Ophidia (змеи), но филогенетический анализ показал, что «Lacertilia» в этом смысле не является монофилетической группой.
Основные апоморфии чешуйчатых, учитывая "мягкие" органы: редукция хрящевых компонентов передней мозговой коробки, связь слезного протока с якобсоновым органом, обособление последнего от главной части носовой капсулы, погружение септомаксиллы, прикрывающей у сквамат якобсонов орган, внутрь носовой капсулы, резкое укорочение носовых костей, перемещение надвисочной кости назад от соответствующего отростка теменной кости, процельность позвонков, наличие тироидной вырезки в тазовом поясе (имеется также у кюнеозаврид), слияние астрагала и пяточной кости с образованием мезотарзального сустава и крючковидность V метатарзальной кости. Кроме того, ящерицы современного типа обладают большим задневисочным окном, а большинство ящериц - также характерными вырезками по переднему краю коракоида и нижней части лопатки, отсутствие костных брюшных ребер (гастралий).
Ящерицы, или формы, близкие к ним, известны с самой верхней перми Южной Африки, но представители современных семейств появляются только в мелу, а более примитивные ящерицы современного облика, принадлежащие к ископаемым семействам современных подотрядов - в верхней юре. Ящерицы отличаются не только утратой нижней височной дуги и стрептостилией, но также и наличием в квадратной кости ушной вырезки, грацильной слуховой косточкой и связью парокципитального отростка с квадратной костью. Характерным у ящериц современного облика является то, что хвостовые позвонки зачастую поделены надвое автотомальной септой, облегчающей произвольную автотомию (отрыв хвоста).
Подтряд Eolacertilia. Доюрские «ящерицы» отличаются многими примитивными признаками, к числу которых относится парность всех костей крыши черепа, субтекодонтность зубов, амфицельность позвонков, пластинчатый тазовый пояс; метатарзальная кость пальца у них, хотя и сильно укорочена, но не крючковидная. К ним относится семейство Paliguanidae с родами Palaeagama, Paliguana и Saurosternon из самой верхней перми и нижнего триаса Южной Африки, а также Blomisaurus из нижнего триаса Кировской области. Более отличается от палигуанид Boreopricea из нижнего триаса о-ва Колгуев, имеющая сильно удлиненный череп, но сходная с Saurosternum парностью грудины [Татаринов, 1978]. Все это мелкие ящерицы с длиной тела без хвоста не более 15-25 см.
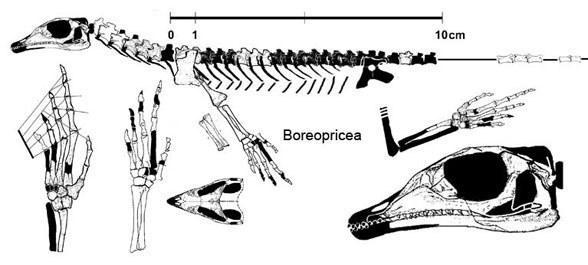
Boreopricea funerea (Tatarinov 1978, Benton and Allen 1997)
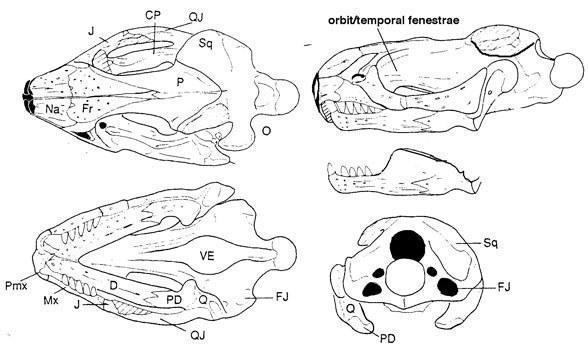
До недавнего времени палигуаниды считались древнейшими представителями ящеричного филогенетического ствола. Но недавно из Мезени, датируемого, возможно, верхами казанского или низами татарского яруса, был описан по черепу еще один возможный представитель мелких эолацертилий - Lanthanolania ivakhnenkoi (Modesto and Reisz 2003). У этого ящера имеется верхнее височное окно, окаймленное снизу треугольной заглазничной костью, а нижнее височное окно, по-видимому, широко открыто снизу, зубы мелкие, однородные.
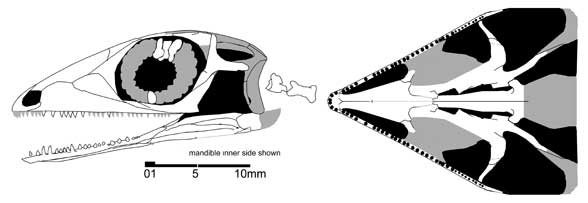
Lanthanolania ivakhnenkoi (Modesto and Reisz 2003)
Татаринов относит к эолацертилиям и кюнеозаврид, однако филогенетический анализ рассматривает этот таксон как более примитивный (Lepidosauria+Kuehneosauridae=Lepidosauriformes). Взгляды на филогению лепидозавров неоднозначны, и прямое родство эолацертилий и ящериц современного облика нуждается в дополнительном подтверждении.
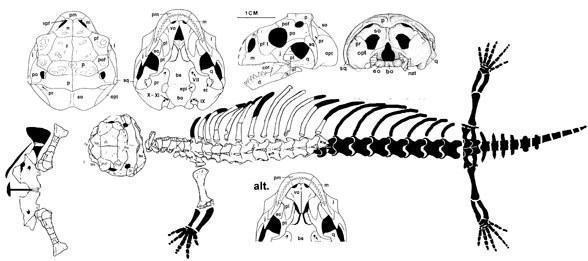
Ящерицы современного облика. Самые древние «настоящие» ящерицы обнаружены в верхней части средней юры Англии, где найдены остатки парамацеллодид, сцинкоморф (Saurillodon) и ангвиоморф (Parviraptor). К этой группе может относиться и род Marmoretta, также из средней юры Англии. Кроме того, из средней юры Мексики описан амфисбеноподобный диапсид Tamaulisaurus morenoi. Находки верхнеюрских ящериц уже более многочисленны, они происходят из США, Западной Европы, Средней (Центральной) Азии и Китая. Наиболее богатая верхнеюрская фауна ящериц происходит из Западной Европы, откуда известно семейство игуан Euposauridae, два семейства геккот (Ardeosauridae и Bavarisauridae), семейство кордилоидов Paramecellodidae. Все юрские ящерицы относятся к вымершим семействам, их связи с более древними лепидозавроморфами не улавливаются.
Marmoretta oxoniensis (Evans 1991)
Не менее разнообразная фауна верхнемеловых ящериц, насчитывая также более 40 видов, раскопана также в Монголии, в которой эта фауна отличается высоким эндемизмом и обилием вымерших семейств. В верхнем мелу Монголии зарегистрированы вымершие Arretosauridae, Hoplocercidae, возможно, Ardaeosauridae, Slavoiidae, Adamisauridae, Carusiidae, Paramacellodidae, Glyptosauridae, Necrosauridae, возможный дорсетозаврид, параваранид. Из современных семейств в верхнем мелу Монголии отмечены агамиды из группы круглоголовок, иногда выделямые в особое семейство, агамиды вымершего подсемейства Priscagaminae, изодонтозавровые агамиды, разнообразные тейиды, вероятный гимнодактилид, лацертид, гекконид, ксенозаврид и вараниды.
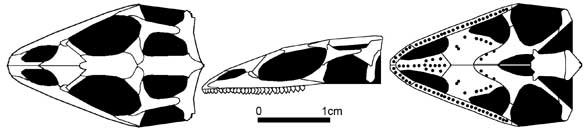
Заслуживает специального внимания одна из самых удивительных находок ископаемых меловых ящериц — это Sineoamphisbaena hexataburis из формации Джадохта Внутренней Монголии (Китай). Синеоамфисбена - некрупная (длина черепа около 4 см) ящерица с коротким и очень широким черепом, маленькой верхней височной ямой, обособленной снизу заглазничной костью, высокой мак- силлой, контактирующей с носовой костью, и плевродонтными зубами.
Sineoamphisbaena hextabularis (Wu et al. 1993)
В кайнозое быстро появляются современные роды ящериц, последние вымершие семейства - Arretosauridae и Necrosauridae - исчезают к концу эоцена-началу олигоцена.