Именно появление эукариотической клетки является одним из самых значимых событий биологической эволюции. Отличие эукариотических организмов от прокариотических состоит в более совершенной системе регуляции генома. А благодаря этому возросла приспособляемость одноклеточных организмов, их способность адаптироваться к меняющимся условиям среды без внесения наследственных изменений в геном. Благодаря возможности адаптироваться эукариоты смогли стать многоклеточными – в многоклеточном организме клетки с одним и тем же геномом, в зависимости от условий, образуют совершенно разные по морфологии и по функции ткани.
Этот ароморфоз произошел на рубеже Архея и Протерозоя (2,6 – 2,7 млрд. лет назад), что определили по биомаркерам – остаткам стероидных соединений, свойственных только эукариотическим клеткам. Появление эукариот совпадает по времени с кислородной революцией.
Считается общепризнанным, что эукариоты появились в результате симбиоза нескольких разновидностей прокариот. Видимо, митохондрии произошли от альфа-протеобактерий (аэробных эубактерий), пластиды – от цианобактерий, а цитоплазма – от неизвестной архебактерии. Пока еще нет общепринятой теории возникновения ядра, цитоскелета, а также жгутиков. Гипотезы возникновения жизни на Земле не внесли ясности в вопрос о возникновении клетки. Если о происхождении прокариотов практически нет никаких гипотез, правдоподобно описывающих их возникновение, то что касается происхождения эукариотических клеток, есть несколько точек зрения.
Основные гипотезы происхождения эукариот:
1. Симбиотическая гипотеза основана на двух концепциях. Согласно первой из этих концепций, самое фундаментальное разграничение в живой природе – это разграничение между бактериями и организмами, состоящими из клеток с истинными ядрами – протистами, животными, грибами и растениями. Вторая концепция состоит в том, что источником некоторых частей эукариотических клеток была эволюция симбиозов – формирование постоянных ассоциаций между организмами разных видов. Предполагается, что три класса органелл – митохондрии, реснички и фотосинтезирующие пластиды произошли от свободно живущих бактерий, которые в результате симбиоза были включены в состав клеток прокариот-хозяев. Эта теория в большой мере опирается на неодарвинистские представления, развитые генетиками, экологами, цитологами, которые связали Менделевскую генетику с Дарвиновской идеей естественного отбора. Она опирается также на молекулярную биологию, особенно на данные о структуре белков и последовательности аминокислот, на микропалеонтологию, изучающую наиболее ранние следы жизни на Земле, и на физику и химию атмосферы, поскольку эти науки имеют отношение к газам биологического происхождения.
2. Инвагинационная гипотеза говорит, что предковой формой эукариотической клетки был аэробный прокариот. Внутри находилось несколько геномов, прикреплявшихся к клеточной оболочке. Корпускулярные органеллы и ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. Потом в процессе эволюции произошло усложнение ядерного генома и появилась система цитоплазматических мембран. Эта гипотеза объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Но она встречается с трудностями в объяснении различий в деталях процесса биосинтеза белка в корпускулярных органеллах и цитоплазме эукариотической клетки. В митохондриях и хлоропластах этот процесс в точности соответствует таковому в современных прокариотических клетках.
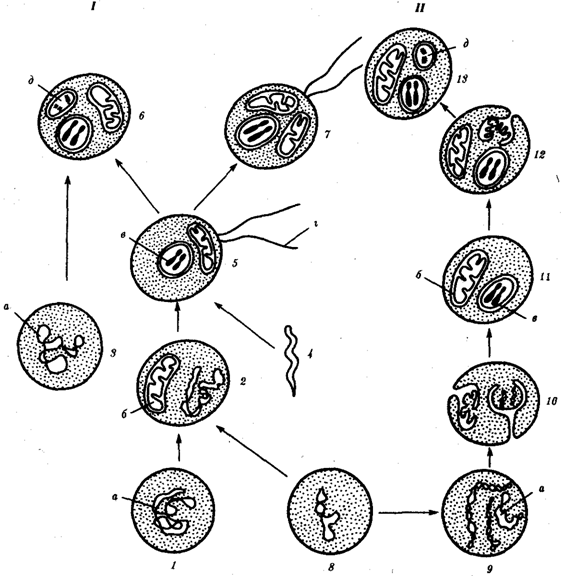
Происхождение эукариотической клетки согласно симбиотической (I) и инвагинационной (II) гипотезам:
1 — анаэробный прокариот (клетка-хозяин), 2 — прокариоты, имеющие митохондрии, 3 — сине-зеленая водоросль (презумптивный хлоропласт), 4 — сиирохетообразная бактерия (презумпгивный жгутик), 5 — примитивный эукариот со жгутиком, 6 — растительная клетка, 7 — животная клетка со жгутиком, 8 — аэробный прокариот (презумптивная митохондрия), 9 — аэробный прокариот (клетка-родоначальница согласно гипотезе II), 10 — инвагинации клеточной оболочки, давшие ядро и митохондрии, 11 — примитивный эукариот 12 — впячивание клеточной оболочки, давшее хлоропласт, 13 — растительная клетка; а —ДНК прокариотической клетки, б — митохондрия, в —ядро эукариотической клетки, г — жгутик, д — хлоропласт.
Имеющихся данных пока недостаточно для того, чтобы отдать предпочтение какой-то одной из гипотез или выработать новую, которая устроила бы большинство ученых, но в последние годы удалось убедительно доказать симбиогенетическую теорию происхождения эукариотической клетки.
Эволюционные возможности клеток эукариотического типа выше, чем прокариотического. Ведущая роль здесь принадлежит ядерному геному эукариот, который превосходит по размерам геном прокариот. Важные отличия заключаются в диплоидности эукариотических клеток благодаря наличию в ядрах двух комплектов генов, а также в многократном повторении некоторых генов.
Усложняется механизм регуляции жизнедеятельности клетки, что проявилось в увеличении относительного количества регуляторных генов, замене кольцевых «голых» молекул ДНК прокариот хромосомами, в которых ДНК соединена с белками.
Аэробное дыхание также послужило предпосылкой для развития многоклеточных форм. Сами эукариотические клетки появились на Земле после того, как концентрация O2 в атмосфере достигла 1% (точка Пастера). А эта концентрация –необходимое условием аэробного дыхания.
Известно, что каждая эукариотическая клетка содержит геномы различного происхождения: в клетках животных и грибов это геномы ядра и митохондрий, а в клетках растений – также и пластид. Небольшая кольцевая ДНК содержится и в базальном тельце жгутиков эукариотических клеток.
Согласно методу молекулярных часов эукариоты возникли тогда же, когда и прокариоты. Но очевидно же, что в течение значительной части истории Земли на ней доминировали прокариоты. Первые клетки, которые соответствуют эукариотическим размерам (акритархи), имеют возраст 3 млрд. лет, но их природа все еще остается неясной. Почти несомненные остатки эукариот имеют возраст около 2 млрд. лет. И лишь после кислородной революции на поверхности планеты сложились благоприятные условия для эукариот (около 1 млрд. лет назад).
Скорее всего, основным предком эукариотических клеток стали архебактерии, которые перешли к питанию путем заглатывания пищевых частиц. Изменение формы клетки, необходимое для такого заглатывания, обеспечивал состоящий из актина и миозина цитоскелет. Наследственный аппарат такой клетки переместился вглубь от ее изменчивой поверхности, сохранив при этом свою связь с мембраной. А уже это послужило причиной возникновения ядерной оболочки с ядерными порами.
Поглощенные клеткой-хозяином бактерии могли продолжить свое существование внутри нее. Так, предками митохондрий стала группа фотосинтезирующих бактерий – пурпурные альфапротеобактерии. Внутри клетки-хозяина они утратили способность к фотосинтезу и приняли на себя окисление органических веществ. Благодаря им эукариотические клетки стали аэробными. Симбиозы с другими фотосинтезирующими клетками стали причиной приобретения растительными клетками пластид. Возможно, жгутики эукариотических клеток произошли в результате симбиоза клеток-хозяев с бактериями, которые были способны к извивающимся движениям.
Наследственный аппарат эукариотических клеток был устроен примерно так же, как у прокариот. Но в связи с необходимостью управления более крупной и сложной клеткой, позже изменилась организация хромосом, а ДНК оказалась связана с белками-гистонами. Прокариотическая организация сохранилась у геномов внутриклеточных симбионтов.
В результате различных актов симбиогенеза возникли различные группы эукариотических организмов: эукариотической клетка + цианобактерия = красные водоросли; эукариотическая клетка + бактерия-прохлорофита = зеленые водоросли. Даже хлоропласты золотистых, диатомовых, бурых и криптомонадовых водорослей возникли в результате двух последовательных симбиозов, о чем говорит наличие у них 4 мембран.
Появление эукариот было приурочено к такому периоду истории биосферы, когда условия были особенно нестабильны и непредсказуемы, когда приспособительная стратегия прокариот (быстрое мутирование, горизонтальный обмен генами и отбор устойчивых клонов) оказалась слишком расточительной и недостаточно эффективной. В такой ситуации большое преимущество могла получить принципиально более универсальная и экономичная адаптивная стратегия, основанная на развитии целесообразной модификационной изменчивости.
Пожалуй, становление эукариот и развитие у них полового процесса сделали структуру изменчивости и биоразнообразия более дискретной и «управляемой» – это должно было привести к ускоренному росту биоразнообразия и к повышению эволюционной пластичности и экологической толерантности видов, сообществ и биоты в целом.
Появление эукариот можно по праву назвать «эталонным» ароморфозом. В этом событии предельно ярко проявилась общая прогрессивная направленность биологической эволюции. Прогресс выразился не только в усложнении организации, расширении суммарной адаптивной зоны жизни, росте биомассы и численности, повышении автономности организмов но и в повышении устойчивости живых систем.
На примере эукариот отчетливо показано, что появление новых форм жизни следует рассматривать не как результат эволюции каких-то отдельных филетических линий или клад, а как закономерный и неизбежный эффект развития систем высшего порядка – сообществ, биосферы и, возможно, всей планеты как единого целого.
Используемые источники:
А. В. Марков, А. М. Куликов. Происхождение эукариот как результат интеграционных процессов в микробном сообществе
А. В. Марков. Проблема происхождения эукариот
М. В. Ларина. Гипотезы происхождения эукариотических клеток. Возникновения многоклеточности